Способ увеличения содержания сложного эфира сахарозы в растениях табака - RU2758117C2
Код документа: RU2758117C2
Чертежи















Описание
Область техники, к которой относится изобретение
Настоящее изобретение относится к улучшению вкуса табака и, в частности, касается содержания сложного эфира сахарозы в растениях табака. В частности, изобретение относится к увеличению содержания сложного эфира сахарозы в растениях табака. Более конкретно, изобретение относится к способам и применениям, ингибирующим активность или экспрессию гена синтеза дитерпена, а также к растениям табака и продуктам их переработки (например, материалам для размножения, собранным листьям, обработанному табаку, табачным изделиям).
Предшествующий уровень техники
Трихомы растений представляют собой специализированные структуры, включающие эпидермальные выросты, которые можно обнаружить на надземных поверхностях многих растений. Их морфология варьирует между видами и даже внутри вида, включая их расположение на органах растений, их размер и плотность. Считается, что они обеспечивают первую линию защиты от насекомых, патогенных микробов и травоядных животных и обеспечивают защиту от окружающей среды, например, от морозных и ветреных условий. Трихомы могут быть железистыми и секретировать продукты, основными функциями которых может быть производство химических веществ, взаимодействующих с вредителями или опылителями, которые хранятся или улетучиваются на поверхности растения. Трихомы таких растений могут служить фабриками по производству натуральных продуктов, имеющих экономическое и коммерческое значение. Преимущественно выделяемые соединения вырабатываются только в трихомных железах и накапливаются вне клеток желез под окружающей кутикулой. Следовательно, экссудат может быть просто и аккуратно извлечен путем погружения растения в неинвазивный растворитель, поскольку материал экссудата по существу накапливается вне растения. Манипуляции с композицией вторичного экссудата растительных трихом были использованы для повышения устойчивости к насекомым (Wang et al. Nature Biotechnology 19, 371–374 (2001), которая включена в настоящее описание ссылкой), а метаболическая инженерия железистых трихом использовалась для улучшения качества эфирных масел (Mahmoud and Croteau Proc Natl Acad Sci USA. 2001 Jul 17; 98 (15): 8915-20. Epub 2001 Jun 26, который включен в настоящий документ ссылкой).
Ряд сортов табака обладают как высокой биомассой, так и высоким потенциалом накопления экссудата. Например, экспериментальная табачная линия TI 1068 может производить до 17% от сухой массы листьев в виде экссудата железистых трихом (Wagner, GJ 1991 Plant Physiology 96, 675–679, который включен в данное описание ссылкой). Примерно 72% экссудата железы трихомы составляют дитерпеноиды и около 24% - сложные эфиры сахарозы.
По-видимому, самым богатым природным источником сложных эфиров сахарозы являются растения табака. Сложные эфиры сахара представляют собой сложные смеси соединений из-за множества возможных ацильных групп и комбинаций ацильных групп, которые могут быть этерифицированы до сахара (сахароза или глюкоза). Эти соединения большей частью ответственны за липкую текстуру листьев табака, которые их производят (Kandra L., GJ Wagner, Archiv es Biochem. Biophys. 265 (1988) 425–432, которая включена в настоящее описание ссылкой). Сложные эфиры сахарозы являются предшественниками 3-метилвалериановой и 3-метилмасляной кислот, которые представляют собой важные вкусовые компоненты дыма турецкого табака (Kandra, L., et al. Eur. J. Biochem. 188 (1990) 385–391, которая включена в данное описание ссылкой), кроме того, считается, что 3-метилвалериановые кислоты обеспечивают «турецкую ноту» для табака.
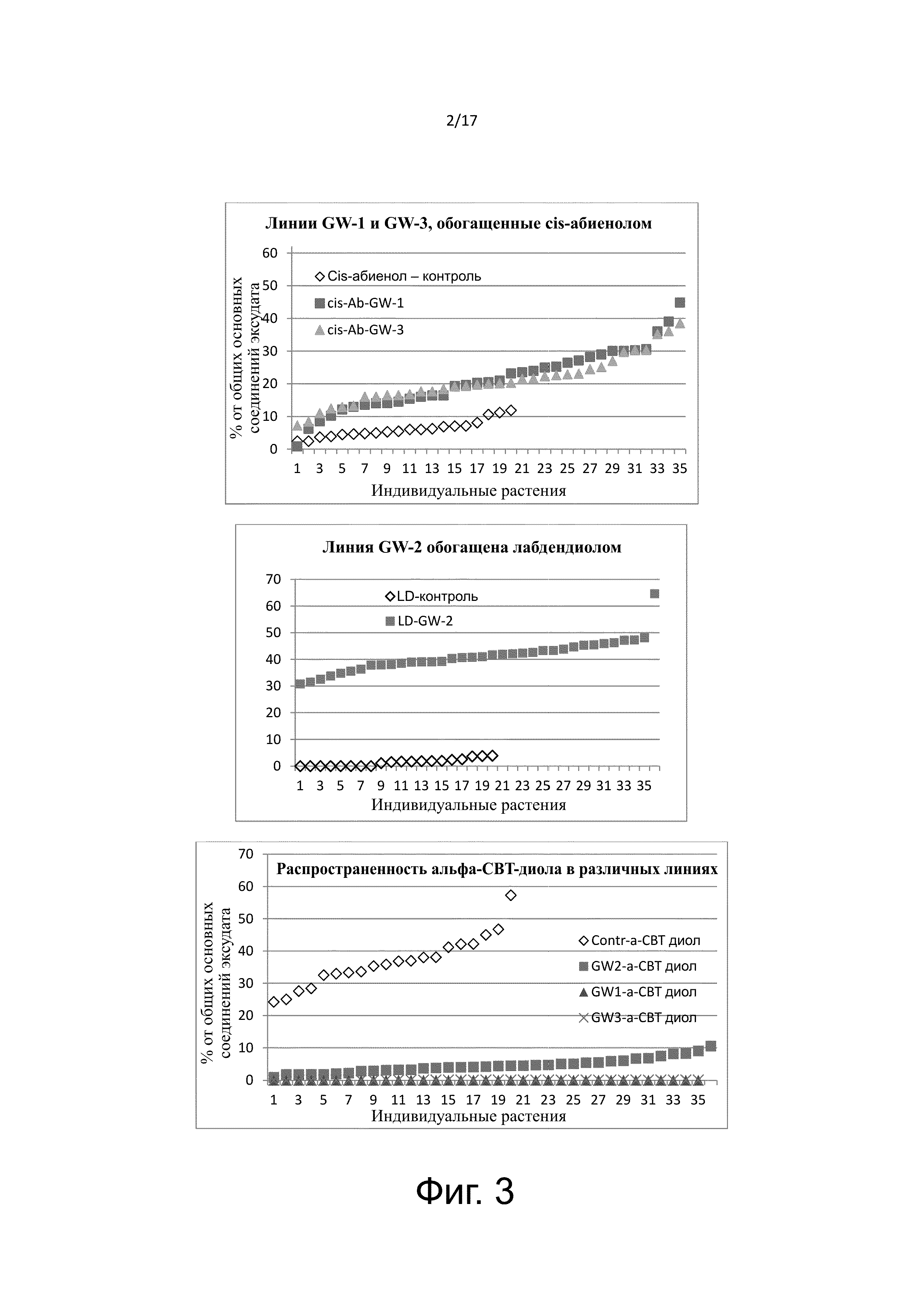
Экссудат трихом TI 1068 содержит в основном цембреновые и лабдановые дитерпеноиды. CBT-диолы (α- и β-позиционные изомеры), цис-абиенол и лабдендиол составляют около 60, 10 и 0,6% от массы экссудата, соответственно. Общий путь синтеза основных дитерпеновых компонентов экссудата трихом TI 1068 и CBT-диолов показаны на Фигуре 2. Метилэритритолфосфатный (MEP) путь хлоропласта поставляет геранилгеранилпирофосфат (GGPP) для синтеза дитерпена.
Органолептические или сенсорные свойства разных типов табака значительно различаются и зависят от сложности факторов, включая генетические различия. Считается, что противогрибковые и органолептические свойства табака связаны с содержанием сложного эфира сахарозы. Уникальный аромат и вкусовой профиль табака являются результатом уникальных вкусовых и ароматических соединений или предшественников этих соединений, которые присутствуют на определенных уровнях в высушенном листе и могут включать содержание сложного эфира сахарозы.
Турецкий табак (иногда называемый восточным табаком) обладает желательными органолептическими свойствами; это очень ароматный, малолистный табак с мягким вкусом и содержит меньше никотина, чем другие сорта табака. Было установлено, что измеримое количество сложного эфира сахарозы переживает процессы сушки/выдержки табака и, таким образом, является неотъемлемым компонентом коммерческих турецких (или восточных) табаков (M. Ashraf-Khorassani,et al. Contributions to Tobacco Research Vol 21: 7;October 2005, который включен в данное описание ссылкой). Кроме того, сложные эфиры сахарозы являются предшественниками ароматических карбоновых кислот в дыме турецкого табака и, таким образом, являются важными вкусовыми компонентами турецкого табака (Arrendale, RF, et al., J. Agric. Food Chem. 38 (1990) 75–85, который включен в данное описание ссылкой).
Природные эфиры сахарозы представляют сложные аналитические проблемы из-за степени этерификации и распределения функциональных эфиров в молекуле. В молекуле сахарозы имеется четыре сайта для потенциальных ацильных групп (например, одна первичная, три вторичных) на глюкозном (GLU) кольце; и еще четыре сайта (например, два первичных, два вторичных) можно найти на кольце фруктозы (FRU). До настоящего времени идентификация сложноэфирных кислотных групп проводилась с помощью GC-MS (газовая хроматография - масс-спектрометрия) и LC-MS (жидкостная хроматография-масс-спектрометрия) (см. Slocombe et al. Plant Physiol. 148, 1830-1846, который включен в данное описание ссылкой) либо метил/бутиловых эфиров сахарозы, либо триметилсилил (TMS) производных сахарозы. Высокотемпературная GC (газовая хроматография) была использована в исследовании с минимальным успехом в попытке охватить производные углеводов с более высокой молекулярной массой (Karrer R. et al., J. High Resolu. Chromatogr. 15 (1992) 585–589, которая включена в настоящее описание ссылкой). Были получены предположения относительно характера ацильного замещения на сложном эфире сахарозы из данных, полученных кислотным гидролизом сложного эфира и повторной дериватизации высвобожденной карбоновой кислоты, но этот подход может быть подвергнут различным интерпретациям.
Сущность изобретения
Неожиданно было обнаружено, что путем ингибирования активности или экспрессии гена синтеза дитерпена, как указано в данном документе, отличительные вкусовые (и/или ароматические и/или вкусовые) характеристики, такие как те, которые обнаружены в турецком табаке, могут быть получены растениями табака с коммерчески желательными признаками путем модификации табака для производства табака с повышенным содержанием сложного эфира сахарозы. Таким образом, могут быть получены табачные изделия с превосходными вкусовыми (и/или ароматическими и/или вкусовыми) характеристиками, востребованные потребителями табачных изделий.
Авторы настоящего изобретения исследовали регуляцию и пути потока углерода в специализированных железистых клетках трихом, которые продуцируют специфические вторичные соединения. Одной из целей изобретателей была оптимизация метаболизма дитерпена трихомовой железы для изменения химического состава экссудата трихом для обеспечения извещенного содержания дитерпена. Были созданы растительные клеточные линии, которые неожиданно продемонстрировали повышенное содержание сложного эфира сахарозы в экссудате по сравнению с их аналогами растений дикого типа, выращенными в тех же условиях. Линии растений предназначались для ферментов, катализирующих образование CBTol (синтазы) и терпен-синтазы 3-8. Терпен-синтаза 3-8 в некоторых воплощениях может упоминаться как цис-абиенолсинтаза.
Авторы настоящего изобретения неожиданно определили способ увеличения содержания сложного эфира сахарозы в растении табака путем ингибирования активности или экспрессии гена синтеза дитерпена. До настоящего изобретения не было известно, что ингибирование активности или экспрессии гена синтеза дитерпена можно использовать для увеличения содержания сложного эфира сахарозы, особенно при улучшении выхода и других коммерчески желательных признаков.
Согласно одному аспекту настоящее изобретение относится к способу увеличения содержания сложного эфира сахарозы в растении табака или его части или в культуре клеток табака, причем способ включает модификацию указанной культуры растения табака или клеток табака путем ингибирования активности или экспрессии гена синтеза дитерпена.
В другом аспекте предложено применение гена синтеза дитерпена для увеличения содержания сложного эфира сахарозы в растении табака или его части или в культуре клеток табака.
В другом аспекте настоящее изобретение относится к способу получения растения табака или его части, материала для размножения растения табака, листа табака, резанного собранного листа табака, обработанного листа табака или резаного и обработанного листа табака или культуры клеток табака, который имеет повышенное содержание сложного эфира сахарозы, причем способ включает модификацию указанного табака для ингибирования активности или экспрессии гена синтеза дитерпена.
В другом аспекте предлагается способ или применение согласно изобретению, в котором содержание сложного эфира сахарозы увеличивается по сравнению с культурой растения табака или клеток табака, которая не была модифицирована для ингибирования активности или экспрессии гена синтеза дитерпена.
В другом аспекте предложено растение табака или его часть или культура клеток табака, которые были модифицированы для достижения увеличения содержания сложного эфира сахарозы по сравнению с немодифицированным растением или культурой немодифицированных клеток табака, где модификация представляет собой ингибирование активности или экспрессии гена синтеза дитерпена.
В другом аспекте предлагается материал для размножения табачных растений (например, семена растений), который можно получить из растения табака или культуры клеток табака в соответствии с изобретением.
В другом аспекте изобретение относится к способу или применению, растению табака или материалу для размножения растения табака, или культуре клеток табака, где ген синтеза дитерпена представляет собой ген циклазы 2 (CYC2), CBTol-циклазы или терпен-синтазы 3-8.
Изобретение относится к одному аспекту способа или применения, растения табака или материала для размножения растения табака, где ген синтеза дитерпена представляет собой ген циклазы 2 (CYC2).
Изобретение относится к одному аспекту способа или применения, растения табака или материала для размножения растения табака, где ген синтеза дитерпена представляет собой CBTol-циклазу.
Изобретение относится к одному аспекту способа или применения, растения табака или материала для размножения растения табака, где ген синтеза дитерпена представляет собой терпен-синтазу 3-8.
Изобретение относится к другому аспекту способа или применения, растения табака или его части или материала для размножения растения табака или культуры клеток табака в соответствии с изобретением, где экспрессия гена синтеза дитерпена ингибируется с применением РНК-интерференции (RNAi).
В другом аспекте предлагается способ, применение, растение табака или материал для размножения растения табака, или культура клеток табака, где экспрессия гена циклазы 2 (CYC2) ингибируется с применением ДНК-конструкции ddRNAi, содержащей, по меньшей мере, часть экзона 1, по меньшей мере, часть экзон 2 и, по меньшей мере, часть экзона 3 гена циклазы 2 (CYC2).
В другом аспекте предлагается способ, применение, растение табака или материал для размножения растения табака, или культура клеток табака, в которых экспрессия гена циклазы 2 (CYC2) ингибируется с помощью dsRNA, которая обрабатывается эндогенными путями в клетке в одноцепочечную короткую интерферирующую РНК (siRNA), которая направляет индуцированный РНК комплекс сайленсинга (RISC) для связывания с мРНК, соответствующей, по меньшей мере, части экзона 1, по меньшей мере, части экзона 2 и, по меньшей мере, части экзона 3 гена циклазы 2 (CYC2). В одном аспекте одноцепочечная siRNA продуцирует антисмысловую (или направляющую) цепь, которая имеет комплементарную последовательность к мРНК, соответствующей, по меньшей мере, части экзона 1, по меньшей мере, части экзона 2 и, по меньшей мере, части экзона 3 гена циклазы 2 (CYC2).
В другом аспекте предлагается способ, применение, растение табака или материал для размножения растения табака, или культура клеток табака в соответствии с изобретением, где экспрессия гена CBTol-циклазы ингибируется с применением ДНК-конструкции ddRNAi, содержащей, по меньшей мере, часть экзона 4, интрон 4. экзон 5, интрон 5, экзон 6, интрон 6 и, по меньшей мере, часть экзона 7 гена CBTol-циклазы.
В другом аспекте предложен способ, применение, растение табака или материал для размножения растения табака, или культура клеток табака в соответствии с изобретением, где экспрессия гена CBTol-циклазы ингибируется при применении dsRNA, которая обрабатывается эндогенными путями в клетке в одноцепочечные siRNA, которая направляет индуцированный РНК комплекс сайленсинга (RISC) связываться с мРНК, соответствующей, по меньшей мере, части экзона 4, интрона 4, экзона 5, интрона 5, экзона 6, интрона 6 и, по меньшей мере, части экзона 7 CBTol-циклазы. В одном аспекте одноцепочечная siRNA продуцирует антисмысловую (или направляющую) цепь, которая имеет комплементарную последовательность к мРНК, соответствующую, по меньшей мере, части экзона 4, экзона 5, экзона 6 и, по меньшей мере, части экзона 7 гена CBTol-циклазы.
В другом аспекте предлагается способ, применение, растение табака или материал для размножения растения табака, или культура клеток табака в соответствии с изобретением, где экспрессия гена терпен-синтазы 3-8 ингибируется с применением ДНК-конструкции ddRNAi, содержащей последовательность, которая соответствует наименьшее количество нуклеотидов с 1497 по 1517 из SEQ ID No. 3.
В другом аспекте предлагается способ, применение, растение табака или материал для размножения растения табака, или культура клеток табака в соответствии с изобретением, где экспрессия гена терпен-синтазы 3-8 ингибируется с применением искусственной microRNAi, (amiRNAi), содержащей последовательность, которая соответствует, по меньшей мере, нуклеотидам с 1497 по 1517 из SEQ ID No. 3.
В другом аспекте предлагается способ, применение, растение табака или материал для размножения растения табака, или культура клеток табака в соответствии с изобретением, где экспрессия гена терпен-синтазы 3-8 ингибируется с применением ДНК-конструкции ddRNAi, содержащей последовательность, которая соответствует наименьшее количество нуклеотидов с 884 по 904 из SEQ ID No. 3.
В другом аспекте предлагается способ, применение, растение табака или материал для размножения растения табака, или культура клеток табака в соответствии с изобретением, где экспрессия гена терпен-синтазы 3-8 ингибируется с применением искусственной microRNAi (amiRNAi), содержащей последовательность, которая соответствует, по меньшей мере, нуклеотидам 884-904 SEQ ID No. 3.
В одном аспекте предлагается способ или применение изобретения, растение табака или его часть по изобретению, или материал для размножения растения табака, или культура клеток табака по изобретению, где содержание сложного эфира сахарозы в растении табака, по меньшей мере, в 2 раза (соответственно, по меньшей мере, в 3 раза) выше в модифицированной культуре растения табака или клеток табака по сравнению с культурой растения табака или клеток табака, которая не была модифицирована для ингибирования активности или экспрессии гена синтеза дитерпена.
В одном аспекте предлагается применение растения табака или его части по изобретению для селекции растения табака.
В другом аспекте изобретение относится к применению растения табака или его части или культуры клеток табака по изобретению для производства продукта табачной промышленности.
В другом аспекте предложено применение растения табака или его части по изобретению для выращивания культуры.
В одном аспекте изобретение относится к высушенному табачному материалу, изготовленному из растения или его части в соответствии с изобретением или его экстракта или культуры табачных клеток в соответствии с изобретением.
В другом аспекте предлагается табачная смесь, содержащая указанный сушенный табачный материал согласно изобретению.
В одном аспекте предлагается применение растения табака или его части по изобретению для получения листа табака (например, обработанного (предпочтительно сушенного) листа табака).
В другом аспекте предлагается собранный лист растения табака по изобретению или получаемый из растения табака, размноженного из материала для размножения по изобретению или получаемого из растения табака, полученного с применением изобретения.
В другом аспекте предлагается собранный лист растения табака по изобретению, где собранный лист растения табака представляет собой резанный собранный лист.
Изобретение обеспечивает в другом аспекте обработанный табачный лист (предпочтительно нежизнеспособный обработанный табачный лист):
i) получаемый из растения табака, получаемого при применении изобретения;
ii) получаемый обработкой растения табака по изобретению;
iii) получаемый из растения табака, размноженного из материала для размножения растения табака по изобретению; или
iv) получаемый обработкой собранного листа растения табака по изобретению.
В другом аспекте предлагается обработанный табачный лист по изобретению, в котором табак обрабатывают путем отверждения, ферментации, пастеризации или их комбинации.
В другом аспекте предлагается обработанный табачный лист по изобретению, в котором обработанный табачный лист представляет собой нарезанный обработанный табачный лист.
Изобретение обеспечивает в другом аспекте продукт табачной промышленности, приготовленный из:
i) растения табака по изобретению или его части или культуры клеток табака по изобретению;
ii) растения табака или его части, размножаемой из материала для размножения растения табака по изобретению;
iii) собранного листа растения табака по изобретению;
iv) обработанного табачного листа по изобретению.
В другом аспекте предлагается продукт табачной промышленности по изобретению, в котором табачный продукт представляет собой курительное изделие.
В другом аспекте предложен продукт табачной промышленности по изобретению, в котором табачный продукт представляет собой бездымный табачный продукт.
В другом аспекте предлагается продукт табачной промышленности по изобретению, в котором табачный продукт представляет собой устройство для нагревания табака, такое как устройство, генерирующее аэрозоль.
В одном аспекте предлагается курительное изделие, бездымный табачный продукт или устройство для нагревания табака, содержащее растение или его часть в соответствии с изобретением или его экстракт (например, экстракт табака) или культуру клеток табака в соответствии с изобретением; или сушенный табачный материал согласно изобретению; или табачная смесь по изобретению.
В другом аспекте предлагается способ, лист табака, растение табака, материал для размножения растения табака, собранный лист, обработанный табак, табачный продукт, применение или их комбинация, по существу, как описано в данном документе со ссылкой на описание и чертежи.
Краткое описание чертежей
Воплощения изобретения теперь будут описаны только в качестве примера со ссылкой на прилагаемые чертежи, на которых:
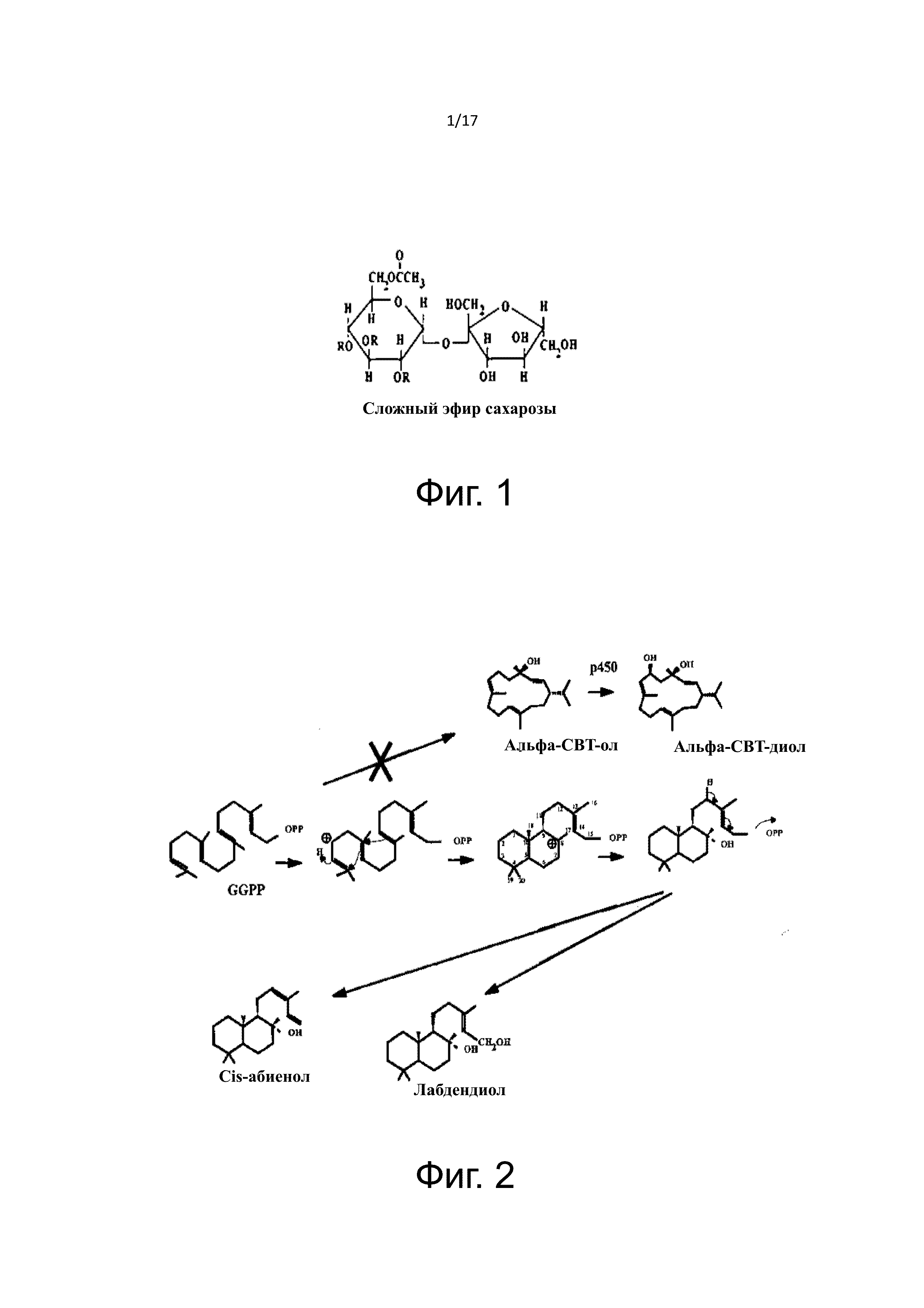
На Фигуре 1 показана общая структура сложного эфира сахарозы.
На Фигуре 2 показана схема пути синтеза дитерпена в пластидах.
На Фигуре 3 показаны графики, демонстрирующие анализ зеленых листьев контрольных линий, выращенных в полевых условиях, в сравнении с трансгенными линиями. Индивидуальные химические составы растений расположены в порядке возрастания количества.
На Фигуре 4 показан график, демонстрирующий общее содержание сложного эфира сахарозы в контрольных и трансгенных линиях.
На Фигуре 5 показан график, демонстрирующий обогащение эфиром сахарозы в линии GW-3 в сравнении с контролем. Содержание сложного эфира сахарозы представлено как процент от общего количества основных соединений экссудата.
На Фигуре 6 показан график, демонстрирующий обогащение цис-абиенолом в линии GW-3 в сравнении с контролем. Содержание цис-абиенола представлено как процент от общего количества основных соединений экссудата.
На Фигуре 7 показана SEQ ID No. 1, описанная ниже.
На Фигуре 8 показана SEQ ID No. 2, описанная ниже.
На Фигуре 9 показана SEQ ID No. 3, описанная ниже.
На Фигуре 10 показана SEQ ID No. 4, описанная ниже.
На Фигуре 11 показана SEQ ID No. 5, описанная ниже.
На Фигуре 12 показана SEQ ID No. 6, описанная ниже.
На Фигуре 13 показана SEQ ID No. 7, описанная ниже.
На Фигуре 14 показана SEQ ID No. 8, описанная ниже.
На Фигуре 15 показана SEQ ID No. 9, описанная ниже.
На Фигуре 16 показана SEQ ID No. 10, описанная ниже.
На Фигуре 17 показана SEQ ID No. 11, описанная ниже.
Перечень последовательностей
Предлагается сводка идентификаторов последовательностей, используемых в предметном описании, и соответствующий перечень последовательностей, где:
SEQ ID No. 1 соответствует нуклеотидной последовательности, кодирующей ген CBTol-циклазы. Эта нуклеотидная последовательность аннотирована в Genbank: AY049090.
SEQ ID No. 2 соответствует аминокислотной последовательности полипептида CBTol-циклазы.
SEQ ID No. 3 соответствует кДНК-последовательности гена, то есть кодирующей последовательности гена терпен-синтазы 3-8 из Nicotiana tabacum. Последовательность гена кДНК полной длины аннотирована в Genbank: AY528645.
SEQ ID No. 4 соответствует аминокислотной последовательности полипептид-терпен-синтазы 3-8 из Nicotiana tabacum.
SEQ ID No. 5 содержит по порядку 5'-смысловой фрагмент от 54 до 716 нуклеотида из последовательности AF401234 (который является полной кодирующей последовательностью мРНК CYC2 и соответствует нумерации нуклеотидов в SEQ ID No. 9); частичный фрагмент GUS A в виде петли-шпильки (от 787 до 1812 нуклеотида); и обратный комплемент смыслового 3'-фрагмента. На фигуре подчеркнутая последовательность соответствует фрагменту GUS A (AF502128) от 787 до 1812 нуклеотида. Последовательность от 54 до 716 нуклеотида из мРНК CYC2 (AF401234, соответствующая SEQ ID No. 9) в смысловой ориентации соответствует следующим геномным последовательностям из AY495694 (AY495694 является геномной последовательностью гена CYC2 и соответствует SEQ ID No. 8): нуклеотиды с 1 по 25, 1-й экзон (нуклеотиды с 26 по 271), 2-й экзон (нуклеотиды с 1253 до 1529) и первые 115 нуклеотидов 3 экзона (нуклеотиды с 2366 по 2480), где нумерация нуклеотидов соответствует нумерации в SEQ ID No. 8. SEQ ID No. 5 кодирует dsRNA и используется в ДНК-конструкции ddRNAi (иногда называемой в данном документе GW1) для получения dsRNA.
SEQ ID No. 6 содержит последовательность из гена CBTol-циклазы Nicotiana tabacum, содержащую частичную последовательность экзона 4, интрона 4, экзона 5, интрона 5, экзона 6, интрона 6 и частичного экзона 7 смысловой и антисмысловой ориентации, разделенных спейсером GUS. Элементы частичного гена показаны на фигуре в следующем порядке: ген N. tabacum cyclase (AY049090), частичный экзон 4 - от нуклеотида 2854 до нуклеотида 4175, прямая ориентация, частичный экзон 4, интрон 4 (жирным шрифтом), экзон 5, интрон 5 (жирным шрифтом), экзон 6, интрон 6 (жирным шрифтом), частичный экзон 7 (соответствует нуклеотидам с 2854 по 4175 из SEQ ID No. 1); затем следует петля GUS - Частичный ген GUS A (учетный номер Genbank AF502128) - с 786 по 1816 (подчеркнуто); за ним следует та же последовательность гена циклазы N. tabacum (AY049090), но в обратном комплементе (обратный комплемент показан штриховкой). Эта последовательность кодирует dsRNA и используется в ДНК-конструкции ddRNAi (иногда называемой в данном документе GW3) для получения dsRNA.
SEQ ID No. 7 содержит 21 нуклеотидную последовательность из гена терпен-синтазы 3-8 из Nicotiana tabacum и последовательности, которые соответствуют последовательности miRNA168 Arabidopsis. Последовательность соответствует Arabidopsis miRNA168a, первая и вторая последовательности, выделенные курсивом, соответствуют 21 нуклеотидной последовательности из гена терпен-синтазы 3-8 AY528645 (с 1497 по 1517 нт). Первая последовательность, выделенная курсивом (отмечена) находится в ориентации обратного комплемента. Вторая последовательность, выделенная курсивом (отмечена) находится в прямой ориентации и имеет три модификации (выделено жирным шрифтом). Последовательность кодирует amiRNAi и используется в ДНК-конструкции ddRNAi (иногда называемой в данном документе GW2) для получения amiRNAi.
SEQ ID No. 8 соответствует геномной последовательности гена циклазы 2 (CYC2). Эта нуклеотидная последовательность аннотирована в Genbank: AY495694.
SEQ ID No. 9 соответствует полной кодирующей последовательности мРНК гена циклазы 2 (CYC2), аннотированной в Genbank: (AF401234).
SEQ ID No. 10 соответствует аминокислотной последовательности полипептида, кодируемого геном циклазы 2 (CYC2) SEQ ID No. 9.
SEQ ID No. 11 содержит 21 нуклеотидную последовательность из гена терпен-синтазы 3-8 из Nicotiana tabacum и последовательности, которые соответствуют последовательности miRNA168a Arabidopsis. Последовательность соответствует последовательности miRNA168a Arabidopsis, первая и вторая последовательности, выделенные курсивом, соответствуют 21 нуклеотидной последовательности гена терпен-синтазы 3-8 AY528645 (от 884 до 904 нт) в обратном комплементе. Первая последовательность, выделенная курсивом (отмечена) находится в ориентации обратного комплемента. Вторая последовательность, выделенная курсивом (отмечена) находится в прямой ориентации и имеет модификации (выделены жирным шрифтом). Последовательность кодирует amiRNAi и используется в ДНК-конструкции ddRNAi (иногда называемой в данном документе GW5) для получения amiRNAi.
Подробное описание
Авторы настоящего изобретения впервые показали, что путем ингибирования активности или экспрессии гена синтеза дитерпена в табаке содержание сложного эфира сахарозы в растении табака может быть увеличено. Не желая быть связанными какой-либо теорией, полагаем, что ингибирование гена синтеза дитерпена приводит к уменьшенному использованию углерода для получения дитерпенов и, таким образом, к увеличению выработки эфира сахарозы.
В одном воплощении настоящее изобретение относится к способу увеличения содержания сложного эфира сахарозы в растении табака, включающему модификацию указанного растения табака путем ингибирования активности или экспрессии гена синтеза дитерпена.
Подходящим геном синтеза дитерпена для применения в изобретении может быть ген циклазы 2 (CYC2). Подходящим геном синтеза дитерпена для применения в изобретении может быть ген CBTol-циклазы. Соответственно, ген синтеза дитерпена для применения в изобретении может представлять собой ген терпен-синтазы 3-8.
Термин «увеличение» используется в данном документе для обозначения того, что концентрация и/или общее содержание сложного эфира сахарозы в продукте согласно настоящему изобретению (например, растении, его части (например, листе), обработанном листе или табачном продукте) выше по сравнению с сопоставимым продуктом, который не был модифицирован в соответствии с настоящим изобретением.
Термин «сравнимый продукт», как он определен в данном описании, будет происходить от растения табака, которое не было модифицировано в соответствии с настоящим изобретением, но в котором все другие соответствующие признаки были одинаковыми (например, вид растения, условия выращивания, способ обработки табака и т.д.). Сравнимый продукт в соответствии с настоящим изобретением может означать растение табака или его часть, такое как лист табака, собранный лист, собранный срезанный лист, обработанный табачный лист или материал для размножения растения табака или табачный продукт или их комбинации, получаемые или полученные из растения табака, которое не было модифицировано для ингибирования активности или экспрессии гена синтеза дитерпена. Сравнимые продукты также могут быть известны как контроли. В одном воплощении сравнимый продукт представляет собой продукт, который не содержит гена синтеза дитерпена, активность или экспрессия которого была ингибирована.
Термин «немодифицированное растение», как он определен в данном описании, будет представлять собой растение табака, которое не было модифицировано в соответствии с настоящим изобретением для ингибирования активности или экспрессии гена синтеза дитерпена и в котором все другие соответствующие признаки были одинаковыми (например, виды растений, условия выращивания, способ переработки табака и т.п.). В одном воплощении немодифицированное растение представляет собой растение, которое не содержит гена синтеза дитерпена, активность или экспрессия которого ингибирована.
В дополнительном аспекте содержание сложного эфира сахарозы измеряют по зеленым листьям. В дополнительном аспекте содержание сложного эфира сахарозы измеряют по отвержденным листьям, например, листьям воздушной сушки, дымовой сушки, огневой сушки или сушки на солнце. В дополнительном аспекте содержание сложного эфира сахарозы измеряют по листьям дымовой сушки. В дополнительном аспекте содержание сложного эфира сахарозы измеряют по листьям воздушной сушки.
Термин «содержание сложного эфира сахарозы» используется в данном документе для обозначения концентрации и/или общего содержания всей группы соединений, классифицируемых как сложные эфиры сахарозы. Сложные эфиры сахарозы, обычно присутствующие в табаке, могут быть представлены формулой, показанной на Фигуре 1.
Для определения концентрации и/или общего содержания сложных эфиров сахарозы может быть использован любой способ, известный в данной области. Один предпочтительный способ анализа сложных эфиров сахарозы (SE) включает анализ ацильных групп, высвобождаемых из сахарозы путем омыления, с последующим анализом их сложных бутиловых эфиров с помощью GC-MS. Анализ с помощью этого способа обеспечивает точное количественное определение эфира сахарозы. Альтернативный способ включает анализ сложных эфиров сахарозы путем измерения сахарозы на мкг/см2 поверхности листа. Определение содержания сложного эфира сахарозы может быть оценено в листе табака. Подходящие способы анализа содержания сложного эфира сахарозы могут включать стадии омыления, например, помещение образцов камеди в 80% метанол на ночь при 22°С для омыления. Затем образцы могут быть распределены между гексаном и водой. Для разделения образцы могут быть сначала высушены, например, в атмосфере N2, затем могут быть добавлены н-BuOH и H2SO4. Образцы могут быть нагреты до 110° в течение 1 часа до импульсного встряхивания. Могут быть сделаны экстракции и высушенный образец может быть растворен в CHCl3 для переноса в другую пробирку и высушен под N2. Образцы могут быть растворены в диметилформамиде (DMF) и бис(триметилсилил)трифторацетамиде. Образцы могут быть дериватизированы, например, при 70°С, прежде чем они будут охлаждены до комнатной температуры. Дериватизированные образцы могут быть проанализированы с помощью GC-MS. Элюированные соединения можно идентифицировать по времени их удерживания и сопоставлению профилей МС со стандартами.
В одном воплощении предложен способ получения растения табака, материала для размножения растения табака, табачного листа, собранного листа, резаного собранного листа, обработанного табачного листа, резаного и обработанного табачного листа, табачного продукта или их комбинаций, которые можно получить или получить с помощью растение табака по изобретению, которое имеет повышенное содержание сложного эфира сахарозы, причем способ включает модификацию указанного табака для ингибирования активности или экспрессии гена синтеза дитерпена. Повышенное содержание сложного эфира сахарозы может быть определено путем сравнения содержания сложного эфира сахарозы в растении табака, материале для размножения табака, листе табака, собранном листе, листе срезанного урожая, листе обработанного табака, листе разрезанного и обработанного табака, табачном продукте или их комбинациях с сопоставимым продуктом.
Соответственно, содержание эфира сахарозы может быть увеличено в растении табака, например, в модифицированном растении табака. Соответственно, содержание сложного эфира сахарозы может быть увеличено в листе табака (например, листе табака из модифицированного растения табака). Соответственно, содержание сложного эфира сахарозы может быть увеличено в собранном листе (например, в собранном листе табака из модифицированного растения табака). Соответственно, содержание сложного эфира сахарозы может быть увеличено в нарезанном собранном листе (например, нарезанном собранном листе табака с модифицированного растения табака). Соответственно, содержание сложного эфира сахарозы может быть увеличено в обработанном листе табака (например, обработанном листе табака из модифицированного растения табака). Соответственно, содержание сложного эфира сахарозы может быть увеличено в нарезанном и обработанном листе табака (например, нарезанном и обработанном листе табака из модифицированного растения табака). Соответственно, содержание сложного эфира сахарозы может быть увеличено в экссудате трихом сушеного листа табака (например, листа табака из модифицированного растения табака). Соответственно, содержание сложного эфира сахарозы может быть увеличено в экстракте листа зеленого табака (например, листа табака из модифицированного растения табака). Соответственно, содержание сложного эфира сахарозы может быть увеличено в табачном продукте (например, в табачном продукте, полученном из модифицированного растения табака или его части). Соответственно, содержание сложного эфира сахарозы может быть увеличено в любом из вышеуказанных продуктов или их комбинациях.
В одном воплощении содержание сложного эфира сахарозы увеличивается в экссудате трихом сушеного листа табака (например, листа табака из модифицированного растения табака).
В одном воплощении содержание сложного эфира сахарозы увеличивается в экстракте листа зеленого табака (например, листа табака из модифицированного растения табака).
В одном воплощении содержание сложного эфира сахарозы в растении табака или его части может быть увеличено, по меньшей мере, в 2, 3, 4, 5, 6, 7, 8, 9, 10 раз по сравнению с содержанием сложного эфира сахарозы в растении табака или его части, соответственно, которая не была модифицирована для ингибирования активности или экспрессии гена синтеза дитерпена, который был выращен в аналогичных условиях роста. Соответственно, содержание сложного эфира сахарозы может быть увеличено от 2 до 10 раз, предпочтительно от 3 до 10 раз, предпочтительно от 3 до 5 раз.
В одном воплощении способ или применение приводят к увеличению содержания сложного эфира сахарозы по сравнению с растением табака или его частью, которое не было модифицировано для ингибирования активности или экспрессии гена синтеза дитерпена, и, более конкретно, по сравнению с или относительно экспрессии растения табака при отсутствии введенного ингибирования.
В одном воплощении растение табака или его часть было модифицировано для достижения увеличения содержания сложного эфира сахарозы по сравнению с растением табака или его частью, соответственно, которое не было модифицировано для ингибирования активности или экспрессии гена синтеза дитерпена.
Используемый в данном документе термин «модифицирующий» или «модифицированный» означает растение табака, которое было преобразовано или изменено. Настоящее изобретение включает модификацию растений с применением методов генетической модификации растений или негенетической модификации растений. Такие способы хорошо известны в данной области техники, и примеры методов генетической модификации включают трансформацию, трансгенику, цисгенику и способы редактирования генов. Пример негенетических методов модификации включают мутагенез быстрыми нейтронами, химический мутагенез и современные подходы популяционного анализа. В одном воплощении термин «модификация» относится к выбору природного варианта, который имеет ингибированный ген синтеза дитерпена, и к селекции этого признака или гена во второе растение, которое имеет коммерчески желательные признаки.
Используемый в данном документе термин «ингибирование» (например, ингибирование активности или экспрессии гена синтеза дитерпена) означает, что активность или экспрессия гена синтеза дитерпена ниже или ниже по сравнению с активностью гена или экспрессией гена в сопоставимом продукте или ниже количество или активность белка, продуцируемого геном синтеза дитерпена.
В одном воплощении термин «ингибирование» (например, ингибирование активности или экспрессии гена синтеза дитерпена) в контексте настоящего описания означает, что активность или экспрессия гена синтеза дитерпена ниже по сравнению с активностью гена или экспрессией гена в сопоставимом продукте.
«Активность» или «функция» гена синтеза дитерпена связана с его способностью функционировать в биосинтезе дитерпенов. Активность или функцию гена синтеза дитерпенов можно определить путем измерения прямых продуктов синтеза дитерпенов, то есть путем измерения уровня дитерпенов. Компоненты экссудата могут быть измерены путем промывки листьев или листовых дисков ацетонитрилом. Промывки концентрируют с помощью роторного испарения с получением маслянистого остатка. Затем остаток может быть дериватизирован с образованием сложных эфиров триметилсилила (TMS) (как описано Severson et al. 1985. J. Agric. Food Chem. 33, 870-875, который включен в настоящий документ ссылкой). Затем производные ТМС могут быть разделены и проанализированы газовой хроматографией в сочетании с масс-спектрометрией (GC-MS). Элюированные соединения могут быть идентифицированы по времени их удерживания и по сравнению профилей MS со стандартами, как описано в Wang et al. 2001 (см. выше, который включен в данное описание ссылкой). В одном воплощении прямым продуктом гена синтеза дитерпена могут представлять собой α-CBT-олы, β-CBT-олы, α-CBT-диолы, β-CBT-диолы или лабдендиол.
Активность специфических генов синтеза дитерпена можно измерить путем измерения транскрипции гена. Способы измерения транскрипции хорошо известны в данной области и включают, среди прочего, нозерн-блот, РНК-Seq, гибридизацию in situ, ДНК-микрочипы и ОТ-ПЦР. Альтернативно, активность гена может быть измерена косвенно путем измерения уровня продукта гена, например, белка, кодируемого указанным геном.
Активность или экспрессия гена может относиться к уровню транскрипции, трансляции, т.е. экспрессии белка, или активности белка, кодируемого геном синтеза дитерпена. Согласно одному аспекту изобретения экспрессия гена может быть ингибирована путем ингибирования транскрипции и/или трансляции. В одном воплощении активность или экспрессия гена могут относиться к уровню транскрипции, то есть количеству продуцированной мРНК, или трансляции, т.е. уровню или количеству продуцируемого белка.
Ингибирование, уменьшение или предотвращение экспрессии и/или функции
Любой способ, известный в данной области техники, для ингибирования или уменьшения или предотвращения экспрессии или функции гена синтеза дитерпена может быть использован в настоящем способе.
В качестве примера, настоящий способ может включать:
- обеспечение мутации в последовательности нуклеиновой кислоты, которая кодирует белок, содержащий аминокислотную последовательность, представленную как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11, или аминокислотную последовательность, которая имеет, по меньшей мере, 70% идентичность последовательности с ней;
- обеспечение мутации в регуляторной области (например, промотора и энхансера), которая способствует контролю экспрессии белка, содержащего аминокислотную последовательность, представленную как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11 или аминокислотную последовательность, которая имеет, по меньшей мере, 70% идентичности последовательности с ними;
- обеспечение антисмысловой РНК, siRNA или miRNA, которая снижает уровень последовательности нуклеиновой кислоты, кодирующей белок, содержащий аминокислотную последовательность, представленную как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11, или аминокислотную последовательность, которая имеет, по меньшей мере, 70% идентичности последовательности с ними.
Каждый из вышеуказанных подходов приводит к снижению или предотвращению экспрессии или функции белка, содержащего аминокислотную последовательность, представленную как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11, или аминокислотную последовательность, которая имеет, по меньшей мере, 70% идентичности последовательности с ними.
Используемый в данном документе термин «мутация» охватывает природный генетический вариант или сконструированный вариант. В частности, термин «мутация» относится к изменению в нуклеотидной последовательности, кодирующей аминокислотную последовательность или в аминокислотной последовательности, по сравнению с последовательностью, представленной как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11 или аминокислотной последовательности, которая имеет, по меньшей мере, 70% идентичности последовательности с ними, которое снижает экспрессию или функцию белка.
В предпочтительном воплощении каждая копия последовательности нуклеиновой кислоты, кодирующей белок, содержащий последовательность, представленную как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11, или последовательность, которая имеет, по меньшей мере, 70% идентичности последовательности с ними, которая присутствует в растении, мутирует, как определено в настоящем документе (например, каждая геномная копия гена, кодирующего указанный белок в растении, является мутированной). Например, каждая копия гена в аллотетраплоидном геноме N. tabacum может быть мутирована.
В предпочтительном воплощении растение или растительная клетка согласно настоящему изобретению являются гомозиготными по мутации.
В одном воплощении предпочтительно, чтобы растение или растительная клетка согласно настоящему изобретению экспрессировали только мутированную нуклеиновую кислоту. Другими словами, в некоторых воплощениях эндогенный (или эндогенный и функциональный белок) не присутствует в растении в соответствии с настоящим изобретением. Другими словами, если присутствует какой-либо эндогенный белок, он предпочтительно находится в неактивной и/или усеченной форме.
В одном воплощении настоящий способ может включать обеспечение мутации в последовательности, представленной как SEQ ID No. 1, SEQ ID No. 3 или SEQ ID No. 8, или последовательности нуклеиновой кислоты, которая имеет, по меньшей мере, 70% идентичности с ними.
Мутация может изменить геном растения так, что последовательность нуклеиновой кислоты, кодирующая белок, содержащий аминокислотную последовательность, представленную как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11, или аминокислотную последовательность, которая имеет, по меньшей мере, 70% идентичности с ними, полностью или частично удаляется или иным образом становится нефункциональной.
Мутация может прерывать последовательность нуклеиновой кислоты, которая кодирует белок, содержащий аминокислотную последовательность, представленную как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11, или аминокислотную последовательность, которая имеет идентичность последовательности, по меньшей мере, 70%.
Прерывание может привести к тому, что последовательность нуклеиновой кислоты не будет транскрибироваться и/или транслироваться.
Последовательность нуклеиновой кислоты может быть прервана, например, путем делеции или иной модификации стартового кодона ATG последовательности нуклеиновой кислоты, так что трансляция белка снижается или предотвращается.
Последовательность нуклеиновой кислоты может содержать одно или несколько нуклеотидных изменений, которые уменьшают или предотвращают экспрессию белка или влияют на транспорт белка. Например, экспрессия белка может быть уменьшена или предотвращена путем введения одного или нескольких стоп-кодонов предварительного созревания, сдвига рамки, мутантного сплайсинга или неприемлемой аминокислотной замены в открытой рамке считывания.
Стоп-кодон предварительного созревания относится к мутации, которая вводит стоп-кодон в открытую рамку считывания и предотвращает трансляцию всей аминокислотной последовательности. Стоп-кодон предварительного созревания может представлять собой кодон TAG («янтарь»), TAA («охра») или TGA («опал» или «умбра»).
Мутация со сдвигом рамки (также называемая ошибкой рамки или сдвигом рамки считывания) - это мутация, вызванная инделами (вставками или делециями) ряда нуклеотидов в последовательности нуклеиновой кислоты, которые не делятся на три. Из-за триплетной природы экспрессии генов кодонами, вставка или делеция может изменить рамку считывания, что приведет к совершенно другой трансляции из оригинала. Мутация смещения рамки считывания часто вызывает считывание кодонов после мутации с кодированием различных аминокислот. Мутация смещения рамки считывания обычно приводит к введению преждевременного стоп-кодона.
Мутант сплайсинга вставляет, удаляет или изменяет ряд нуклеотидов в определенном сайте, в котором происходит сплайсинг во время процессинга предшественника информационной РНК в зрелую информационную РНК. Удаление сайта сплайсинга приводит к тому, что один или несколько интронов остаются в зрелой мРНК и могут приводить к образованию аномальных белков.
Непереносимая аминокислотная замена относится к мутации, которая вызывает несинонимичную аминокислотную замену в белке, что приводит к снижению или абляции функции белка.
Любой способ, известный в данной области техники для обеспечения мутации в последовательности нуклеиновой кислоты, может быть использован в настоящем способе. Например, может быть использована гомологичная рекомбинация, при которой создается вектор, в которой соответствующие последовательности нуклеиновой кислоты мутированы и используются для трансформации растений или растительных клеток. Затем могут быть отобраны рекомбинантные растения или растительные клетки, экспрессирующие мутированную последовательность.
В одном воплощении мутация вводит недопустимую аминокислотную замену в белке, содержащем аминокислотную последовательность, представленную как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11, или последовательность, которая имеет, по меньшей мере, 70% идентичности с ними.
В одном воплощении мутация снижает активность белка по отношению к белку, показанному как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11, или последовательности, которая имеет, по меньшей мере, 70% идентичности последовательности с ними.
В одном воплощении мутация не изменяет уровень или экспрессию, но снижает активность белка по отношению к белку, показанному как SEQ ID No. 2, SEQ ID No. 4 или SEQ ID No. 11, или последовательности, которая имеет, по меньшей мере, 70% идентичности последовательности с ними.
Последовательность нуклеиновой кислоты может быть полностью или частично удалена. Делеция может быть непрерывным или может содержать множество участков последовательности. Эта делеция предпочтительно удаляет достаточное количество нуклеотидной последовательности, так что последовательность нуклеиновой кислоты больше не кодирует функциональный белок. Делеция может, например, удалять, по меньшей мере, 50, 60, 70, 80 или 90% кодирующей части последовательности нуклеиновой кислоты.
Эта делеция может удалить один или несколько доменов гена синтеза дитерпена. Эта делеция может быть полной, и в этом случае 100% кодирующей части последовательности нуклеиновой кислоты отсутствует по сравнению с соответствующим геномом сопоставимого немодифицированного растения.
Способы удаления последовательностей нуклеиновых кислот в растениях известны в данной области. Например, может использоваться гомологичная рекомбинация, в которой создается вектор, в котором отсутствуют соответствующие последовательности нуклеиновой кислоты, и он используется для трансформации растений или растительных клеток. Затем могут быть отобраны рекомбинантные растения или растительные клетки, экспрессирующие новую часть последовательности.
В некоторых воплощениях активность или экспрессия гена синтеза дитерпена могут быть ингибированы или уменьшены, по меньшей мере, на около 10%, 20%, 30% или 40%, предпочтительно, по меньшей мере, на около 50%, 60%, 70%, более предпочтительно, по меньшей мере, на около 80%, 90%, 95% или 100% по сравнению с активностью или экспрессией гена синтеза дитерпена в растении табака, которое не было модифицировано в соответствии с настоящим изобретением.
В предпочтительном воплощении ген синтеза дитерпена может практически не иметь активности или экспрессии, что означает, что растение может иметь активность или экспрессию менее чем около 1% (соответственно менее чем около 0,1%), предпочтительно по сравнению с растением, которое не было модифицированный для ингибирования активности или экспрессии гена синтеза дитерпена.
Используемый в настоящем описании ген синтеза дитерпена относится к любому гену, который участвует в производстве дитерпенов. Подходящим геном синтеза дитерпена является ген синтеза растительного дитерпена. Такие гены могут быть непосредственно частью пути биосинтеза, который продуцирует дитерпены, или могут косвенно снабжать путь биосинтеза, например, путем обеспечения метаболических предшественников. Гены синтеза дитерпенов могут участвовать в специализированном (вторичном) метаболизме или общем (первичном) метаболизме. В одном воплощении ген синтеза дитерпена, используемый в данном документе, относится к гену, который участвует в специализированном (вторичном) метаболизме. Соответственно, ген синтеза дитерпена, используемый в настоящем изобретении, может быть выбран из группы генов синтеза дитерпена, состоящей из гена циклазы 2 (CYC2), CBTol-циклазы и терпен-синтазы 3-8 или гомологичной любому из числа гена циклазы 2 (CYC2).), CBTol-циклазы или терпен-синтазы 3-8. Соответственно, ген синтеза дитерпена, используемый в настоящем изобретении, может быть выбран из группы генов синтеза дитерпена, состоящей из гена циклазы 2 (CYC2), CBTol-циклазы и терпен-синтазы 3-8.
Подходящим геном синтеза дитерпена, который используется в настоящем изобретении, является ген циклазы 2 (CYC2). Подходящим геном синтеза дитерпена, который используется в настоящем изобретении, является CBTol-циклаза. Подходящим геном синтеза дитерпена, который используется в настоящем изобретении, является терпенсинтаза 3-8.
В одном воплощении ген синтеза дитерпена, на который ссылаются в данном документе, представляет собой ген циклазы 2 (CYC2) и может кодироваться полинуклеотидной последовательностью, содержащей:
i) полинуклеотидную последовательность, представленную в данном документе как SEQ ID No. 8 или SEQ ID No. 9, или
ii) функциональный фрагмент полинуклеотидной последовательности, показанной на i), причем этот функциональный фрагмент кодирует ген синтеза дитерпена, или
iii) полинуклеотид, который кодирует полипептид, содержащий аминокислотную последовательность, представленную в данном документе как SEQ ID No. 10, или
iv) полинуклеотидную последовательность, которая может гибридизоваться с полинуклеотидом, описанным в пунктах i), ii) или iii) выше, в условиях высокой жесткости, или
v) полинуклеотидную последовательность, которая имеет, по меньшей мере, 70% (предпочтительно 85%, более предпочтительно 90%) идентичности с полинуклеотидом, представленным в i), ii) или iii) выше, или
vi) полинуклеотидную последовательность, которая отличается от полинуклеотида, представленного в i), ii) или iii), из-за вырожденности генетического кода.
В одном воплощении ген синтеза дитерпена, на который ссылаются в данном документе, представляет собой CBTol-циклазу и может кодироваться полинуклеотидной последовательностью, содержащей:
i) полинуклеотидную последовательность, представленную в данном документе как SEQ ID No. 1, или
ii) функциональный фрагмент полинуклеотидной последовательности, представленной в i), причем этот функциональный фрагмент кодирует ген синтеза дитерпена, или
iii) полинуклеотид, который кодирует полипептид, содержащий аминокислотную последовательность, представленную в данном документе как SEQ ID No. 2, или
iv) полинуклеотидную последовательность, которая может гибридизоваться с полинуклеотидом, описанным в пунктах i), ii) или iii) выше, в условиях высокой жесткости, или
v) полинуклеотидную последовательность, которая имеет, по меньшей мере, 70% (предпочтительно 85%, более предпочтительно 90%) идентичности с полинуклеотидом, представленным в i), ii) или iii) выше, или
vi) полинуклеотидную последовательность, которая отличается от полинуклеотида, представленного в i), ii) или iii), из-за вырожденности генетического кода.
В одном воплощении ген синтеза дитерпена, на который ссылаются в данном документе, представляет собой терпен-синтазу 3-8 и может кодироваться полинуклеотидной последовательностью, содержащей:
i) полинуклеотидную последовательность, представленную в данном документе как SEQ ID No. 3, или
ii) функциональный фрагмент полинуклеотидной последовательности, представленной в i), причем этот функциональный фрагмент кодирует ген синтеза дитерпена, или
iii) полинуклеотид, который кодирует полипептид, содержащий аминокислотную последовательность, представленную в данном документе как SEQ ID No. 4, или
iv) полинуклеотидную последовательность, которая может гибридизоваться с полинуклеотидом, описанным в пунктах i), ii) или iii) выше, в условиях высокой жесткости, или
v) полинуклеотидную последовательность, которая имеет, по меньшей мере, 70% (предпочтительно 85%, более предпочтительно 90%) идентичности с полинуклеотидом, представленным в i), ii) или iii) выше, или
vi) полинуклеотидную последовательность, которая отличается от полинуклеотида, представленного в i), ii) или iii), из-за вырожденности генетического кода.
В одном воплощении ген синтеза дитерпена для применения в соответствии с настоящим изобретением может быть эндогенным для растения табака.
Ссылка в данном документе на «эндогенный» ген не только относится к рассматриваемому гену, обнаруженному в растении в его естественной форме (т.е. без какого-либо вмешательства человека), но также относится к тому же гену (или по существу гомологичной нуклеиновой кислоте/гену) в выделенной форме, впоследствии (повторно) внедренной в растение (трансген). Например, трансгенное растение, содержащее такой трансген, может столкнуться со значительным снижением экспрессии трансгена и/или существенным снижением экспрессии эндогенного гена. Выделенный ген может быть выделен из организма или может быть создан человеком, например, химическим синтезом.
В другом воплощении ген синтеза дитерпена для применения в соответствии с настоящим изобретением может быть экзогенным для растения табака.
Чтобы определить, является ли ген геном синтеза дитерпена для применения в соответствии с настоящим изобретением, специалист может определить, способен ли данный ген продуцировать дитерпены. Можно измерить содержание дитерпенов, как описано выше в настоящем документе. Вкратце, способ измерения содержания дитерпена может включать сбор экссудата с листа или листьев путем промывки ацетонитрилом листьев или дисков из листьев. Остаток от промывок затем может быть дериватизирован с образованием сложных эфиров триметилсилила (TMS) (как описано Severson et al. 1985, см. выше, который включен в данное описание ссылкой). Затем производные TMS могут быть разделены и проанализированы газовой хроматографией в сочетании с масс-спектрометрией (GC-MS). Элюированные соединения можно идентифицировать по времени их удерживания и сопоставлению профилей МС со стандартами.
Настоящее изобретение также предусматривает применение гена синтеза дитерпена для увеличения содержания сложных эфиров сахарозы в растении.
Способы снижения экспрессии генов или генных продуктов хорошо документированы в данной области.
В одном воплощении активность или экспрессию гена синтеза дитерпена можно ингибировать любым способом, известным в данной области. Соответственно, активность или экспрессия гена синтеза дитерпена, выбранного из группы генов синтеза дитерпена, состоящей из гена циклазы 2 (CYC2), CBTol-циклазы и терпен-синтазы 3-8, может быть ингибирована любым способом, известным в данной области. Соответственно, активность или экспрессия гена циклазы 2 (CYC2) может быть ингибирована любым способом, известным в данной области. Соответственно, активность или экспрессия CBTol-циклазы может быть ингибирована любым способом, известным в данной области. Соответственно, активность или экспрессия терпенсинтазы 3-8 могут быть ингибированы любым способом, известным в данной области.
Способы ингибирования активности или экспрессии гена синтеза дитерпена могут включать РНК-интерференцию, антисмысловую или смысловую косупрессию (см. Wang and Wagner 2003, Planta Volume 216, Issue 4, pp 686–691, который включен в настоящий документ ссылкой) редактирование генов или направленный мутагенез. В одном воплощении ингибирование активности или экспрессии гена синтеза дитерпена может быть достигнуто с помощью редактирования генов. Редактирование генов может быть выполнено с применением любого способа, известного в данной области. Несколько неограничивающих примеров представлены в данном документе.
В одном воплощении ингибирование активности или экспрессии гена синтеза дитерпена может быть достигнуто с применением способов редактирования генов, включая CRISPR, включая применение системы CRISPR-Cas9. Инструменты геномного редактирования CRISPR/Cas9 доступны на рынке, такие как «Guide-it» от Clontech (Avenue du President Kennedy 78100 Saint-Germain-en-Laye, Франция).
Другой способ редактирования генов включает применение технологии TALEN (эффекторная нуклеаза, подобная активаторам транскрипции) с коммерчески доступными наборами (например, от Addgene, 1Kendall Sq. Ste. B7102, Кембридж, MA 02139, США). В одном воплощении ингибирование активности или экспрессии гена синтеза дитерпена может быть достигнуто с применением TALEN.
В другом воплощении способ может включать применение нуклеаз с цинковыми пальцами, например, как технология нуклеаз с цинковыми пальцами CompoZr®, доступная от Sigma-Aldrich. Другое воплощение может включать применение мегануклеаз (или другого способа), описанных в Silva et al. Curr Gene Ther. Feb 2011; 11(1): 11–27 (описание которого включено в данное описание ссылкой).
В одном воплощении способ ингибирования активности или экспрессии гена синтеза дитерпена может представлять собой направленный мутагенез. Может быть использован любой способ направленного мутагенеза. В одном воплощении способ может представлять собой олигонуклеотид-направленный мутагенез (ODM), такой как KeyBase®, доступный от Keygene (Agro Business Park 90, 6708 PW Wageningen, Нидерланды). В другом воплощении ингибирование активности или экспрессии гена синтеза дитерпена может быть достигнуто путем использования конструкции или вектора (например, плазмиды).
Генетические конструкции по изобретению могут быть в форме кассеты экспрессии, которая может быть подходящей для ингибирования активности или экспрессии гена синтеза дитерпена в клетке-хозяине. Генетическая конструкция может быть введена в клетку-хозяина без ее включения в вектор. Например, генетическая конструкция, которая может представлять собой молекулу нуклеиновой кислоты, может быть включена в липосому или вирусную частицу. Альтернативно, очищенная молекула нуклеиновой кислоты (например, не содержащая гистонов ДНК или оголенная ДНК) может быть вставлена непосредственно в клетку-хозяина подходящими способами, например, прямым эндоцитозным захватом. Генетическая конструкция может быть введена непосредственно в клетки объекта-хозяина (например, растения) путем трансфекции, инфекции, микроинъекции, слияния клеток, слияния протопластов или баллистической бомбардировки. Альтернативно, генетические конструкции по изобретению могут быть введены непосредственно в клетку-хозяина с применением генной пушки.
Альтернативно, генетическая конструкция может содержать или находиться в рекомбинантном векторе для экспрессии в подходящей клетке-хозяине. Рекомбинантный вектор может представлять собой плазмиду, космиду или фаг. Такие рекомбинантные векторы очень полезны для трансформации клеток-хозяев генетической конструкцией по изобретению и для репликации в них экспрессирующей кассеты. Специалист в данной области поймет, что генетические конструкции по изобретению можно комбинировать со многими типами каркасного вектора для целей экспрессии. Основным вектором может быть бинарный вектор, например, который может реплицироваться как в E.coli, так и в Agrobacterium tumefaciens. Например, подходящим вектором может быть плазмида pBIN, такая как pBIN19 (Bevan M., 1984, Nucleic Acids Research 12: 8711-21).
Рекомбинантные векторы могут включать множество других функциональных элементов в дополнение к последовательности, которая ингибирует активность или экспрессию гена синтеза дитерпена. Например, вектор может содержать промотор. Кроме того, рекомбинантный вектор может быть сконструирован таким образом, что он автономно реплицируется в цитозоле клетки-хозяина. В этом случае элементы, которые индуцируют или регулируют репликацию ДНК, могут потребоваться в рекомбинантном векторе. Альтернативно, рекомбинантный вектор может быть сконструирован таким образом, что он интегрируется в геном клетки-хозяина. В этом случае предусматриваются последовательности ДНК, которые способствуют направленной интеграции (например, путем гомологичной рекомбинации).
Рекомбинантный вектор также может содержать ДНК, кодирующую ген, который можно использовать в качестве селективного маркера в процессе клонирования, то есть, позволяющего осуществить отбор клеток, которые были трансфицированы или трансформированы, и позволяющего осуществить отбор клеток, содержащих векторы, включающие гетерологичную ДНК. Вектор также может содержать ДНК, участвующую в регуляции экспрессии кодирующей последовательности или для нацеливания экспрессированного полипептида на определенную часть клетки-хозяина, например, на трихомы или железистые трихомы. Следовательно, вектор может включать, по меньшей мере, один дополнительный элемент, выбранный из группы, состоящей из: селектируемого маркерного гена (например, гена устойчивости к антибиотику); сигнал терминации полипептида; и белковой нацеливающей последовательности (например, транзитного пептида).
В одном воплощении способ или применение могут включать ингибирование активности или экспрессии гена синтеза дитерпена с применением RNAi. В одном воплощении в способе RNAi используется miRNA, например, искусственная microRNA (amiRNA). В одном воплощении в способе RNAi используется siRNA. В одном воплощении в способе RNAi используется dsRNA.
В одном воплощении способ или применение могут включать ингибирование активности или экспрессии гена синтеза дитерпена с применением интерферирующего олигонуклеотида. В одном воплощении олигонуклеотид основан на РНК. В одном воплощении олигонуклеотид представляет собой РНК-интерференцию (RNAi), например, dsRNAi. В одном воплощении способ может включать трансформацию клетки растения табака с помощью молекулы RNAi, например, dsRNAi, которая ингибирует активность или экспрессию гена синтеза дитерпена.
В одном воплощении предлагаются растения табака и материалы для размножения растений табака, в которых экспрессия гена синтеза дитерпена ингибируется с применением RNAi (например, dsRNAi). Способ может включать регенерацию растения табака из трансформированной клетки.
Молекула RNAi (например, dsRNAi) может быть способна снижать в трансформированном растении активность или экспрессию гена синтеза дитерпена, по меньшей мере, на 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или более, или на 100% по сравнению с концентрацией полипептида в растении дикого типа, который не был трансформирован с помощью молекулы RNAi. В одном воплощении молекула RNAi может быть способна уменьшать в трансформированном растении активность или экспрессию гена синтеза дитерпена на 30-100%, предпочтительно на 40-100%, более предпочтительно на 90-100%.
Активность или экспрессия гена синтеза дитерпена можно ингибировать любым способом, известным в данной области. Соответственно, активность или экспрессия гена синтеза дитерпена, выбранного из группы генов синтеза дитерпена, состоящей из гена циклазы 2 (CYC2), CBTol-циклазы и терпен-синтазы 3-8, может быть ингибирована любым способом, известным в данной области.
Соответственно, способ может включать трансформацию клетки растения табака с помощью ДНК-конструкции ddRNAi, которая кодирует РНК, которая образует шпилечную структуру, которая обрабатывается эндогенными путями в клетке, в малую или короткую интерферирующую РНК (siRNA).
Соответственно, активность или экспрессия гена циклазы 2 (CYC2) может быть ингибирована любым способом, известным в данной области. В одном воплощении последовательность гена циклазы 2 (CYC2) является такой, как представленная в SEQ ID No. 8. В одном воплощении аминокислотная последовательность гена циклазы 2 (CYC2) является такой, как представленная в SEQ ID No. 10. Экспрессия гена циклазы 2 (CYC2) может быть ингибирована любым методом, включая методы редактирования генов, включая CRISPR, включая применение системы CRISPR-Cas9, интерференцию РНК (RNAi), антисмысловую или смысловую косупрессию, редактирование генов или направленный мутагенез. В одном воплощении активность или экспрессия гена циклазы 2 (CYC2) может быть ингибирована с помощью RNAi. Активность или экспрессия гена циклазы 2 (CYC2) может быть ингибирована с помощью RNAi с применением miRNA, siRNA, dsRNA или shRNA.
В одном воплощении способ ингибирования активности или экспрессии гена циклазы 2 (CYC2) направлен, по меньшей мере, на часть экзона 1, по меньшей мере, на часть экзона 2 и, по меньшей мере, на часть экзона 3 гена циклазы 2 (CYC2). В предпочтительном воплощении способ нацелен, по меньшей мере, на часть экзона 1, по меньшей мере, на часть экзона 2 и, по меньшей мере, часть экзона 3 гена циклазы 2 (CYC2). В одном воплощении способ нацелен, по меньшей мере, на часть экзона 1, по меньшей мере, на часть экзона 2 и первые 115 нуклеотидов 3 экзона гена циклазы 2 (CYC2). В одном воплощении способ нацелен на нуклеотиды 1-25, нуклеотиды 26-271 (экзон 1), нуклеотиды 1253-1529 (экзон 2) и первые 115 нуклеотидов 3 экзона (нуклеотиды 2366-2480) гена циклазы 2 (CYC2), где нумерация определяется путем выравнивания с SEQ ID No. 8.
В любом из предыдущих воплощений активность или экспрессию гена циклазы 2 (CYC2) можно ингибировать с применением способов редактирования генов, включая CRISPR, включая применение системы CRISPR-Cas9. В любом из предыдущих воплощений активность или экспрессию гена циклазы 2 (CYC2) можно ингибировать с применением способа RNAi.
Соответственно, активность или экспрессия гена CBTol-циклазы может быть ингибирована любым способом, известным в данной области. В одном воплощении последовательность гена CBTol-циклазы является такой, как представленная в SEQ ID No. 1. В одном воплощении аминокислотная последовательность CBTol-циклазы является такой, как представленная в SEQ ID No. 2. Экспрессия гена CBTol-циклазы может быть ингибирована любым способом, включая способы редактирования генов, включая CRISPR, включая применение системы CRISPR-Cas9, интерференцию РНК (RNAi), антисмысловую или смысловую косупрессию, редактирование генов или направленный мутагенез. В одном воплощении активность или экспрессия гена CBTol-циклазы может быть ингибирована с помощью RNAi. Активность или экспрессия гена CBTol-циклазы может быть ингибирована с помощью RNAi с применением miRNA, siRNA, dsRNA или shRNA.
В другом воплощении способ ингибирования активности или экспрессии гена CBTol-циклазы направлен, по меньшей мере, на часть экзона 4, интрона 4, экзона 5, интрона 5, экзона 6, интрона 6 и, по меньшей мере, части экзона 7 CBTol-циклазы. В предпочтительном воплощении способ нацелен, по меньшей мере, на часть экзона 4, интрона 4, экзона 5, интрона 5, экзона 6, интрона 6 и, по меньшей мере, часть экзона 7 гена CBTol-циклазы. В одном воплощении способ нацелен на нуклеотиды 5'-2854-4175-3' гена CBTol-циклазы, где нумерация определяется выравниванием с SEQ ID No. 1.
В любом из предыдущих воплощений активность или экспрессию гена CBTol-циклазы можно ингибировать с применением способов редактирования генов, включая CRISPR, включая применение системы CRISPR-Cas9. В любом из предыдущих воплощений активность или экспрессию гена CBTol-циклазы можно ингибировать с применением способа RNAi.
Соответственно, активность или экспрессия гена терпен-синтазы 3-8 может быть ингибирована любым способом, известным в данной области. В одном воплощении последовательность гена терпен-синтазы 3-8 является такой, как представленная в SEQ ID No. 3. В одном воплощении аминокислотная последовательность терпенсинтазы 3-8 является такой, как представленная в SEQ ID No. 4. Экспрессия гена терпен-синтазы 3-8 может быть ингибирована любым способом, включая способы редактирования генов, включая CRISPR, включая применение системы CRISPR-Cas9, интерференцию РНК (RNAi), антисмысловую или смысловую косупрессию, редактирование генов или направленный мутагенез. В одном воплощении активность или экспрессия гена терпен-синтазы 3-8 могут быть ингибированы с помощью RNAi. Активность или экспрессия гена терпен-синтазы 3-8 может быть ингибирована с помощью RNAi с применением miRNA, siRNA, dsRNA или shRNA. В одном воплощении способ ингибирования активности или экспрессии гена терпен-синтазы 3-8 нацелен, по меньшей мере, на нуклеотиды с 1497 по 1517 гена терпен-синтазы 3-8, где нумерация определяется выравниванием с SEQ ID No. 3. В одном воплощении способ ингибирования активности или экспрессии гена терпен-синтазы 3-8 нацелен, по меньшей мере, на нуклеотиды с 884 по 904 гена терпен-синтазы 3-8, где нумерация определяется выравниванием с SEQ ID No. 3. В предпочтительном воплощении способ RNAi нацелен на нуклеотиды с 884 по 904 гена терпен-синтазы 3-8, где нумерация определяется выравниванием с SEQ ID No. 3.
В любом из предыдущих воплощений активность или экспрессию гена терпен-синтазы 3-8 можно ингибировать с применением способов редактирования генов, включая CRISPR, включая применение системы CRISPR-Cas9. В любом из предыдущих воплощений активность или экспрессию гена терпен-синтазы 3-8 можно ингибировать с применением метода RNAi.
В одном воплощении способ может включать трансформацию клетки растения табака с помощью конструкции ДНК-направленной РНК-интерференции (ddRNAi), которая включает нуклеотидную последовательность, которая кодирует dsRNA, или нуклеотидную последовательность, которая кодирует amiRNA, упакованную в вектор доставки.
В одном воплощении молекула RNAi представляет собой dsRNA, которая кодируется ДНК-конструкцией ddRNAi.
В одном воплощении молекула RNAi представляет собой amiRNA, которая кодируется ДНК-конструкцией ddRNAi. В одном воплощении настоящее изобретение относится к конструкции, содержащей последовательность ДНК ddRNAi, которая предназначена для кодирования dsRNA, которая ингибирует экспрессию гена синтеза дитерпена.
В другом воплощении настоящее изобретение относится к конструкции, содержащей последовательность ДНК ddRNAi, которая предназначена для кодирования amiRNA, которая ингибирует экспрессию гена синтеза дитерпена.
Конструкция может содержаться в векторе. Подходящим вектором может быть плазмида.
В одном воплощении вектор для применения в настоящем изобретении представляет собой плазмиду на основе Agrobacterium pKYLX71:35S2. Эта плазмида основана на плазмиде pGA471, описанной An et al. (An, G. et al. 1985, EMBO J. 4, 277-284, который включен в данное описание ссылкой). В другом воплощении вектор для применения в настоящем изобретении представляет собой pCAMBIA2300. Основная цепь вектора pCAMBIA получена из векторов pPZP.
Используемый в данном документе термин «ДНК-направленная РНК-интерференция (ddRNAi)» означает ДНК-конструкцию, которая используется для активации путей эндогенной РНК-интерференции (РНКi) клетки. Соответственно, эти конструкции предназначены для экспрессии самокомплементарных РНК, например, двухцепочечной РНК (dsRNA) или одноцепочечной РНК (ssRNA) или коротких шпилечных РНК (shRNA) или микроРНК (например, искусственной микроРНК - amiRNA), которые после обработки вызывают сайленсинг целевого гена или генов. Преимущественно применение ddRNAi означает, что экспрессированная РНК (например, dsRNA или amiRNA) непрерывно продуцируется и, таким образом, способна обеспечивать длительный сайленсинг целевых генов. В отличие от этого, небольшая интерферирующая РНК (siRNA), вводимая непосредственно в клетку (например, не экспрессируемая непрерывно с ДНК-конструкции ddRNAi в соответствии с настоящим изобретением), превращается внутри клетки и только временно заглушает гены.
В одном воплощении способ или применение могут включать ингибирование активности или экспрессии гена синтеза дитерпена с применением dsRNA, которая экспрессируется из ДНК-конструкции ddRNAi.
В другом воплощении способ или применение могут включать ингибирование активности или экспрессии гена синтеза дитерпена с применением amiRNA, которая экспрессируется из ДНК-конструкции ddRNAi.
Соответственно, в одном воплощении предусмотрены растения табака и материалы для размножения растений табака, листья табака, срезанные собранные листья, обработанные листья табака или разрезанные и обработанные листья табака, в которых экспрессия гена синтеза дитерпена ингибируется с применением ДНК-конструкции ddRNAi. Соответственно, ДНК-конструкция ddRNAi может быть включена в геномную ДНК растения. Растение табака, материал для размножения растения табака, лист табака, срезанный собранный лист, обработанный табачный лист или нарезанный и обработанный табачный лист могут содержать ДНК-конструкцию ddRNAi, которая экспрессирует dsRNA или amiRNA, которая ингибирует активность или экспрессию гена синтеза дитерпена. Растение табака, материал для размножения растения табака, лист табака, срезанный собранный лист, обработанный лист табака или разрезанный и обработанный лист табака, который содержит конструкцию ДНК ddRNAi, которая экспрессирует dsRNA или amiRNA, которая ингибирует активность или экспрессию гена синтеза дитерпена, может иметь повышенное содержание сложного эфира сахарозы по сравнению с растением табака, материалом для размножения табачных растений, листом табака, срезанным собранным листом, обработанным листом табака или разрезанным и обработанным листом табака, который не содержит ДНК-конструкцию ddRNAi.
ДНК-конструкция ddRNAi может включать весь ген синтеза дитерпена или его часть. Конструкция может содержать экзоны и/или интроны гена синтеза дитерпена. ДНК-конструкция ddRNAi может содержать частичные генные последовательности, которые при транскрибировании образуют шпилечную РНК-структуру. РНК, кодируемая ДНК-конструкцией ddRNAi, может иметь либо структуру интрон-шпилька, либо GUS-шпилька, или может представлять собой одну полноразмерную РНК. Как описано в данном документе, авторы изобретения продемонстрировали удивительную эффективность молекул RNAi, которые ингибируют активность или экспрессию гена синтеза дитерпена для применения в увеличении содержания сложного эфира сахарозы в растениях табака.
Используемый в данном документе термин «экзон» означает часть гена, которая кодирует конечную зрелую РНК, продуцируемую геном после удаления интронов при сплайсинге РНК.
Используемый в данном документе термин «интрон» означает нуклеотидную последовательность в гене, которая удаляется путем сплайсинга РНК во время созревания конечного продукта РНК.
Термин «по меньшей мере, часть» или «частичная последовательность», используемый в данном документе, означает последовательность, включающую, по меньшей мере, 5, 10, 15, 20, 25, 30, 40, 45, 50, 60, 70, 80, 90, 100. 120, 140, 160, 180, 200, 220, 240, 260, 280, 300, 400 или, по меньшей мере, 500 смежных нуклеотидов. Например, ДНК-конструкция ddRNAi может содержать, по меньшей мере, часть экзона, где эта конструкция содержит 100 смежных нуклеотидов из указанного экзона.
Используемый в данном документе термин «мРНК, соответствующий» означает, что РНК имеет ту же последовательность, что и ДНК, т.е. последовательность нуклеотидов одинакова как в мРНК, так и в последовательности ДНК, за исключением того, что в РНК тимин (Т) заменен урацилом. (U), и дезоксирибоза замещена рибозой.
ДНК-конструкция ddRNAi может быть использована для ингибирования экспрессии гена циклазы 2 (CYC2). В одном воплощении последовательность гена циклазы 2 (CYC2) является такой, как представленная в SEQ ID No. 8. В одном воплощении аминокислотная последовательность полипептида, кодируемого геном циклазы 2 (CYC2), является такой, как представленная в SEQ ID No. 10. Соответственно, ДНК-конструкция ddRNAi может содержать весь или часть гена циклазы 2 (CYC2).
В одном воплощении ДНК-конструкция ddRNAi может содержать, по меньшей мере, часть экзона 1, по меньшей мере, часть экзона 2 и, по меньшей мере, часть экзона 3 гена циклазы 2 (CYC2). Соответственно, ДНК-конструкция ddRNAi может содержать, по меньшей мере, часть экзона 1, по меньшей мере, часть экзона 2 и первые 115 нуклеотидов 3 экзона гена циклазы 2 (CYC2). Соответственно, ДНК-конструкция ddRNAi может содержать нуклеотиды 1-25, нуклеотиды 26-271 (экзон 1), нуклеотиды 1253-1529 (экзон 2) и первые 115 нуклеотидов 3 экзона (нуклеотиды 2366-2480) гена циклазы 2 (CYC2), где нумерация определяется выравниванием с SEQ ID No. 8. Соответственно, ДНК-конструкция ddRNAi может содержать последовательность, приведенную в SEQ ID No. 5.
ДНК-конструкция ddRNAi может быть использована для ингибирования экспрессии гена CBTol-циклазы. В одном воплощении последовательность гена CBTol-циклазы является такой, как представленная в SEQ ID No. 1. В одном воплощении аминокислотная последовательность CBTol-циклазы является такой, как представленная в SEQ ID No. 2. Соответственно, ДНК-конструкция ddRNAi может включать весь или часть гена CBTol-циклазы.
В другом воплощении ДНК-конструкция ddRNAi может содержать, по меньшей мере, часть экзона 4, интрона 4, экзона 5, интрона 5, экзона 6, интрона 6 и, по меньшей мере, часть экзона 7 гена CBTol-циклазы. Соответственно, ДНК-конструкция ddRNAi может содержать нуклеотиды 5'-2854-4175-3' гена CBTol-циклазы, где нумерация определяется выравниванием с SEQ ID No. 1. Соответственно, ДНК-конструкция ddRNAi может содержать последовательность, приведенную в SEQ ID No. 6.
ДНК-конструкция ddRNAi может быть использована для ингибирования экспрессии гена терпен-синтазы 3-8. Соответственно, ДНК-конструкция ddRNAi может включать весь или часть гена терпен-синтазы 3-8. Соответственно, ДНК-конструкция ddRNAi может содержать, по меньшей мере, нуклеотиды с 1497 по 1517 гена терпен-синтазы 3-8, где нумерация определяется выравниванием с SEQ ID No. 3. В одном воплощении последовательность гена терпен-синтазы 3-8 является такой, как представленная в SEQ ID No. 3. В одном воплощении аминокислотная последовательность терпенсинтазы 3-8 является такой, как представленная в SEQ ID No. 4. Соответственно, ДНК-конструкция ddRNAi может содержать последовательность, приведенную в SEQ ID No. 7. Соответственно, ДНК-конструкция ddRNAi может содержать, по меньшей мере, нуклеотиды от 884 до 904 гена терпен-синтазы 3-8, где нумерация определяется путем выравнивания с SEQ ID No. 3. Соответственно, ДНК-конструкция ddRNAi может содержать последовательность, приведенную в SEQ ID No. 11.
Последовательность ДНК-конструкции ddRNAi, которая кодирует dsRNA в одном воплощении, может включать последовательность, изложенную в SEQ ID No. 7. dsRNA, экспрессируемая из ДНК-конструкции ddRNAi, содержащей SEQ ID No. 7, может быть использована для ингибирования активности или экспрессии или для подавления гена синтеза дитерпена, где ген синтеза дитерпена представляет собой ген циклазы 2 (CYC2).
Последовательность ДНК-конструкции ddRNAi, которая кодирует dsRNA в другом воплощении, может включать последовательность, изложенную в SEQ ID No. 8. dsRNA, экспрессируемая ДНК-конструкцией ddRNAi, содержащей SEQ ID No. 8, может быть использована для ингибирования активности или экспрессии или для подавления гена синтеза дитерпена, где ген синтеза дитерпена представляет собой ген CBTol-циклазы.
Последовательность ДНК-конструкции ddRNAi, которая кодирует amiRNA в одном воплощении, может включать последовательность, представленную в SEQ ID No. 9. amiRNA, экспрессируемая из ДНК-конструкции ddRNAi, содержащей SEQ ID No. 9, может быть использована для ингибирования активности или экспрессии или для подавления гена синтеза дитерпена, где ген синтеза дитерпена представляет собой ген терпен-синтазы 3-8.
Используемый в данном документе термин «GW1» или «GW-1» может означать конструкцию ДНК ddRNAi, содержащую, по меньшей мере, часть экзона 1, по меньшей мере, часть экзона 2 и первые 115 нуклеотидов 3 экзона гена циклазы 2 (CYC2).
Используемый в данном документе термин «GW2» или «GW-2» может означать конструкцию ДНК ddRNAi, содержащую, по меньшей мере, нуклеотиды и от 1497 до 1517 гена терпен-синтазы 3-8, где нумерация определяется выравниванием с SEQ ID No. 3.
Используемый в данном документе термин «GW3» или «GW-3» может означать конструкцию ДНК ddRNAi, включающую, по меньшей мере, часть экзона 4, интрона 4, экзона 5, интрона 5, экзона 6, интрона 6 и, по меньшей мере, части экзона 7 ген CBTol-циклазы.
Используемый в данном документе термин «GW5» или «GW-5» может означать конструкцию ДНК ddRNAi, содержащую, по меньшей мере, нуклеотиды от 884 до 904 гена терпен-синтазы 3-8, где нумерация определяется выравниванием с SEQ ID No. 3.
Таким образом, ДНК-конструкция ddRNAi по изобретению может содержать нуклеотидную последовательность, которая кодирует dsRNA, где кодирующая последовательность dsRNA представляет собой нуклеотидную последовательность, по существу, изложенную в SEQ ID No. 5 или 8, или ее функциональный вариант, или его фрагмент.
Соответственно, ДНК-конструкция ddRNAi по изобретению может содержать нуклеотидную последовательность, которая кодирует dsRNA, где последовательность, кодирующая dsRNA, представляет собой нуклеотидную последовательность, в основном такую, как изложена в SEQ ID No. 5, или ее функциональный вариант, или его фрагмент.
Соответственно, конструкция ДНК ddRNAi по изобретению может содержать нуклеотидную последовательность, которая кодирует dsRNA, где кодирующая последовательность dsRNA представляет собой нуклеотидную последовательность, в основном такую, как изложена в SEQ ID No. 6, или ее функциональный вариант, или его фрагмент.
В другом воплощении ДНК-конструкция ddRNAi по изобретению может содержать нуклеотидную последовательность, которая кодирует amiRNA, где последовательность, кодирующая amiRNA, представляет собой нуклеотидную последовательность, по существу, как указано в SEQ ID No. 9.
Понятно, что для специалиста было бы относительно просто модифицировать последовательность, которая кодирует dsRNA (например, SEQ ID No. 7) или amiRNA (например, SEQ ID No. 9), чтобы получить варианты или фрагменты ДНК-конструкции ddRNAi, которые по-прежнему функционировали бы для ингибирования или подавления экспрессии гена синтеза дитерпена, тем самым увеличивая содержание сахарозы в растении табака. Функциональные варианты и фрагменты ДНК-конструкции ddRNAi могут быть легко идентифицированы с применением стандартных лабораторных способов, чтобы определить, был ли уровень мРНК, кодируемой геном синтеза дитерпена, ниже уровня мРНК того же гена синтеза дитерпена в соответствующей растительной клетке дикого типа, выращенной в тех же условиях. Примером такой методики является полимеразная цепная реакция (ПЦР). Специалист в данной области поймет, что концентрацию полипептида, кодируемого геном синтеза дитерпена, можно измерить непосредственно в диких и трансгенных растениях с применением стандартных методов, таких как иммуноферментные анализы (ELISA), флуоресцентно-активированная сортировка клеток, вестерн-блоттинг или иммунопреципитация хроматина (ChIP).
В другом воплощении клетка табака, растение табака или его часть и/или материал для размножения растений могут содержать конструкцию, которая ингибирует активность или экспрессию гена синтеза дитерпена. В предпочтительном воплощении конструкция представляет собой ДНК-конструкцию ddRNAi, более предпочтительно ДНК-конструкция ddRNAi содержит модуль RNAi, который ингибирует активность или экспрессию гена циклазы 2 (CYC2), гена CBTol-циклазы или гена терпен-синтазы 3-8.
В дополнительном воплощении клетка табака, растение табака или его часть и/или материал для размножения растений могут содержать:
i) полинуклеотидную последовательность, показанную в данном документе как SEQ ID No. 5, SEQ ID No. 6 или SEQ ID No. 7, SEQ ID No. 11 или
ii) фрагмент полинуклеотидной последовательности, показанной в i), функциональный фрагмент которой ингибирует активность или экспрессию гена синтеза дитерпена, или
iii) полинуклеотидную последовательность, которая имеет, по меньшей мере, 70% (предпочтительно 85%, более предпочтительно 90%) идентичности с полинуклеотидом, показанным в i) или ii) выше.
В одном воплощении полинуклеотидная последовательность может иметь, по меньшей мере, 80% идентичности с SEQ ID No. 5, SEQ ID No. 6 или SEQ ID No. 7, или SEQ ID No. 11. Соответственно, полинуклеотидная последовательность может иметь, по меньшей мере, 90% идентичности с SEQ ID No. 5, SEQ ID No. 6 или SEQ ID No. 7, или SEQ ID No. 11. Подходящая полинуклеотидная последовательность может иметь, по меньшей мере, 95% идентичности (более подходящей идентичности, по меньшей мере, на 99%) с SEQ ID No. 5, SEQ ID No. 6 или SEQ ID No. 7, или SEQ ID No. 11.
В предпочтительном воплощении ингибирование активности или экспрессии гена синтеза дитерпена может привести к увеличению содержания сложного эфира сахарозы в растении табака и одновременно к изменению одного или нескольких других вкусовых компонентов растения табака. Ингибирование активности или экспрессии гена синтеза дитерпена может привести к увеличению содержания сложного эфира сахарозы и изменению содержания дитерпена. Увеличение содержания сложного эфира сахарозы может сопровождаться увеличением содержания одного или нескольких ароматических соединений, выбранных из группы, состоящей из: цис-абиенола, лабдендиола и CBT-ола.
Увеличение содержания сложного эфира сахарозы может сопровождаться увеличением содержания цис-абиенола и увеличением содержания лабдендиола. В другом воплощении увеличение содержания сложного эфира сахарозы может сопровождаться увеличением содержания лабдендиола. Увеличение содержания сложного эфира сахарозы может сопровождаться увеличением содержания CBT-ола и высоким содержанием цис-абиенола.
В одном воплощении способ по изобретению позволяет получить лист Берли или лист дымовой сушки, который имеет химию экссудата, как у турецкого табака. Предпочтительно, если лист Берли или лист дымовой сушки по химическому составу больше похож на табак, подобный турецкому, чем сравнительное растение Берли или растение дымовой сушки, в котором активность или экспрессия гена синтеза дитерпена не ингибирована.
В одном воплощении растение табака или его часть в соответствии с настоящим изобретением представляет собой растение Берли или растение дымовой сушки, модифицированное в соответствии с настоящим изобретением. В одном воплощении настоящее изобретение относится к растению Берли или растению дымовой сушки, модифицированной в соответствии с настоящим изобретением, чтобы иметь химический состав экссудата, подобный турецкому табаку. В одном воплощении растение табака (например, модифицированное растение табака) согласно настоящему изобретению не является растением табака восточного или турецкого типа.
В одном воплощении растение табака или его часть высушивают. В одном воплощении растение табака или его часть подвергают сушке, например, сушке воздухом, сушке дымом, сушке огнем или сушке на солнце. В дополнительном аспекте растение табака или его часть подвергают сушке дымом. В дополнительном аспекте растение табака или его часть подвергают сушке воздухом.
Сушка дымом хорошо известно в данной области и относится к процессу сушки табака с помощью дымоходов, которые питаются топками или системами, работающими на газе. Этот процесс нагревает табак, не подвергая его воздействию дыма, медленно повышая температуру в течение процесса сушки. Этот способ позволяет получить табак с высоким содержанием сахара и содержанием никотина от среднего до высокого. Сарай Смита для сушки табака (англ. Smith Tobacco Barn) является примером традиционного сарая для сушки табака дымом.
Табаки, высушенные воздухом, включают Берли, Мэриленд и темные табаки. Общим фактором является то, что сушка происходит в основном без искусственных источников тепла и влажности. Табаки Берли имеют цвет от светло- до темно-коричневого, с высоким содержанием масла и низким содержанием сахара. Табаки Берли высушены воздухом в сараях. Основными странами, выращивающими Берли, являются Аргентина, Бразилия, Италия, Малави и США. К растениям табака Берли относятся, например, Clay 402, Clay 403, Clay 502, Ky 14, Ky 907, Ky 910, Ky 8959, NC 2, NC 3, NC 4, NC 5, NC 2000, TN 86, TN 90, TN 97, R 610, R 630, R711, R 712, NCBH 129, Bu 21xKy 10, HB04P, Ky 14xL 8, Kt 200, Newton 98, Pedigo 561, Pf561 и Va 509.
Табаки Мэриленда чрезвычайно рыхлые, имеют хорошие свойства горения, низкий уровень никотина и нейтральный аромат. Основными странами, выращивающими Мэриленд, являются США и Италия. Темные табаки, полученные сушкой воздухом, отличаются от других типов, главным образом, процессом ферментации, который придает темному табаку воздушной сушки его цвет от среднего до темно-коричневого и отчетливый аромат. Их листья имеют низкое содержание сахара, но высокое содержание никотина. Темные табаки воздушной сушки в основном используются для производства жевательного табака и нюхательного табака. Основными регионами выращивания темных табаков огневой сушки являются Теннесси, Кентукки и Вирджиния, США.
В предпочтительном воплощении табак по изобретению содержит, по меньшей мере, столько же 3-метилвалерата, ацетилированного эфира сахарозы, сколько табаки, подобные турецким.
«Химический состав эксудата табака, подобного турецкому» означает, что экссудат, полученный из растения согласно настоящему изобретению, имеет химический состав эксудата, схожий с химическим составом эксудата, наблюдаемым в турецком табаке. Химический состав турецкого (или восточного) табака, например, N. tabacum var. Samsun охарактеризован R. Severson et al. 1985 (см. выше, который включен в данное описание ссылкой). Известно, что турецкий табак содержит низкие уровни CBT-диолов, высокие уровни продуктов окисления CBT и высокие уровни 3-метилвалериановой кислоты, содержащей сложный эфир сахарозы. Соответственно, настоящее изобретение преимущественно обеспечивает растения табака с большой биомассой, с эксудатом трихом табака, подобного турецкому.
В одном воплощении предложено растение табака, которое содержит сложные эфиры 3-метилвалерата и сахарозы. Растение табака может содержать сложные эфиры 3-метилвалерата и сахарозы по сравнению с растением табака, которое не было модифицировано в соответствии с настоящим изобретением, т.е. растением, в которое не было введено ингибирование гена синтеза дитерпена. Предпочтительно табачные линии по изобретению содержат, по меньшей мере, вдвое больше сложного эфира 3-метилвалерата и сахарозы, чем контрольное растение табака, которое не было модифицировано для ингибирования активности или экспрессии гена синтеза дитерпена. В дополнительном аспекте растение табака по изобретению содержит, по меньшей мере, столько же 3- метилвалерата, сколько табаки турецкого типа.
Преимущественно растения табака по изобретению имеют нормальный рост в полевых условиях. В одном воплощении растения, нокдауны CBTol-синтазы, преимущественно имеют низкое содержание CBT-диолов, сложные эфиры сахарозы с высоким содержанием цис-абиенола и высоким содержанием 3-метилвалериата, такие как турецкие табаки, но в отличие от турецких типов, параметры нокдауна CBTol-синтазы по изобретению могут давать более высокую биомассу, чем растения турецкого табака. Для турецких видов табака урожайность обычно находится в диапазоне от 600 до 1100 фунтов/акр.
Используемый в данном документе термин «функциональный фрагмент» относится к части полинуклеотида, которая способна функционировать так же, как и полинуклеотид. Например, если полинуклеотид является геном синтеза дитерпена, то функциональный фрагмент должен быть способен функционировать как ген синтеза дитерпена, например, функциональный фрагмент сохраняет активность гена синтеза дитерпена. Функциональный фрагмент может иметь уровень активности, который равен или превышает уровень активности полноразмерного полинуклеотида. Если полинуклеотид кодирует dsRNA или amiRNA, то функциональный фрагмент должен быть способен ингибировать активность или экспрессию гена синтеза дитерпена.
В одном воплощении функциональный фрагмент может быть частью гена синтеза дитерпена, как описано в данном документе, включающего, по меньшей мере, 50, 75, 100, 150, 200, 300, 400, 500, 600, 700, 800, 900 или 1000 смежных нуклеотидов. В некоторых воплощениях функциональный фрагмент может содержать, по меньшей мере, 150 нуклеотидов гена синтеза дитерпена, обсуждаемого в данном документе. В одном воплощении, где функциональный фрагмент представляет собой функциональный фрагмент ДНК-конструкции ddRNAi, функциональный фрагмент может быть частью RNAi, как обсуждается в настоящем документе, содержащей, по меньшей мере, 50, 75, 100, 150, 200 или 250 смежных нуклеотидов.
Используемый в данном документе термин «вырожденность генетического кода» относится к избыточности в кодонах, кодирующих полипептидные последовательности, которые проявляются как множественность комбинаций из трех кодонов, определяющих аминокислоту. Например, в молекуле мРНК, кодирующей полипептид, содержащий аминокислоту изолейцина, изолейцин может кодироваться AUU, AUC или AUA. Это означает, что молекула ДНК, кодирующая РНК, может иметь несколько последовательностей, но полученный полипептид будет иметь одну и ту же последовательность. Другими словами, полиморфные нуклеотидные последовательности могут кодировать один и тот же полипептидный продукт. Это означает, что одна последовательность нуклеиновой кислоты может содержать последовательность с очень низкой идентичностью последовательности со второй последовательностью при кодировании той же самой полипептидной последовательности.
Способ и применения согласно настоящему изобретению включают ингибирование, по меньшей мере, одного гена синтеза дитерпена. Ингибирование может быть достигнуто любым способом, известным специалисту в данной области техники.
В некоторых воплощениях настоящего изобретения может быть предусмотрен промотор. Промотор для применения в настоящем изобретении может быть одним или несколькими, выбранными из группы, состоящей из: конститутивного промотора, промотора, специфического для старения, тканеспецифичного промотора, промотора, регулируемого развитием и индуцибельного промотора. В одном воплощении промотор может быть конститутивным промотором.
Конститутивный промотор направляет экспрессию гена в различных частях растения непрерывно во время развития растения, хотя ген может не экспрессироваться на одном уровне во всех типах клеток. Примеры известных конститутивных промоторов включают промоторы, ассоциированные с транскриптом вируса мозаики цветной капусты 35S (Odell JT, Nagy F, Chua NH. (1985). Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter, Nature. 313 810-2, который включен в данное описание ссылкой) и гена актина 1 риса (Zhang W, McElroy D, Wu R. (1991) Analysis of rice Act1 5' region activity in transgenic rice plants. (Plant Cell 3 1155-65), который включен в данное описание ссылкой) и гена убиквитина 1 кукурузы (Cornejo MJ, Luth D, Blankenship KM, Anderson OD, Blechl AE. (1993). Activity of a maize ubiquitin promoter in transgenic rice. Plant Molec. Biol. 23 567-81 (который включен в данное описание ссылкой)). Конститутивные промоторы, такие как промотор вируса кольцевой гравировки гвоздики (CERV) (Hull R, Sadler J, LongstaffM (1986) The sequence of carnation etched ring virus DNA: comparison with cauliflower mosaic virus and retroviruses. EMBO Journal, 5 (2): 3083-3090), который включен в данное описание ссылкой).
Конститутивный промотор может быть выбран из: промотора вируса кольцевой гравировки гвоздики (CERV), вируса мозаики цветной капусты (промотор CaMV 35S), промотора гена актина 1 риса или гена убиквитина 1 кукурузы. Подходящим промотором может быть промотор CERV.
Альтернативно в некоторых воплощениях промотор может не представлять собой промотор вируса мозаики цветной капусты (промотор CaMV 35S). В одном воплощении промотор может представлять собой промотор, специфичный для старения. «Промотор, специфичный для старения» (SAG) может быть промотором, который связан с контролем экспрессии гена, ассоциированного со старением. Следовательно, промотор может ограничивать экспрессию кодирующей последовательности (то есть гена), с которой он функционально связан, по существу, исключительно в стареющей ткани. Следовательно, промотор, специфический для старения, может быть промотором, способным преимущественно стимулировать экспрессию генов в ткани растения регулируемым развитием образом, так что экспрессия 3'-области, кодирующей белок, происходит по существу только тогда, когда ткань растения подвергается старению. Понятно, что старение имеет тенденцию происходить в более старых частях растения, таких как более старые листья, а не в более молодых частях растения, таких как семена.
Одним из примеров растения, которое, как известно, экспрессирует многочисленные гены, связанные со старением, является Arabidopsis. Следовательно, промотор может быть выделен из гена, ассоциированного со старением у Arabidopsis. Gepstein et al. (The Plant Journal, 2003, 36, 629-642), который включен в данное описание ссылкой, провел детальное исследование SAG и их промоторов, используя Arabidopsis в качестве модели. Генетическая конструкция может содержать промотор из любых SAG, раскрытых в этой работе. Например, подходящий промотор может быть выбран из группы, состоящей из SAG12, SAG13, SAG101, SAG21 и SAG18, или их функционального варианта или функционального фрагмента.
В одном воплощении промотор может представлять собой промотор SAG12 или SAG13. В одном воплощении промотор может представлять собой промотор SAG12, который будет известен квалифицированному специалисту, или его функциональный вариант или его фрагмент (Gan & Amasino, 1997, Plant Physiology, 113: 313-319), который включен в данное описание ссылкой. Подходящие промоторы и их последовательности можно найти в WO 2010/097623 (включенной в настоящий документ ссылкой).
В другом воплощении промотор может представлять собой тканеспецифичный промотор. Тканеспецифичным промотором является тот, который направляет экспрессию гена в одной (или нескольких) частях растения, обычно на протяжении всей жизни этих частей растения. Категория тканеспецифичных промоторов обычно также включает промоторы, специфичность которых не является абсолютной, то есть они могут также направлять экспрессию на более низком уровне в тканях, отличных от предпочтительной ткани. Ряд тканеспецифичных промоторов известен в данной области и включает те, которые связаны с геном пататина, экспрессируемым в клубнях картофеля, и высокомолекулярным геном глютенина, экспрессируемым в эндосперме пшеницы, ячменя или кукурузы. Любой из этих промоторов может быть использован в настоящем изобретении.
Подходящим тканеспецифичным промотором может быть листоспецифичный промотор. Подходящие листоспецифичные промоторы могут включать ASYMMETRIC LEAVES 1 (AS1). В особенно предпочтительном воплощении тканеспецифичный промотор не является корнеспецифичным промотором.
В другом воплощении промотор может представлять собой промотор, регулируемый развитием. Регулятор, регулируемый развитием, направляет изменение экспрессии гена в одной или нескольких частях растения в определенное время в процессе развития растения. Ген может экспрессироваться в этой части растения в другое время на другом (обычно более низком) уровне, а также может экспрессироваться в других частях растения.
В одном воплощении промотор может быть индуцибельным промотором. Индуцибельный промотор способен направлять экспрессию гена в ответ на индуктор. В отсутствие индуктора ген не будет экспрессироваться. Индуктор может действовать непосредственно на промоторную последовательность или может действовать путем противодействия действию молекулы-репрессора. Индуктор может представлять собой химический агент, такой как метаболит, белок, регулятор роста или токсический элемент, физиологический стресс, такой как тепло, ранение или осмотическое давление, или косвенное последствие действия патогена или вредителя. Регулятор, регулируемый развитием, может быть описан как специфический тип индуцибельного промотора, реагирующий на эндогенный индуктор, продуцируемый растением, или на стимул окружающей среды в определенной точке жизненного цикла растения. Примеры известных индуцибельных промоторов включают промоторы, связанные с реакцией на рану, такие как описанные Warner SA, Scott R, Draper J. (1993) (Isolation of an asparagus intracellular PR gene (AoPR1) wound-responsive promoter by the inverse polymerase chain reaction and its characterization in transgenic tobacco. Plant J. 3 191-201.) который включен в данное описание ссылкой, температурный отклик, как раскрыто Benfey & Chua (1989) (Benfey, P.N., and Chua, N-H. (1989) Regulated genes in transgenic plants. Science 244 174-181), который включен в данное описание ссылкой, и химически индуцированные, как описано Gatz (1995) (Gatz, C. (1995) Novel inducible/repressible gene expression systems. Методы в клеточном биол. 50 411-424), который включен в данное описание ссылкой.
Таким образом, в одном воплощении промотор может быть выбран из группы, состоящей из: промотора CERV, промотора 35S вируса мозаики цветной капусты (полного или усеченного), промотора rubisco, промотора горохового пластоцианина, промотора нопалинсинтазы, промотор белка, связывающего хлорофилл r/b, промотора высокомолекулярного глютенина, промотора α, β-глиадина, промотора гордеина и промотора пататина.
В одном воплощении промотор может быть промотором 35S CaMV или модифицированным промотором 35S с дублированной энхансерной областью или двойной энхансерной областью (R. Kay et al. Science. 1987 Jun 5; 236 (4806): 1299-302, который включен в данное описание ссылкой).
Рекомбинантный вектор также может содержать ДНК, кодирующую ген, который можно использовать в качестве селектируемого маркера в процессе клонирования, то есть, чтобы сделать возможным отбор клеток, которые были трансфицированы или трансформированы, и сделать возможным отбор клеток, содержащих векторы, включающие гетерологичную ДНК. Вектор также может содержать ДНК, участвующую в регуляции экспрессии кодирующей последовательности или для нацеливания экспрессированного полипептида на определенную часть клетки-хозяина, например, хлоропласт. Следовательно, вектор может содержать, по меньшей мере, один дополнительный элемент, выбранный из группы, состоящей из: селектируемого маркерного гена (например, гена устойчивости к антибиотику); сигнала терминации полипептида; и белковой таргетной последовательности (например, транзитный пептид хлоропласта).
Примеры подходящих маркерных генов включают гены устойчивости к антибиотикам, такие как гены, придающие устойчивость к канамицину, генетицину (G418) и гигромицину (npt-II, hyg-B); гены устойчивости к гербицидам, такие как гены, придающие устойчивость к гербицидам на основе фосфинотрицина и сульфонамида (соответственно bar и suI; EP-A-242246, EP-A-0249637); и маркеры для скрининга, такие как бета-глюкуронидаза (GB2197653), люцифераза и зеленый флуоресцентный белок (GFP). Маркерный ген может контролироваться вторым промотором, который обеспечивает экспрессию в клетках, которые могут присутствовать или не присутствовать в семени, что позволяет отбирать клетки или ткани, содержащие маркер, на любой стадии развития растения. Подходящими вторыми промоторами являются промотор гена нопалинсинтазы Agrobacterium и промотор, полученный из гена, который кодирует транскрипт вируса мозаики цветной капусты 35S (CaMV). Однако может быть использован любой другой подходящий второй промотор.
Коммерчески желательные признаки
Термин «коммерчески желательные признаки» будет включать такие признаки, как урожайность, высота зрелого растения, количество собираемых листьев, средняя длина узла, длина листа режущей кромки, ширина листа режущей кромки, качество, устойчивость к абиотическим стрессам (например, к засухе), устойчивость к гербицидам и/или биотическая стрессоустойчивость (например, к насекомым, бактериям или грибкам).
Термин «коммерчески желательные признаки», при описании в данном документе, означает одну или несколько характеристик, выбранных из группы, состоящей из устойчивости к засухе, устойчивости к вредителям, высоты зрелого растения, количества собираемых листьев, средней длины узла, длины листа режущей части, ширины листа режущей части и сопоставимой урожайности по сравнению с этими указанными признаками у высушенного дымом родителя сопоставимого растения при выращивании в аналогичных полевых условиях.
Если не указано иное, используемая в данном документе урожайность табака относится к урожайности высушенных листьев, которая рассчитывается на основе массы высушенных листьев табака на акр в стандартных полевых условиях в соответствии со стандартной агрономической практикой и практикой отверждения.
В одном аспекте растение табака согласно настоящему изобретению имеет выход от 50% до 150%, от 55% до 145%, от 60% до 140%, от 65% до 135%, от 70% до 130%, от 75% до 125%, от 80% до 120%, от 85% до 115%, от 90% до 110%, от 95% до 105%, от 50% до 100%, от 55% до 100%, от 60% и 100%, от 65% до 100%, от 70% до 100%, от 75% до 100%, от 80% до 100%, от 85% до 100%, от 90% до 100%, от 95% до 100%, от 100% до 150%, от 105% до 150%, от 110% до 150%, от 115% до 150%, от 120% до 150%, от 125% до 150%, от 130% до 150 %, от 135% до 150%, от 140% до 150% или от 145% до 150% урожая сопоставимого растения при выращивании в аналогичных полевых условиях.
В другом аспекте урожайность табачных растений согласно настоящему изобретению составляет около 1,5, 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 2,6, 2,7, 2,8, 2,9 или 3,0 раза от урожайности сопоставимого растения при выращивании в аналогичных полевых условиях
В другом аспекте урожайность растения табака согласно настоящему изобретению сопоставима с урожайностью растения дымовой сушки, при выращивании в аналогичных полевых условиях.
В одном аспекте растение табака согласно настоящему изобретению обеспечивает урожайность, выбранную из группы, состоящей из около от 1200 до 3500, от 1300 до 3400, от 1400 до 3300, от 1500 до 3200, от 1600 до 3100, от 1700 до 3000 от 1800 до 2900, от 1900 до 2800, от 2000 до 2700, от 2100 до 2600, от 2200 до 2500 и от 2300 до 2400 фунтами/акр.
В другом аспекте растение табака согласно настоящему изобретению обеспечивает урожайность, выбранную из группы, состоящей из около от 1200 до 3500, от 1300 до 3500, от 1400 до 3500, от 1500 до 3500, от 1600 до 3500, от 1700 до 3500 от 1800 до 3500, от 1900 до 3500, от 2000 до 3500, от 2100 до 3500, от 2200 до 3500, от 2300 до 3500, от 2400 до 3500, от 2500 до 3500, от 2600 до 3500, от 2700 до 3500 от 2800 до 3500, от 2900 до 3500, от 3000 до 3500 и от 3100 до 3500 фунтами/акр.
В дополнительном аспекте растение табака согласно настоящему изобретению обеспечивает урожайность, выбранную из группы, состоящей из около от 1200 до 3500, от 1200 до 3400, от 1200 до 3300, от 1200 до 3200, от 1200 до 3100, от 1200 до 3000, от 1200 до 2900, от 1200 до 2800, от 1200 до 2700, от 1200 до 2600, от 1200 до 2500, от 1200 до 2400, от 1200 до 2300, от 1200 до 2200, от 1200 до 2100, от 1200 до 2000, от 1200 до 1900, от 1200 до 1800, от 1200 до 1700, от 1200 до 1600, от 1200 до 1500 и от 1200 до 1400 фунтами/акр.
Растения табака
Настоящее изобретение относится к способам, применениям, направленным на растения табака, а также к клетке табака, растению табака и материалу для размножения растений.
Используемый в данном документе термин «табак» относится к растению рода Nicotiana, которое используется для производства табачных изделий. Неограничивающие примеры подходящих растений «табака» включают N. tabacum и N. rustica (например, N. tabacum L., LA B21, LN KY171, Tl 1406, Basma, Galpao, Perique, Beinhart 1000-1 и Petico).
В одном воплощении подходящим растением табака может быть любой вид N. tabacum.
В другом воплощении подходящее растение табака может представлять собой разновидность, не относящуюся к табаку.
Табачный материал может быть получен или его получают из разновидностей Nicotiana tabacum, широко известных как разновидности Берли, дымовые или светлые и темные. В некоторых воплощениях табачный материал получают из растений Берли, Вирджинии или темного табака. Растение табака может быть выбрано из табака Берли, редкого табака, специального табака, взорванного табака или т.п.
Также в настоящем документе предлагается применение сортов табака и элитных сортов табака. Следовательно, растение табака для применения в данном документе может представлять собой сорт табака или элитный сорт табака. Особенно полезные сорта Nicotiana tabacum включают табаки типа Берли, темного типа и дымового типа.
В некоторых воплощениях растение табака может быть выбрано, например, из одного или нескольких следующих сортов: N. tabacum L. cultivar TI 1068, N. tabacum AA 37-1, N. tabacum B 13P, N.tabacum Xanthi. (Митчелл-Мор), N.tabacum KT D # 3 Hybrid 107, N. tabacum Bel-W3, N.tabacum 79-615, N.tabacum Samsun Holmes NN, F4 из кросса N.tabacum BU21 x N.tabacum Hoja Parado, линия 97, N.tabacum KTRDC # 2 Hybrid 49, N.tabacum KTRDC # 4 Hybrid 1 10, N.tabacum Burley 21, N.tabacum PM016, N.tabacum KTRDC # 5 KY 160 SI, N.tabacum KTRDC # 7 FCA, N.tabacum KTRDC # 6 TN 86 SI, N.tabacum PM021, N.tabacum K 149, N.tabacum K 326, N.tabacum K 346, N.tabacum K 358, N.tabacum K 394, N.tabacum K 399, N.tabacum K 730, N.tabacum KY 10, N.tabacum KY 14, N.tabacum KY 160, N.tabacum KY 17, N.tabacum KY 8959, N.tabacum KY 9, N.tabacum KY 907, N.tabacum MD 609, N.tabacum McNair 373, N.tabacum NC 2000, N.tabacum PG 01, N.tabacum PG 04, N.tabacum P01, N.tabacum P02, N.tabacum P03, N.tabacum RG 11, N.tabacum RG 17, N.tabacum R G 8, N.tabacum Speight G-28, N.tabacum TN 86, N.tabacum TN 90, N.tabacum VA 509, N.tabacum AS44, N.tabacum Banket A1, N.tabacum Basma Drama B84/31, N.tabacum Basma I Zichna ZP4/B, N.tabacum Basma Xanthi BX 2A, N.tabacum Batek, N.tabacum Besuki Jember, N.tabacum C104, N.tabacum Coker 319, N.tabacum Coker 347, N.tabacum Criollo Misionero, N.tabacum PM092, N.tabacum Delcrest, N.tabacum Djebel 81, N.tabacum DVH 405, N.tabacum GalpaoComum, N.tabacum HB04P, N.tabacum Hicks Broadleaf, N.tabacum Kabakulak Elassona, N.tabacum PM102, N.tabacum Kutsage E1, N.tabacum KY 14xL8, N.tabacum KY 171, N.tabacum LA BU 21, N.tabacum McNair 944, N.tabacum NC 2326, N.tabacum NC 71, N.tabacum NC 297, N.tabacum NC 3, N.tabacum PVH 03, N.tabacum PVH 09, N.tabacum PVH 19, N.tabacum PVH 21 10, N.tabacum Red Russian, N.tabacum Samsun, N.tabacum Saplak, N.tabacum Simmaba, N.tabacum Talgar 28, N.tabacum PM132, N.tabacum Wislica, N.tabacum Yayaldag, N.tabacum NC 4, N.tabacum TR Madole, N.tabacum Prilep HC-72, N.tabacum Prilep P23, N.tabacum Prilep PB 156/1, N.tabacum Prilep P12-2/1, N.tabacum Yaka JK-48, N.tabacum Yaka JB 125/3, N.tabacum ΤΊ-1068, N.tabacum KDH-960, N.tabacum TI-1070, N.tabacum TW136, N.tabacum PM204, N.tabacum PM205, N.tabacum Basma, N.tabacum TKF 4028, N.tabacum L8, N.tabacum TKF 2002, N.tabacum TN90, N.tabacum GR141, N.tabacum Basma xanthi, N.tabacum GR149, N.tabacum GR153 и N. tabacum Petit Havana.
Неограничивающими примерами разновидностей или сортов являются: BD 64, CC 101 , CC 200, CC 27, CC 301 , CC 400, CC 500, CC 600, CC 700, CC 800, CC 900, Coker 176, Coker 319, Coker 371 Gold, Coker 48, CD 263, DF91 1 , DT 538 LC Galpao tobacco, GL 26H, GL 350, GL 600, GL 737, GL 939, GL 973, HB 04P, HB 04P LC, HB3307PLC, Hybrid 403LC, Hybrid 404LC, Hybrid 501 LC, K 149, K 326, K 346, K 358, K394, K 399, K 730, KDH 959, KT 200, KT204LC, KY10, KY14, KY 160, KY 17, KY 171 , KY 907, KY907LC, KTY14xL8 LC, Little Crittenden, McNair 373, McNair 944, msKY 14xL8, Narrow Leaf Madole, Narrow Leaf Madole LC, NBH 98, N-126, N-777LC, N-7371 LC, NC 100, NC 102, NC 2000, NC 291 , NC 297, NC 299, NC 3, NC 4, NC 5, NC 6, NC7, NC 606, NC 71 , NC 72, NC 810, NC BH 129, NC 2002, Neal Smith Madole, OXFORD 207, PD 7302 LC, PD 7309 LC, PD 7312 LC 'Periq'e' tobacco, PVH03, PVH09, PVH19, PVH50, PVH51 , R 610, R 630, R 7-1 1 , R 7-12, RG 17, RG 81 , RG H51 , RGH 4, RGH 51 , RS 1410, Speight 168, Speight 172, Speight 179, Speight 210, Speight 220, Speight 225, Speight 227, Speight 234, Speight G-28, Speight G-70, Speight H-6, Speight H20, Speight NF3, Tl 1406, Tl 1269, TN 86, TN86LC, TN 90, TN 97, TN97LC, TN D94, TN D950, TR (Tom Rosson) Madole, VA 309, VA359, AA 37-1 , B 13P, Xanthi (Mitchell-Mor), Bel-W3, 79-615, Samsun Holmes NN, KTRDC number 2 Hybrid 49, Burley 21 , KY 8959, KY 9, MD 609, PG 01 , PG 04, P01 , P02, P03, RG 11 , RG 8, VA 509, AS44, Banket A1 , Basma Drama B84/31 , Basma I Zichna ZP4/B, Basma Xanthi BX 2A, Batek, Besuki Jember, C104, Coker 347, Criollo Misionero, Delcrest, Djebel 81 , DVH 405, GalpaoComum, HB04P, Hicks Broadleaf, Kabakulak Elassona, Kutsage E1 , LA BU 21, NC 2326, NC 297, PVH 21 10, Red Russian, Samsun, Saplak, Simmaba, Talgar 28, Wislica, Yayaldag, Prilep HC-72, Prilep P23, Prilep PB 156/1 , Prilep P12-2/1 , Yaka JK-48, Yaka JB 125/3, TI-1068, KDH-960, Tl-1070, TW136, Basma, TKF 4028, L8, TKF 2002, GR141 , Basma xanthi, GR149, GR153, Petit Havana. Также предусмотрены подразновидности вышеописанных, даже если они конкретно не определены в настоящем документе.
Табак может быть табаком Берли.
В одном воплощении растение табака представляет собой N. tabacum L. В наиболее предпочтительном воплощении растение табака представляет собой N. tabacum L культурный сорт TI 1068.
В одном воплощении материал для размножения растений может быть получен из растения табака по изобретению. Используемый в данном документе термин «материал для размножения растений» относится к любому растительному веществу, взятому из растения, из которого могут быть получены другие растения. Подходящим материалом для размножения растений может быть семя.
В одном воплощении клетки табака, растение табака и/или материал для размножения растений по изобретению может содержать ингибированный или подавленный ген синтеза дитерпена. В другом воплощении клетки табака, растение табака и/или материал для размножения растений могут содержать конструкцию или вектор согласно изобретению. В другом воплощении клетки табака, растение табака и/или материал для размножения растений могут быть получены (например, получен) способом согласно изобретению.
Соответственно, растение табака или его часть в соответствии с настоящим изобретением может иметь пониженную экспрессию, по меньшей мере, одного гена синтеза дитерпена по сравнению с растением табака или его частью, которое не было модифицировано для ингибирования экспрессии гена синтеза дитерпена.
В одном воплощении растение табака или его часть в соответствии с настоящим изобретением содержит клетку табака по изобретению. В другом воплощении материал для размножения растений может быть получен (например, получен) из растения табака по изобретению.
В одном воплощении предлагается применение клеток табака, как предусмотрено в предыдущих воплощениях, для производства табачного изделия. Кроме того, предлагается применение растения табака, как описано в данном документе, для селекции растений табака.
Настоящее изобретение также относится в другом воплощении к применению растения табака вышеупомянутых воплощений для производства табачного продукта. В другом воплощении предлагается применение растения табака по изобретению для выращивания культуры. В одном воплощении применение гена синтеза дитерпена в соответствии с настоящим изобретением может привести к изменению химического состава экссудата растения табака.
В одном воплощении применение гена синтеза дитерпена согласно настоящему изобретению может привести к увеличению содержания сложного эфира сахарозы в растении табака. В другом воплощении применение гена синтеза дитерпена (например, его ингибирование) может привести к увеличению концентрации эфира сахарозы в экссудате трихом и к увеличению содержания одного или нескольких из числа CBT-ола, цис-абиенола и лабдендиола. Соответственно, это может наблюдаться, когда растение проявляет пониженную экспрессию гена синтеза дитерпена по сравнению с растениями дикого типа.
В одном воплощении настоящее изобретение относится к культуре клеток табака (например, в культуре in vitro). Культура клеток табака может представлять собой суспензионную культуру клеток табака. Эти клетки, культивируемые in vitro, могут быть включены в табачный продукт, например, в качестве заменителя обычных табачных частиц, кусочков, тонко нарезанного или длинного среза табачной пластинки, в качестве добавочного ингредиента или в качестве заменителя и добавки.
В одном воплощении предлагается применение культуры клеток табака, например, культуры собранных и/или обработанных клеток табака, или их экстракта согласно настоящему изобретению для производства табачного продукта.
Клетки табака, собранные из культуры in vitro, могут быть высушены, например, лиофилизированы, например, для получения порошка.
Специалист в данной области осведомлен об известных способах создания культур клеток табака in vitro. В качестве примера можно использовать только следующий метод: сбор семян из интересующего растения табака и стерилизацию их внешней поверхности для уничтожения нежелательных организмов, посадку указанных семян для выращивания интересующего растения табака, удаление ткани из растения табака (например, из стебля табака) для использования в качестве эксплантата, создания каллюсной культуры из эксплантата табака, создания культуры клеточной суспензии из каллюсной культуры и сбора культурального материала (например, включая клетки табака) для получения культуры клеток табака.
Клетки табака можно собирать различными способами, включая фильтрацию, например, вакуумную фильтрацию. Образец может быть промыт в фильтре путем добавления воды, а оставшаяся жидкость удалена фильтрацией, например, вакуумной фильтрацией.
Собранную культуру клеток табака можно дополнительно обработать, например, высушить, например, высушить на воздухе и/или высушить лиофилизацией. Собранная культура клеток табака или высушенная культура клеток табака или экстракт из нее могут быть включены в табачные изделия согласно настоящему изобретению.
Продукты
Настоящее изобретение также относится к продуктам, которые можно получить или которые получены из табака в соответствии с настоящим изобретением. Предлагаются продукты, которые можно получить или которые получают из растения табака, в котором активность или экспрессия гена синтеза дитерпена была ингибирована и которое содержит повышенное содержание сложного эфира сахарозы.
Используемый в данном документе термин «продукт табачной промышленности» предназначен для включения горючих курительных изделий, таких как сигареты, сигариллы, сигары, табак для трубок или для сигарет без упаковки (на основе табака, производных табака, взорванного табака, восстановленного табака, заменителей табака или другого курительного материала), системы подачи негорючего аэрозоля, такие как продукты нагрева, которые выделяют соединения из материалов подложки без сжигания, такие как электронные сигареты, продукты нагревания табака и гибридные системы для генерирования аэрозоля из комбинации материалов подложки, например, гибридные системы, содержащие жидкую или гелевую или твердую подложку, а также аэрозолизируемые материалы подложки, используемые в этих системах подачи аэрозоля; и изделия для доставки без аэрозолей, такие как пастилки, жевательные резинки, пластыри, изделия, содержащие воздухопроницаемые порошки, и бездымные табачные изделия, такие как снюс и нюхательный табак, из которых изделия для доставки без аэрозолей могут доставлять или не доставлять никотин.
Соответственно, продукт табачной промышленности может быть приготовлен из (например, может содержать) растения табака или его части в соответствии с настоящим изобретением.
Соответственно, продукт табачной промышленности может быть получен из культуры клеток табака в соответствии с настоящим изобретением.
Соответственно, продукт табачной промышленности может быть приготовлен из (например, может содержать) растения табака или его части, размноженного из материала для размножения табака в соответствии с настоящим изобретением.
Соответственно, продукт табачной промышленности может быть приготовлен из (например, может содержать) собранного листа растения табака согласно настоящему изобретению.
Соответственно, продукт табачной промышленности может быть приготовлен из (например, может содержать) обработанного табачного листа в соответствии с настоящим изобретением.
Соответственно, продукт табачной промышленности может быть приготовлен из (например, может содержать) высушенного табачного материала согласно настоящему изобретению.
Соответственно, продукт табачной промышленности может быть приготовлен из (например, может содержать) табачной смеси согласно настоящему изобретению.
В одном воплощении продукт табачной промышленности представляет собой горючее курительное изделие, выбранное из группы, состоящей из сигареты, сигариллы и сигары.
В одном воплощении продукт табачной промышленности содержит один или более компонентов горючего курительного изделия, такого как фильтр, фильтрующий стержень, сегменты фильтрующего стержня, табак, табачный стержень, сегмент табачного стержня, жгут, высвобождающийся компонент добавки, такой как капсула, нить, шарики, бумага, такая как фицелла, ободковая бумага или сигаретная бумага.
В одном воплощении продукт табачной промышленности представляет собой систему подачи негорючего аэрозоля.
В одном воплощении продукт табачной промышленности содержит один или несколько компонентов негорючей системы подачи аэрозоля, такой как нагреватель и аэрозолизируемый субстрат.
В одном воплощении система подачи аэрозоля представляет собой электронную сигарету, также известную как устройство для вейпинга.
В одном воплощении электронная сигарета содержит нагреватель, источник питания, способный подавать питание на нагреватель, аэрозолизируемый субстрат, такой как жидкость или гель, корпус и, при необходимости, мундштук.
В одном воплощении аэрозолизируемый субстрат содержится в контейнере для субстрата. В одном воплощении контейнер с субстратом объединен с нагревателем или содержит его.
В одном воплощении продукт табачной промышленности представляет собой продукт для нагревания, который высвобождает одно или несколько соединений при нагревании, но не при сжигании, материала субстрата. Материал субстрата представляет собой аэрозолизируемый материал, который может представлять собой, например, табак или другие нетабачные продукты, которые могут содержать или не содержать никотин. В одном воплощении продукт нагревания представляет собой продукт нагревания табака.
В одном воплощении продукт нагревания представляет собой электронное устройство.
В одном воплощении продукт для нагревания табака содержит нагреватель, источник питания, способный подавать питание на нагреватель, аэрозолизируемый субстрат, такой как твердый или гелеобразный материал.
В одном воплощении нагревательный продукт представляет собой неэлектронное изделие.
В одном воплощении нагревательный продукт содержит аэрозолизируемый субстрат, такой как твердый или гелеобразный материал, и источник тепла, который способен подавать тепловую энергию на аэрозолизируемый субстрат без каких-либо электронных средств, как например, путем сжигания горючего материала, такого как уголь.
В одном воплощении нагревательный продукт также содержит фильтр, способный фильтровать аэрозоль, образующийся при нагревании аэрозолизируемого субстрата.
В некоторых воплощениях изобретения материал субстрата, способный к аэрозолизации, может содержать парообразующий или аэрозольгенерирующий агент или увлажнитель, такой как глицерин, пропиленгликоль, триацетин или диэтиленгликоль.
В одном воплощении продукт табачной промышленности представляет собой гибридную систему для генерирования аэрозоля путем нагревания, но не сжигания, комбинации материалов субстрата. Материалы субстрата могут содержать, например, твердое вещество, жидкость или гель, которые могут содержать или не содержать никотин. В одном воплощении гибридная система содержит жидкий или гелевый субстрат и твердый субстрат. Твердым субстратом может быть, например, табак или другие нетабачные продукты, которые могут содержать или не содержать никотин. В одном воплощении гибридная система содержит жидкий или гелевый субстрат и табак.
В другом воплощении продукт может содержать ДНК-конструкцию ddRNAi по изобретению, которая ингибирует активность или экспрессию гена синтеза дитерпена и повышенное содержание сложного эфира сахарозы.
В одном воплощении предлагается применение растения табака по изобретению для получения листа табака. Соответственно, лист табака может быть подвергнут последующим применениям, таким как обработка. Таким образом, в одном воплощении применение вышеизложенного воплощения может обеспечить обработанный табачный лист. Соответственно, табачный лист можно подвергать сушке, ферментации, пастеризации или их комбинациям.
В другом воплощении табачный лист может быть разрезан. В некоторых воплощениях табачный лист может быть разрезан до или после его сушки, ферментации, пастеризации или их комбинаций.
В одном воплощении в настоящем изобретении предлагается собранный лист растения табака по изобретению. В одном воплощении собранный лист можно получить из растения табака, которое ингибировало активность или экспрессию гена синтеза дитерпена и повышенное содержание сложного эфира сахарозы. В дополнительном воплощении собранный лист может быть получен (например, получен) из растения табака, размноженного из материала для размножения согласно настоящему изобретению. В другом воплощении предлагается собранный лист, получаемый способом или применением согласно настоящему изобретению. Соответственно, собранный лист может быть срезанным собранным листом. В некоторых воплощениях собранный лист может содержать жизнеспособные клетки табака. В некоторых воплощениях собранный лист не содержит жизнеспособных клеток табака. В других воплощениях собранный лист может быть подвергнут дальнейшей обработке.
Некоторые растения табака можно собирать, обрезая стебли и собирая все листья одновременно (например, как с табаком Берли). Другие растения табака (например, табак дымовой сушки) можно собирать постадийно в процессе, таком как праймирование, при котором отдельные листья удаляют со стебля по мере их созревания.
Также предлагается обработанный табачный лист. Обработанный табачный лист может быть получен из растения табака по изобретению. Соответственно, обработанный табачный лист может быть получен из растения табака, полученного в соответствии с любым из способов и/или применений настоящего изобретения. В одном воплощении обработанный лист табака может быть получен из растения табака, которое ингибировало активность или экспрессию гена синтеза дитерпена и повышенное содержание сложного эфира сахарозы, предпочтительно по сравнению с контрольным листом, т.е. по сравнению с листом из растения табака, которое не было модифицировано по изобретению. Обработанный табачный лист может иметь снижение активности или экспрессии гена синтеза дитерпена и повышенное содержание сложного эфира сахарозы.
В другом воплощении обработанный лист табака может быть получен из растения табака, размноженного из материала для размножения растения табака согласно настоящему изобретению. Обработанный табачный лист согласно настоящему изобретению получают обработкой собранного листа согласно настоящему изобретению.
Используемый в данном документе термин «обработанный табачный лист» относится к табачному листу, который прошел одну или несколько стадий обработки, которым подвергается табак в данной области техники. «Обработанный табачный лист» не содержит или практически не содержит жизнеспособных клеток.
Термин «жизнеспособные клетки» относится к клеткам, которые способны расти и/или являются метаболически активными. Таким образом, если клетка считается нежизнеспособной, также называемой нежизнеспособной, то клетка не демонстрирует характеристики жизнеспособной клетки.
Термин «по существу не жизнеспособные клетки» означает, что жизнеспособны менее чем около 5% всех клеток. Предпочтительно менее чем около 3%, более предпочтительно менее чем около 1%, еще более предпочтительно менее чем около 0,1% всех клеток являются жизнеспособными.
В одном воплощении обработанный табачный лист может быть обработан одним или несколькими способами: сушки, ферментации и/или пастеризации. Соответственно обработанный табачный лист может быть обработан сушкой. Табачный лист может быть высушен любым способом, известным в данной области. В одном воплощении табачный лист может быть высушен одним или несколькими способами сушки, выбранными из группы, состоящей из: воздушной сушки, огневой сушки, дымовой сушки и сушки на солнце. Соответственно, табачный лист может быть высушен на воздухе. Соответственно, табачный лист может быть высушен дымовой сушкой.
В одном воплощении обработанный лист согласно настоящему изобретению имеет повышенное содержание сложного эфира сахарозы по сравнению с сопоставимым продуктом (например, обработанным листом), который не был модифицирован в соответствии с настоящим изобретением.
В одном аспекте лист, высушенный на воздухе, в соответствии с настоящим изобретением имеет повышенное содержание сложного эфира сахарозы по сравнению со сравнимым продуктом (например, листом, отверждаемым воздухом), который не был модифицирован в соответствии с настоящим изобретением.
Соответственно, содержание сложного эфира сахарозы может быть измерено в нижних 1/3 листьев растений, высушенных на воздухе. Соответственно, содержание сложного эфира сахарозы может быть измерено в средних 1/3 листьев растений, срезанных со стеблем и высушенных на воздухе. Соответственно, содержание сложного эфира сахарозы может быть измерено в верхних 1/3 листьев растений, срезанных со стеблем и высушенных на воздухе.
Используемый в данном документе термин «нижние листья» относятся к листьям в нижней трети растения (например, листья, расположенные ближе всего к основанию растения), «верхние листья» в данном описании относятся к листьям в верхней трети растения (например, листья дальше всего от основания растения). «Средние листья» в контексте настоящего описания относятся к центральной трети растения между нижним и верхним положениями (например, листья на полпути между нижним и верхним листьями).
Соответственно, нижняя 1/3 листьев растений, срезанных со стеблем и высушенных на воздухе, согласно настоящему изобретению может иметь повышенное содержание сложного эфира сахарозы по сравнению с сопоставимым продуктом (например, нижняя 1/3 листьев растений, срезанных со стеблем и высушенных на воздухе), который не был модифицирован в соответствии с настоящим изобретением. Соответственно, средние 1/3 листьев растений, срезанных со стеблем и высушенных на воздухе, согласно настоящему изобретению могут иметь повышенное содержание сложного эфира сахарозы по сравнению с сопоставимым продуктом (например, средние 1/3 листьев растений, срезанных со стеблем и высушенных на воздухе), который не был модифицирован в соответствии с настоящим изобретением. Соответственно, верхняя 1/3 листьев растений, срезанных со стеблем и высушенных на воздухе, согласно настоящему изобретению может иметь повышенное содержание сложного эфира сахарозы по сравнению с сопоставимым продуктом (например, верхняя 1/3 листьев растений, срезанных со стеблем и высушенных на воздухе), который не был модифицирован в соответствии с настоящим изобретением.
В одном аспекте лист, высушенный дымовой сушкой, в соответствии с настоящим изобретением имеет повышенное содержание сложного эфира сахарозы по сравнению с сопоставимым продуктом (например, лист, высушенный дымовой сушкой), который не был модифицирован в соответствии с настоящим изобретением.
Соответственно, содержание сложного эфира сахарозы может быть измерено при первой ломке растений, высушиваемых дымовой сушкой. Соответственно, содержание сложного эфира сахарозы может быть измерено во второй (или, альтернативно, в средней) ломке растений, высушиваемых дымовой сушкой. Соответственно, содержание сложного эфира сахарозы может быть измерено при третьей (или, альтернативно, последней) ломке растений, высушенных дымовой сушкой.
Используемый в данном документе термин «ломка» относится к удалению листьев с растений табака. Это может относиться к удалению зрелых или спелых листьев растений, высушиваемых дымовой сушкой.
«Первая ломка» в контексте настоящего описания относится к листьям, которые сначала собирают с растения табака (например, листья с самой низкой части растения табака). «Вторая ломка (или средняя ломка)», при использовании в данном описании, относится к листьям, которые собирают с растения табака после первоначальной ломки (например, листья из средней части растения табака). «Третья (или последняя) ломка», при использовании в данном описании, относится к листьям, которые собирают последними с растения табака (например, листья из верхней части растения табака).
Соответственно, первая ломка листьев растений, высушиваемых дымовой сушкой, в соответствии с настоящим изобретением может содержать повышенное содержание сложного эфира сахарозы по сравнению с сопоставимым продуктом (например, первая ломка листьев растений, высушиваемых дымовой сушкой), который не был модифицирован в соответствии с настоящим изобретением. Соответственно, вторая (или средняя) ломка листьев растений, высушиваемых дымовой сушкой, в соответствии с настоящим изобретением может содержать повышенное содержание сложного эфира сахарозы по сравнению с сопоставимым продуктом (например, вторая (или средняя) ломка листьев растений, подвергнутых дымовой сушке), который не был модифицирован в соответствии с настоящим изобретением. Соответственно, третья (или заключительная) ломка листьев растений, высушиваемых дымовой сушкой, согласно настоящему изобретению может иметь повышенное содержание сложного эфира сахарозы по сравнению с сопоставимым продуктом (например, третья (или последняя) ломка листьев растений, высушиваемых дымовой сушкой), которые не были модифицированы в соответствии с настоящим изобретением.
Обычно воздушная сушка достигается путем подвешивания табачного листа в хорошо проветриваемых помещениях и высушивания. Это обычно выполняется в течение четырех-восьми недель. Воздушная сушка особенно подходит для табака Берли.
Соответственно, табачный лист может быть высушен огневой сушкой. Огневая сушка обычно достигается путем подвешивания табачного листа в больших помещениях, где огонь твердой древесины поддерживается на непрерывной или периодической низкой температуре тления и обычно занимает от трех дней до десяти недель, в зависимости от процесса и табака.
В другом воплощении табачный лист может быть высушен дымовой сушкой. Дымовая сушка может включать натягивание табачных листьев на палки для навешивания пучков табачных листьев и подвешивание их на ярусах в сушильных помещениях. У помещения обычно есть дымоход, который поступает от внешних топочных камер. Как правило, это приводит к тому, что табак подвергался термосушке без воздействия дыма. Обычно температура будет медленно повышаться в течение процесса сушки, причем весь процесс занимает около 1 недели.
Соответственно, табачный лист может быть высушен на солнце. Этот метод обычно включает в себя воздействие открытого табака солнцу.
Соответственно обработанный табачный лист может быть обработан путем ферментации. Ферментация может быть проведена любым способом, известным в данной области. Обычно во время ферментации листья табака складывают в стопки (большую часть) высушенного табака, покрытые, например, мешковиной, чтобы сохранить влагу. Комбинация оставшейся воды внутри листа и масса табака генерируют естественное тепло, которое способствует созреванию табака. Температура в центре массы контролируется ежедневно. В некоторых способах каждую неделю всю массу открывают. Листья затем удаляются, чтобы встряхнуть и увлажнить, и массу вращают так, чтобы внутренние листья выходили наружу, а нижние листья размещались на вершине массы. Это обеспечивает равномерную ферментацию по всей массе. Дополнительная влага на листьях, а также фактическое вращение самих листьев генерируют тепло, выделяя природный аммиак табака и снижая никотин, а также углубляя цвет и улучшая аромат табака. Обычно процесс ферментации продолжается до 6 месяцев, в зависимости от сорта табака, положения стебля на листе, толщины и предполагаемого применения листа.
Соответственно обработанный табачный лист может быть обработан пастеризацией. Пастеризация может быть особенно предпочтительной, когда табачный лист будет использоваться для изготовления бездымного табачного продукта, наиболее предпочтительно снюса. Пастеризацию листьев табака можно проводить любым способом, известным в данной области. Например, пастеризация может быть проведена, как описано в J Foulds, L Ramstrom, M Burke, K Fagerstrom. Effect of smokeless tobacco (snus) on smoking and public health in Sweden. Tobacc Control (2003) 12: 349–359, идеи которого включены в настоящее описание ссылкой.
Во время производства снюса пастеризация обычно осуществляется с помощью процесса, в котором табак подвергается тепловой обработке паром в течение 24–36 часов (температура достигает около 100°C). Это приводит к почти стерильному продукту, и, не желая быть связанными какой-либо теорией, считаем, что одним из последствий этого является ограничение дальнейшего образования TSNA.
В одном воплощении пастеризация может быть паровой пастеризацией.
В некоторых воплощениях обработанный табачный лист может быть разрезан. Обработанный табачный лист может быть разрезан до или после обработки. Соответственно, обработанный табачный лист может быть разрезан после обработки.
В некоторых воплощениях растение табака, собранный лист растения табака и/или обработанный лист табака могут быть использованы для экстракции никотина. Экстракция никотина может быть достигнута с применением любого способа, известного в данной области. Например, способ извлечения никотина из табака описан в патенте US 2162738, который включен в данное описание ссылкой.
В одном аспекте настоящее изобретение относится к высушенному табачному материалу, изготовленному из растения табака или его части в соответствии с изобретением или из культуры клеток табака в соответствии с настоящим изобретением.
В другом аспекте настоящее изобретение относится к табачной смеси, содержащей табачный материал, изготовленный из растения табака или его части в соответствии с настоящим изобретением или из культуры клеток табака в соответствии с настоящим изобретением. В одном аспекте настоящее изобретение относится к табачной смеси, содержащей высушенный табачный материал согласно настоящему изобретению.
Соответственно, табачная смесь согласно настоящему изобретению может содержать около 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% табака из растения табака или его части в соответствии с настоящего изобретения или из культуры клеток табака в соответствии с настоящим изобретением. Соответственно, табачная смесь может содержать около 10% табака из растения табака или его части согласно настоящему изобретению или из культуры клеток табака согласно настоящему изобретению. Соответственно, табачная смесь может содержать около 20% табака из растения табака или его части согласно настоящему изобретению или из культуры клеток табака согласно настоящему изобретению. Соответственно, табачная смесь может содержать около 30% табака из растения табака или его части согласно настоящему изобретению или из культуры клеток табака согласно настоящему изобретению. Соответственно, табачная смесь может содержать около 40% табака из растения табака или его части согласно настоящему изобретению или из культуры клеток табака согласно настоящему изобретению. Соответственно, табачная смесь может содержать около 50% табака из растения табака или его части согласно настоящему изобретению или из культуры клеток табака согласно настоящему изобретению. Соответственно, табачная смесь может содержать около 60% табака из растения табака или его части согласно настоящему изобретению или из культуры клеток табака согласно настоящему изобретению. Соответственно, табачная смесь может содержать около 70% табака из растения табака или его части согласно настоящему изобретению или из культуры клеток табака согласно настоящему изобретению. Соответственно, табачная смесь может содержать около 80% табака из растения табака или его части согласно настоящему изобретению или из культуры клеток табака согласно настоящему изобретению. Соответственно, табачная смесь может содержать около 90% табака из растения табака или его части согласно настоящему изобретению или из культуры клеток табака согласно настоящему изобретению.
В одном аспекте табачная смесь согласно настоящему изобретению содержит, по меньшей мере, около 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 95 процентов от сухой массы табака, высушенного из растения табака, или его части согласно настоящему изобретению или культуры клеток табака согласно настоящему изобретению.
Соответственно, высушенный табачный материал может быть высушен воздушной сушкой. Соответственно, высушенный табачный материал может быть высушен дымовой сушкой. Соответственно, высушенный табачный материал может быть высушен на солнце.
Табачный продукт или курительное изделие в соответствии с настоящим изобретением может содержать табачный материал (например, высушенный табачный материал) в соответствии с настоящим изобретением.
В другом аспекте в настоящем изобретении предложен табачный продукт. Табачный продукт согласно настоящему изобретению может представлять собой смешанный табачный продукт. В одном воплощении табачный продукт может быть приготовлен из растения табака по изобретению или его части. В одном воплощении табачный продукт может быть получен из растения табака, которое ингибировало активность или экспрессию гена синтеза дитерпена и повышенное содержание сложного эфира сахарозы. Табачный продукт может иметь пониженную активность гена синтеза дитерпена или экспрессию и повышенное содержание сложного эфира сахарозы. Соответственно, растение табака или его часть может размножаться из материала для размножения растения табака согласно настоящему изобретению.
Термин «его часть», используемый в данном документе в контексте растения табака, относится к части растения табака. Предпочтительно «его часть» представляет собой лист растения табака.
В другом воплощении табачный продукт может быть приготовлен из собранного листа по изобретению. В дополнительном воплощении табачный продукт может быть приготовлен из обработанного табачного листа по изобретению. Соответственно, табачный продукт может быть приготовлен из табачного листа, обработанного одним или более способами: сушки, ферментации и/или пастеризации. Соответственно, табачный продукт может содержать нарезанный табачный лист, необязательно обработанный в соответствии с вышеизложенным воплощением.
В одном воплощении табачный продукт может быть курительным изделием. Используемый в данном документе термин «курительное изделие» может включать курительные изделия, такие как гранулированный табак, сигареты, сигары и сигариллы на основе табака, производные табака, взорванный табак, восстановленный табак или заменители табака.
В другом воплощении табачный продукт может представлять собой бездымный табачный продукт. Используемый в данном документе термин «бездымный табачный продукт» относится к табачному продукту, который не предназначен для курения и/или сжигания. В одном воплощении бездымный табачный продукт может включать снюс, нюхательный табак, жевательный табак или тому подобное.
В дополнительном воплощении табачный продукт может представлять собой устройство для нагревания табака или гибридное устройство, или электронные сигареты, или тому подобное. Обычно в нагревающих устройствах или гибридных устройствах аэрозоль генерируется путем передачи тепла от источника тепла к физически раздельному аэрозолеобразующему субстрату или материалу, который может быть расположен внутри, вокруг или ниже источника нагревания. Во время курения летучие соединения высвобождаются из аэрозолеобразующего субстрата за счет передачи тепла от источника тепла и уносятся воздухом, разрываемым через курительное изделие. По мере того как высвобождаемые соединения охлаждаются, они конденсируются, образуя аэрозоль, который вдыхается пользователем.
Генерирующие аэрозоль изделия и устройства для потребления или нагревающие устройства для курения табака известны в данной области. Они могут включать, например, устройства с электрическим подогревом, генерирующие аэрозоль, в которых аэрозоль генерируется путем передачи тепла от одного или более электрических нагревательных элементов устройства, генерирующего аэрозоль, к аэрозолеобразующей подложке устройства для нагрева табака. Подходящим устройством для нагревания табака может быть устройство, генерирующее аэрозоль.
Предпочтительно устройство для нагревания табака может представлять собой устройство для нагревания без сжигания (heat-not-burn). Устройства для нагревания без сжигания известны в данной области техники и выделяют соединения путем нагревания, но не сжигания табака. Примером подходящего устройства для нагревания без сжигания может быть то, которое описано в WO 2013/034459 или GB2515502, которые включены в данное описание ссылкой.
В одном воплощении аэрозолеобразующий субстрат устройства для нагревания табака может представлять собой табачный продукт в соответствии с настоящим изобретением.
В одном воплощении устройство для нагревания табака может быть гибридным устройством.
Полинуклеотиды/полипептиды/конструкции
В некоторых воплощениях настоящего изобретения конструкции, которые ингибируют активность или экспрессию гена синтеза дитерпена, могут быть трансформированы в растительные клетки под управлением промотора. В некоторых воплощениях настоящего изобретения ДНК-конструкции ddRNAi, которые экспрессируют dsRNA или amiRNA для ингибирования активности или экспрессии гена синтеза дитерпена, могут трансформироваться в растительные клетки под управлением промотора.
Конструкции могут быть введены в растения согласно настоящему изобретению посредством подходящего вектора, например, векторов трансформации растений. Вектор трансформации растения может содержать экспрессирующую кассету, содержащую в направлении транскрипции 5'-3' последовательность промотора, последовательность ДНК-конструкции ddRNAi, нацеленную на ген синтеза дитерпена, предпочтительно нацеленную на ген циклазы 2 (CYC2), CBTol-циклазы или терпен-синтазы 3-8, необязательно, включая интроны, и, необязательно, 3'-нетранслируемую последовательность терминации, включающую стоп-сигнал для РНК-полимеразы и сигнал полиаденилирования для полиаденилазы. Промоторная последовательность может присутствовать в одной или более копиях, и такие копии могут быть идентичными или вариантами промоторной последовательности, как описано выше. Терминаторная последовательность может быть получена из растительных, бактериальных или вирусных генов. Подходящие терминаторные последовательности представляют собой, например, последовательность терминатора rbcS E9 гороха, последовательность терминатора nos, полученную из гена нопалинсинтазы Agrobacterium tumefaciens, и последовательность терминатора 35S из вируса мозаики цветной капусты. Специалист в данной области техники осведомлен о других подходящих терминаторных последовательностях.
Конструкция согласно настоящему изобретению может также включать механизм усиления экспрессии гена для увеличения силы промотора. Примером такого энхансерного элемента является тот, который получен из части промотора гена пластоцианина гороха, и который является объектом международной патентной заявки № WO 97/20056, которая включена в настоящий документ ссылкой. Подходящими энхансерными элементами могут быть, например, энхансерный элемент nos, полученный из гена нопалин-синтазы Agrobacterium tumefaciens, и энхансерный элемент 35S из вируса мозаики цветной капусты.
Эти регуляторные области могут быть получены из того же гена, что и последовательность промоторной ДНК, или могут быть получены из разных генов, из Nicotiana tabacum или других организмов, например, из растения семейства Solanaceae или из подсемейства Cestroideae. Все регуляторные области должны быть способны функционировать в клетках трансформируемой ткани.
Последовательность ДНК промотора может быть получена из того же гена, что и ген, представляющий интерес (например, ген, которым будет управлять промотор, например, ген, кодирующий дерегулированную нитратредуктазу по изобретению), кодирующую последовательность, используемую в настоящем изобретении, или может относиться к другому гену из Nicotiana tabacum или другого организма, например, из растения семейства Solanaceae или из подсемейства Cestroideae.
Экспрессирующая кассета может быть включена в основной вектор трансформации растений, такой как pBIN 19 Plus, pBI 101, pKYLX71: 35S2, pCAMBIA2300 или другие подходящие векторы трансформации растений, известные в данной области. В дополнение к экспрессирующей кассете вектор трансформации растения будет содержать такие последовательности, которые необходимы для процесса трансформации. Они могут включать в себя гены Agrobacterium vir, одну или более граничных последовательностей Т-ДНК и маркер селекции или другие средства идентификации трансгенных растительных клеток.
Термин «вектор трансформации растения» означает конструкцию, способную к экспрессии in vivo или in vitro. Предпочтительно, чтобы экспрессирующий вектор был включен в геном организма. Термин «включенный» предпочтительно охватывает стабильное включение в геном.
Способы трансформации растений хорошо известны в данной области и включают, например, опосредуемую Agrobacterium трансформацию. Основной принцип в строительстве генетически модифицированных растений заключается во вставке генетической информации в геном растения, чтобы обеспечить стабильное поддержание встроенного генетического материала. Обзор общих методов можно найти в статьях Potrykus (Annu Rev Plant Physiol Plant Mol Biol [1991] 42: 205-225) и Christon (AgroFood-Industry Hi-Tech March/April1994 17-27), которые включены в настоящее описание ссылкой.
Как правило, при трансформации, опосредованной Agrobacterium, бинарный вектор, несущий интересующую чужеродную ДНК, то есть ДНК-конструкцию ddRNAi, переносится из соответствующего штамма Agrobacterium в целевое растение путем совместного культивирования Agrobacterium с эксплантами из целевого растения. Трансформированную растительную ткань затем регенерируют на селекционной среде, причем селекционная среда содержит селектируемый маркер и гормоны роста растений. Альтернативой является способ погружения цветов (Clough & Bent, 1998 Plant J. 1998 Dec; 16 (6): 735-43, который включен в данное описание ссылкой), посредством которого цветочные почки интактного растения приводят в контакт с суспензией штамма Agrobacterium, содержащий химерный ген, и после образования семян, трансформированные особи проращивают и идентифицируют по росту на селективной среде. Прямая инфекция тканей растений Agrobacterium является простой техникой, которая широко используется и которая описана в публикации Butcher DN et al., (1980), Tissue Culture Methods for Plant Pathologists, eds.: DS Ingrams и JP Helgeson, 203-208. которая включена в настоящее описание ссылкой.
Другие подходящие способы трансформации включают прямой перенос гена в протопласты с применением методов с полиэтиленгликолем или электропорации, бомбардировки частицами, микроинъекции и использования, например, волокон карбида кремния. Трансформирующие установки баллистической трансформации, включая метод с карбидокремниевыми волокнами, описаны в Frame BR, Drayton PR, Bagnaall SV, Lewnau CJ, Bullock WP, Wilson HM, Dunwell JM, Thompson JA & Wang K (1994), который включен в данное описание ссылкой. Получение фертильных трансгенный растений кукурузы с помощью трансформации, опосредованной карбидом кремния, описано в The Plant Journal 6: 941-948, которая включена в данное описание ссылкой), а методы вирусной трансформации - в Meyer P, Heidmmm I & Niedenhof I (1992). The use of cassava mosaic virus as a vector system for plants is taught in Gene 110: 213-217, которая включена в данное описание ссылкой. Дополнительные указания по трансформации растений можно найти в EP-A-0449375, включенном в настоящий документ ссылкой.
В дополнительном аспекте настоящее изобретение относится к векторной системе, которая несет ДНК-конструкцию ddRNAi и введение ее в геном организма, такого как растение. Векторная система может содержать один вектор, но она может содержать два вектора. В случае двух векторов векторную систему обычно называют бинарной векторной системой. Бинарные векторные системы описаны более подробно в Gynheung Anetal, (1980), Binary Vectors, Plant Molecular Biology Manual A3, 1-19, которое включено в настоящий документ ссылкой.
В одной широко используемой системе трансформации растительных клеток используется плазмида Ti из Agrobacterium tumefaciens или плазмида Ri из Agrobacterium rhizogenes Anetal., (1986), Plant Physiol. 81, 301-305 и Butcher DN et al., (1980), Tissue Culture Methods for Plant Pathologists, eds.: DS Ingrams и JP Helgeson, 203-208, которые включены в данное описание ссылкой. После каждого способа введения желаемого экзогенного гена согласно настоящему изобретению в растения может возникнуть необходимость в присутствии и/или вставке дополнительных последовательностей ДНК. Применение Т-ДНК для трансформации растительных клеток интенсивно изучалось и описано в EP-A-120516; Hoekema, in: The Binary Plant Vector System Offset-drukkerij Kanters B.B., Amsterdam, 1985, Chapter V; Fraley, et al., Crit.Rev. Plant Sci., 4: 1-46; и Anetal., EMBO J (1985) 4: 277-284, включенных в настоящее описание ссылкой.
Растительные клетки, трансформированные ДНК-конструкцией ddRNAi, которая экспрессирует dsRNA или amiRNA, которая ингибирует активность или экспрессию гена синтеза дитерпена, можно выращивать и поддерживать в соответствии с хорошо известными культуральными способами, такими как культивирование клеток в подходящей культуральной среде, поставляемой в комплекте с необходимыми факторами роста, такими как аминокислоты, гормоны растений, витамины и т.д.
Термин «трансгенное растение» по отношению к настоящему изобретению включает любое растение, которое содержит ДНК-конструкцию ddRNAi по изобретению. Соответственно, трансгенное растение представляет собой растение, которое было трансформировано ДНК-конструкцией ddRNAi согласно изобретению. Предпочтительно трансгенное растение проявляет ингибированную активность или экспрессию гена синтеза дитерпена и повышенное содержание сложного эфира сахарозы в соответствии с настоящим изобретением. Термин «трансгенное растение» не охватывает последовательности, кодирующие нативные нуклеотиды, в их естественной среде, когда они находятся под контролем их нативного промотора, который также находится в его естественной среде.
В одном аспекте ДНК-конструкция ddRNAi, вектор трансформации растения или растительная клетка в соответствии с настоящим изобретением находятся в выделенной форме. Термин «выделенный» означает, что последовательность, по меньшей мере, по существу, свободна, по меньшей мере, от одного другого компонента, с которым последовательность естественным образом связана в природе и с которым она обнаруживается в природе.
В одном аспекте ДНК-конструкция ddRNAi, вектор трансформации растения или растительная клетка по изобретению находятся в очищенной форме. Термин «очищенный» означает относительно чистое состояние, например, по меньшей мере, около 90% чистоты, или, по меньшей мере, около 95% чистоты, или, по меньшей мере, около 98% чистоты.
Используемый в данном документе термин «нуклеотидная последовательность» относится к олигонуклеотидной последовательности или полинуклеотидной последовательности, а также к варианту, гомологам, фрагментам и их производным (таким как их части). Нуклеотидная последовательность может иметь геномное, синтетическое или рекомбинантное происхождение, которая может быть двухцепочечной или одноцепочечной, независимо от того, представляет ли она смысловую или антисмысловую цепь.
Термин «нуклеотидная последовательность» по отношению к настоящему изобретению включает геномную ДНК, кДНК, синтетическую ДНК и РНК. Предпочтительно это означает ДНК, более предпочтительно последовательность кДНК, кодирующую настоящее изобретение.
В предпочтительном воплощении изобретения нуклеотидная последовательность, когда она относится к настоящему изобретению и когда охвачена объемом настоящего изобретения, т.е. ген синтеза дитерпена, включает нативную нуклеотидную последовательность, когда это имеет место в его естественной среде, и, когда она связана с его естественно ассоциированной последовательностью (последовательностями), которая(которые) также находятся в своей/их естественной среде. Для удобства ссылок мы будем называть это предпочтительное воплощение «нативной нуклеотидной последовательностью». В этом отношении термин «нативная нуклеотидная последовательность» означает цельную нуклеотидную последовательность, которая находится в ее нативном окружении и когда она функционально связана со всем промотором, с которым она естественным образом связана, этот промотор также находится в своем нативном окружении.
Как правило, молекула RNAi, охватываемая объемом настоящего изобретения, получена с применением методов рекомбинантной ДНК (т.е. рекомбинантной ДНК). Однако в альтернативном воплощении изобретения нуклеотидную последовательность можно синтезировать, полностью или частично, с применением химических методов, хорошо известных в данной области (см. Caruthers MH et al., (1980) Nuc Acids Res Symp Ser 215-23 и Horn T et al., (1980) Nuc Acids Res Symp Ser 225-232, которые включены в настоящее описание ссылкой).
Нуклеотидная последовательность, кодирующая либо белок, который обладает специфическими свойствами в качестве гена синтеза дитерпена, как определено в настоящем документе, либо белок, который подходит для модификации, может быть идентифицирована и/или выделена и/или очищена из любой клетки или организма, продуцирующих указанный белок. В данной области хорошо известны различные способы идентификации и/или выделения и/или очистки нуклеотидных последовательностей. Например, методы ПЦР-амплификации для получения большего количества последовательности могут быть использованы после того, как подходящая последовательность была идентифицирована и/или выделена и/или очищена.
В качестве дополнительного примера, библиотека геномной ДНК и/или кДНК может быть сконструирована с применением хромосомной ДНК или информационной РНК из организма, продуцирующего фермент. Если аминокислотная последовательность фермента известна, меченые олигонуклеотидные зонды могут быть синтезированы и использованы для идентификации клонов, кодирующих фермент, из геномной библиотеки, полученной из организма. Альтернативно, меченый олигонуклеотидный зонд, содержащий последовательности, гомологичные другому известному гену фермента, можно использовать для идентификации клонов, кодирующих фермент в последнем случае используются условия гибридизации и отмывки более низкой жесткости.
Еще в одном варианте нуклеотидная последовательность, кодирующая фермент, может быть получена синтетическим путем с помощью общепринятых стандартных методов, например, метода фосфороамидита, описанного в статье Beucage SL et al., (1981) Tetrahedron Letters 22, p 1859-1869, которая включена в настоящее описание ссылкой, или способа, описанного в статье Matthes et al., (1984) EMBO J. 3, р 801-805, который включен в настоящий документ ссылкой. В фосфороамидатном способе олигонуклеотиды синтезируют, например, в автоматическом синтезаторе ДНК, очищают, отжигают, лигируют и клонируют в соответствующих векторах.
Используемый в данном документе термин «аминокислотная последовательность» является синонимом термина «полипептид» и/или термина «белок». В некоторых случаях термин «аминокислотная последовательность» является синонимом термина «пептид». В некоторых случаях термин «аминокислотная последовательность» является синонимом термина «фермент».
Настоящее изобретение также охватывает применение последовательностей, имеющих степень идентичности последовательности или гомологию последовательности с аминокислотной последовательностью (последовательностями) полипептида, имеющего определенные свойства, определенные в данном документе, или любой нуклеотидной последовательностью, то есть геном синтеза дитерпена, кодирующим такой полипептид (далее упоминаемый в данном документе) как «гомологичная(ые) последовательность(и)»). в данном документе термин «гомолог» означает объект, имеющий определенную гомологию с аминокислотными последовательностями объекта и нуклеотидными последовательностями объекта. В данном документе термин «гомология» можно отождествить с «идентичностью».
Гомологичная аминокислотная последовательность и/или нуклеотидная последовательность и/или фрагменты должны обеспечивать и/или кодировать полипептид, который сохраняет функциональную активность и/или усиливает активность гена синтеза дитерпена. Как правило, гомологичные последовательности будут содержать те же активные сайты и т.д., что и, например, рассматриваемая аминокислотная последовательность, или будут кодировать те же активные сайты. Хотя гомологию также можно рассматривать с точки зрения сходства (т.е. аминокислотные остатки, имеющие сходные химические свойства/функции), в контексте настоящего изобретения предпочтительно выражать гомологию с точки зрения идентичности последовательности.
В одном воплощении изобретения гомологичная последовательность включает аминокислотную последовательность или нуклеотидную последовательность, которая имеет одну или более вставок, делеций и/или замен по сравнению с рассматриваемой последовательностью.
Сравнение гомологии или идентичности может проводиться визуально или, чаще, с помощью легко доступных программ сравнения последовательностей. Эти коммерчески доступные компьютерные программы могут рассчитывать % гомологии между двумя или более последовательностями. % гомологии или % идентичности могут быть рассчитаны для смежных последовательностей, то есть одна последовательность выровнена с другой последовательностью, и каждая аминокислота в одной последовательности напрямую сравнивается с соответствующей аминокислотой в другой последовательности, по одному остатку за раз. Это называется выравниванием «без разрывов». Как правило, такие выравнивания «без разрывов» выполняются только на относительно небольшом количестве остатков.
Хотя это очень простой и последовательный способ, он не учитывает, что, например, в другой идентичной паре последовательностей одна вставка или делеция приведет к тому, что следующие аминокислотные остатки будут выровнены, что потенциально может привести к большему снижению % гомологии, когда выполняется глобальное выравнивание. Следовательно, большинство способов сравнения последовательностей предназначены для получения оптимальных выравниваний, учитывающих возможные вставки и делеции, без чрезмерного ухудшения общего показателя гомологии. Это достигается путем вставки «разрывов» в выравнивание последовательности, чтобы попытаться максимизировать локальную гомологию.
Однако эти более сложные способы назначают «штрафы за разрывы» для каждого разрыва, возникающего при выравнивании, чтобы при одинаковом количестве идентичных аминокислот выравнивание последовательностей происходило с минимально возможным количеством разрывов, что отражает то, что более высокая степень родства между двумя сравниваемыми последовательностями достигнет более высокой балльной оценки, чем в случае, когда много разрывов. Обычно используются «цена за аффинный разрыв», которая взимает относительно высокую цену за существование разрыва и меньший штраф за каждый последующий остаток в разрыве. Это наиболее часто используемая система оценки разрывов. Высокие штрафы за разрывы, конечно, приведут к оптимизированным выравниваниям с меньшим количеством разрывов. Большинство программ выравнивания позволяют изменять штрафы за разрывы. Однако предпочтительно использовать значения по умолчанию при использовании такого программного обеспечения для сравнения последовательностей.
Поэтому расчет максимального % гомологии, во-первых, требует получения оптимального выравнивания с учетом штрафов за разрывы. Подходящей компьютерной программой для проведения такого выравнивания является Vector NTI (Invitrogen Corp.). Примеры программного обеспечения, которое может выполнять сравнение последовательностей, включают, без ограничения указанным, пакет BLAST (см. Ausubel et al. 1999 Short Protocols in Molecular Biology, 4th Ed - Chapter 18), BLAST 2 (см. FEMS Microbiol Lett 1999 174 (2): 247-50; FEMS Microbiol Lett 1999 177 (1): 187-8 и tatiana@ncbi.nlm. nih.gov), FASTA (Altschul et al. 1990 J. Mol. Biol. 403-410) и AlignX, например, по меньшей мере, BLAST, BLAST 2 и FASTA доступны для автономного и онлайн-поиска (см. Ausubel et al. 1999, стр. с 7-58 по 7-60).
Хотя конечный процент гомологии может быть измерен с точки зрения идентичности, сам процесс выравнивания обычно не основан на сравнении пар «все или ничего». Вместо этого обычно используется масштабированная матрица баллов сходства, которая присваивает баллы каждому парному сравнению на основе химического сходства или эволюционного расстояния. Примером такой широко используемой матрицы является матрица BLOSUM62 - матрица по умолчанию для набора программ BLAST. Программы Vector NTI обычно используют либо общедоступные значения по умолчанию, либо пользовательскую таблицу сравнения символов, если она имеется (см. Руководство пользователя для получения дополнительной информации). Для некоторых приложений предпочтительно использовать значения по умолчанию для пакета Vector NTI.
Альтернативно, процентные гомологии могут быть рассчитаны с применением функции множественного выравнивания в Vector NTI (Invitrogen Corp.) на основе алгоритма, аналогичного CLUSTAL (Higgins DG & Sharp PM (1988), Gene 73 (1), 237-244). Как только программное обеспечение произведет оптимальное выравнивание, можно рассчитать процент гомологии, предпочтительно процент идентичности последовательности. Программное обеспечение обычно делает это как часть сравнения последовательностей и генерирует числовой результат.
Если при определении идентичности последовательности используются штрафы за разрыв, то для парного выравнивания предпочтительно использовать следующие параметры:
В одном воплощении CLUSTAL может использоваться с штрафом за разрывы и удлинение разрыва, установленным, как определено выше. В некоторых воплощениях штрафы за разрывы, используемые для выравнивания BLAST или CLUSTAL, могут отличаться от тех, которые подробно описаны выше. Специалисту будет понятно, что стандартные параметры для выполнения выравниваний BLAST и CLUSTAL могут периодически меняться, и сможет выбрать соответствующие параметры на основе стандартных параметров, подробно описанных для алгоритмов выравнивания BLAST или CLUSTAL в данный момент.
Соответственно, степень идентичности в отношении нуклеотидной последовательности определяют, по меньшей мере, для 20 смежных нуклеотидов, предпочтительно, по меньшей мере, для 30 смежных нуклеотидов, предпочтительно, по меньшей мере, для 40 смежных нуклеотидов, предпочтительно, по меньшей мере, для 50 смежных нуклеотидов, предпочтительно, по меньшей мере, для более 60 смежных нуклеотидов, предпочтительно, по меньшей мере, для более 100 смежных нуклеотидов; где нуклеотидная последовательность кодирует ДНК-конструкцию ddRNAi. Степень идентичности в отношении нуклеотидной последовательности определяют, по меньшей мере, по 10 смежным нуклеотидам, предпочтительно по 20 смежным нуклеотидам, предпочтительно, по меньшей мере, по 30 смежным нуклеотидам, предпочтительно, по меньшей мере, по 40 смежным нуклеотидам, предпочтительно, по меньшей мере, по 50 смежным нуклеотидам, предпочтительно, по меньшей мере, более 60 смежных нуклеотидов, предпочтительно, по меньшей мере, более 100 смежных нуклеотидов.
Соответственно, степень идентичности в отношении нуклеотидной последовательности может быть определена по всей последовательности.
Последовательности также могут иметь делеции, вставки или замены аминокислотных остатков, которые вызывают молчащие изменения и приводят к функционально эквивалентному веществу. Умышленные аминокислотные замены могут быть сделаны на основе сходства в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфипатической природе остатков, при условии, что сохраняется вторичная связывающая активность вещества. Например, отрицательно заряженные аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают лизин и аргинин; и аминокислоты с незаряженными полярными головными группами, имеющие сходные значения гидрофильности, включают лейцин, изолейцин, валин, глицин, аланин, аспарагин, глутамин, серин, треонин, фенилаланин и тирозин.
Консервативные замены могут быть сделаны, например, согласно таблице ниже. Аминокислоты в одном и том же блоке во втором столбце и предпочтительно в одной и той же строке в третьем столбце могут заменять друг друга:
Настоящее изобретение также охватывает гомологичное замещение (оба замещения и замены используются в данном документе для обозначения взаимозамены существующего аминокислотного остатка на альтернативный остаток), которое может происходить, то есть аналогичное замещение, такое как основное для основного, кислое для кислого полярное для полярного и т.д. Также может происходить негомологичное замещение, то есть остаток одного класса на остаток другого класса или, альтернативно, оно содержит включение искусственных аминокислот, таких как орнитин (далее называемый Z), орнитин диаминомасляной кислоты (далее называемый B), норлейцин, орнитин (далее называемый О), пирилаланин, тиенилаланин, нафтилаланин и фенилглицин.
Замены, которые также могут быть сделаны искусственными аминокислотами, включают; альфа* и альфа-дизамещенные* аминокислоты, N-алкиламинокислоты*, молочную кислоту*, галогенидные производные природных аминокислот, такие как трифтортирозин*, p-Cl-фенилаланин*, p-Br-фенилаланин*, p-фенилаланин*, L-аллил-глицин*, бета-аланин*, L-α-аминомасляная кислота*, L-γ-аминомасляная кислота*, L-α-аминоизомасляная кислота*, L-ε-аминокапроновая кислота#, 7-аминогептановая кислота*, L-метионинсульфон#*, L-норлейцин*, L-норвалин*, п-нитро-L-фенилаланин*, L-гидроксипролин#, L-тиопролин*, метилпроизводные фенилаланина (Phe), такие как 4-метил-Phe*, пентаметил-Phe*, L-Phe (4-амино)#, L-Tyr (метил)*, L-Phe (4-изопропил)*, L-Tic (1,2,3), 4-тетрагидроизохинолин-3-карбоновая кислота)*, L-диаминопропионовая кислота# и L-Phe (4-бензил)*. Обозначение * было использовано для целей обсуждения выше (касающегося гомологичного или негомологичного замещения), чтобы указать на гидрофобную природу производного, тогда как # был использован для указания на гидрофильную природу производного, #* означает амфипатические характеристики.
Вариантные аминокислотные последовательности могут включать подходящие спейсерные группы, которые могут быть вставлены между любыми двумя аминокислотными остатками последовательности, включая алкильные группы, такие как метильные, этильные или пропильные группы, в дополнение к аминокислотным спейсерам, таким как остатки глицина или β-аланина. Другая форма вариации включает присутствие одного или более аминокислотных остатков в пептоидной форме, что будет хорошо понятно специалистам в данной области. Во избежании сомнений, «пептоидная форма» используются для обозначения остатков аминокислот варианта, в котором α-углерод-замещающая группа находится в положении атома азота, а не α-углерода. Способы получения пептидов в пептоидной форме известны в данной области, например, Simon RJ et al., PNAS (1992) 89 (20), 9367-9371 и Horwell DC, Trends Biotechnol. (1995) 13 (4), 132-134.
Нуклеотидные последовательности для применения в настоящем изобретении могут включать синтетические или модифицированные нуклеотиды. Ряд различных типов модификации олигонуклеотидов известен в данной области. Они включают метилфосфонатные и фосфоротиоатные каркасы и/или добавление акридиновых или полилизиновых цепей на 3' и/или 5' концах молекулы. Для целей настоящего изобретения следует понимать, что описанные в данном документе нуклеотидные последовательности могут быть модифицированы любым способом, доступным в данной области. Такие модификации могут быть выполнены для того, чтобы повысить активность in vivo или продолжительность жизни нуклеотидных последовательностей согласно настоящему изобретению.
Настоящее изобретение также охватывает последовательности, которые являются комплементарными последовательностям нуклеиновой кислоты согласно настоящему изобретению, или последовательности, которые способны гибридизоваться либо с последовательностями согласно настоящему изобретению, либо с последовательностями, которые являются комплементарными им. Термин «гибридизация», используемый в данном документе, должен включать «процесс, посредством которого цепь нуклеиновой кислоты соединяется с комплементарной цепью посредством спаривания оснований», а также процесс амплификации, проводимый в технологиях полимеразной цепной реакции (ПЦР).
Настоящее изобретение также относится к нуклеотидным последовательностям, которые могут гибридизоваться с нуклеотидными последовательностями согласно настоящему изобретению (включая комплементарные последовательности из представленных в данном документе). Предпочтительно гибридизацию определяют в жестких условиях (например, 50°C и 0,2 × SSC × SSC = 0,15 М NaCl, 0,015 М Na3 цитрат, рН 7,0). Более предпочтительно, гибридизацию определяют в условиях высокой жесткости (например, 65°C и 0,1 × SSC × SSC = 0,15 М NaCl, 0,015 М Na3 цитрат, рН 7,0).
В одном аспекте последовательность для применения в настоящем изобретении представляет собой синтетическую последовательность, то есть последовательность, которая была получена химическим или ферментативным синтезом in vitro. Он включает, без ограничения указанным, последовательности, сделанные с оптимальным применением кодонов для организмов-хозяев.
Термин «экспрессирующий вектор» означает конструкцию, способную к экспрессии in vivo или in vitro. В одном воплощении вектор согласно настоящему изобретению экспрессирует dsRNA. В одном воплощении вектор согласно настоящему изобретению экспрессирует amiRNA. Предпочтительно, экспрессирующий вектор включен в геном подходящего организма-хозяина. Термин «включенный» предпочтительно охватывает стабильное включение в геном.
Нуклеотидная последовательность согласно настоящему изобретению может присутствовать в векторе, в котором нуклеотидная последовательность функционально связана с регуляторными последовательностями, способными обеспечивать экспрессию нуклеотидной последовательности подходящим организмом-хозяином. ДНК-конструкции ddRNAi для применения в настоящем изобретении могут быть трансформированы в подходящую клетку-хозяина, как описано в настоящем документе, для обеспечения экспрессии полипептида согласно настоящему изобретению. Выбор вектора, например, плазмидного, космидного или фагового вектора, часто будет зависеть от клетки-хозяина, в которую он должен быть введен. Векторы могут быть использованы in vitro, например, для продуцирования РНК или использованы для трансфекции, трансформации, трансдукции или инфицирования клетки-хозяина.
Таким образом, в дополнительном воплощении изобретение относится к способу получения нуклеотидных последовательностей согласно настоящему изобретению путем введения нуклеотидной последовательности согласно настоящему изобретению в реплицируемый вектор, введения вектора в совместимую клетку-хозяина и выращивания клетки-хозяина в условиях, которые приводят к репликации вектора. Вектор может дополнительно содержать нуклеотидную последовательность, позволяющую вектору реплицироваться в рассматриваемой клетке-хозяине. Примерами таких последовательностей являются последовательности начала репликации плазмид pUC19, pACYC177, pUB110, pE194, pAMB1 и pIJ702.
В некоторых применениях нуклеотидная последовательность для применения в настоящем изобретении функционально связана с регуляторной последовательностью, которая способна обеспечивать экспрессию нуклеотидной последовательности, например, выбранной клеткой-хозяином. Например, настоящее изобретение охватывает вектор, содержащий нуклеотидную последовательность согласно настоящему изобретению, функционально связанную с такой регуляторной последовательностью, то есть вектор является экспрессирующим вектором.
Термин «функционально связанный» относится к расположению, в котором описанные компоненты находятся во взаимосвязи, позволяющей им функционировать по назначению. Регуляторная последовательность, «функционально связанная» с кодирующей последовательностью, лигируется таким образом, что экспрессия кодирующей последовательности достигается в условиях, совместимых с контрольными последовательностями.
Термин «регуляторные последовательности» включает промоторы и энхансеры и другие сигналы регуляции экспрессии. Термин «промотор» используется в обычном смысле, например, сайт связывания РНК-полимеразы. Нуклеотидная последовательность в ДНК-конструкции ddRNAi, которая кодирует dsRNA или amiRNA, может быть функционально связана, по меньшей мере, с промотором.
Термин «конструкция», который является синонимом таких терминов, как «кассета» или «вектор», включает нуклеотидную последовательность для применения в соответствии с настоящим изобретением, прямо или косвенно связанную с промотором.
Примером косвенного присоединения является предоставление подходящей спейсерной группы, такой как последовательность интрона, такого как интрон Sh1 или интрон ADH, промежуточного промотора и нуклеотидной последовательности согласно настоящему изобретению. То же самое относится к термину «слитый» по отношению к настоящему изобретению, который включает в себя прямое или косвенное присоединение. В некоторых случаях термины не охватывают естественную комбинацию нуклеотидной последовательности, кодирующей белок, обычно связанный с промотором гена дикого типа, и когда они оба находятся в их естественной среде. ДНК-конструкция ddRNAi может даже содержать или экспрессировать маркер, который позволяет осуществлять селекцию генетической конструкции.
Обзор общих методов, используемых для трансформации растений, можно найти в статьях Potrykus (Annu Rev Plant Physiol Plant Mol Biol [1991] 42: 205-225) и Christou (Agro-Food-Industry Hi-Tech, March/April 1994 17-27), которые включены в данный документ ссылкой. Дополнительные указания по трансформации растений можно найти в EP-A-0449375, включенном в настоящий документ ссылкой.
Если не указано иное, то все технические и научные термины, используемые в данном документе, имеют такое же значение, которое обычно понимается специалистом в области, к которой принадлежит данное описание. Singleton, et al., DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOGY, 20 ED., John Wiley and Sons, New York (1994), and Hale & Marham, THE HARPER COLLINS DICTIONARY OF BIOLOGY, Harper Perennial, NY (1991) предоставляют специалистам в данной области общий словарь многих терминов, используемых в настоящем раскрытии.
Это раскрытие не ограничено примерными способами и материалами, раскрытыми в данном документе, и любые способы и материалы, подобные или эквивалентные тем, которые описаны в данном документе, могут использоваться на практике или при тестировании воплощений этого раскрытия. Числовые диапазоны включают числа, определяющие диапазон. Если не указано иное, любые последовательности нуклеиновых кислот пишутся слева направо в 5'-3' ориентации; аминокислотные последовательности пишутся слева направо в ориентации от N-конца к С-концу, соответственно.
Заголовки, представленные в данном документе, не являются ограничениями различных аспектов или воплощений этого раскрытия, которые могут иметься со ссылкой на описание в целом. Соответственно, термины, определенные непосредственно ниже, более полно определены ссылкой на описание в целом.
Аминокислоты упоминаются в данном документе с применением названия аминокислоты, трехбуквенного сокращения или однобуквенного сокращения. Используемый в данном документе термин «белок» включает белки, полипептиды и пептиды. Используемый в данном документе термин «аминокислотная последовательность» является синонимом термина «полипептид» и/или термина «белок». В некоторых случаях термин «аминокислотная последовательность» является синонимом термина «пептид». В некоторых случаях термин «аминокислотная последовательность» является синонимом термина «фермент».
Термины «белок» и «полипептид» используются в данном документе взаимозаменяемо. В настоящем описании и формуле изобретения могут использоваться обычные однобуквенные и трехбуквенные коды для аминокислотных остатков. Трехбуквенный код для аминокислот, как определено в соответствии с Совместной комиссией IUPACIUB по биохимической номенклатуре (JCBN). Также понятно, что полипептид может кодироваться более чем одной нуклеотидной последовательностью из-за вырожденности генетического кода.
Другие определения терминов могут встречаться в описании. Прежде чем примерные воплощения будут описаны более подробно, следует понимать, что данное раскрытие не ограничено конкретными описанными воплощениями, которые, конечно, могут варьироваться. Также следует понимать, что используемая в данном документе терминология предназначена только для описания конкретных воплощений и не предназначена для ограничения, так как объем настоящего раскрытия будет ограничен только прилагаемой формулой изобретения.
Когда предоставляется диапазон значений, подразумевается, что каждое промежуточное значение с точностью до десятой доли нижнего предела, если контекст явно не диктует иное, между верхним и нижним пределами этого диапазона также конкретно раскрыт. Каждый меньший диапазон между любым заявленным значением или промежуточным значением в указанном диапазоне и любым другим заявленным или промежуточным значением в этом заявленном диапазоне включен в настоящее раскрытие. Верхние и нижние пределы этих меньших диапазонов могут независимо включаться или исключаться из диапазона, и каждый диапазон, где либо меньшие, либо оба предела включены в меньшие диапазоны, также включен в данное раскрытие при условии любого специально исключенного предела в заявленный диапазон. В тех случаях, когда указанный диапазон включает одно или оба из пределов, диапазоны, исключающие один или оба из этих включенных пределов, также включены в настоящее раскрытие.
Следует отметить, что используемые в данном документе и в прилагаемой формуле изобретения формы единственного числа «а», «an» и «the» включают ссылки на множественное число, если контекст явно не предписывает иное. Таким образом, например, ссылка на «фермент» или «нитратредуктазу» включает в себя множество таких агентов-кандидатов и их эквиваленты, известные специалистам в данной области, и так далее.
Публикации, обсуждаемые в настоящем документе, предназначены исключительно для их раскрытия до даты подачи настоящей заявки. Ничто в данном документе не должно быть истолковано как признание того, что такие публикации составляют предшествующий уровень техники к прилагаемой формуле изобретения.
Преимущества
Преимущественно ингибирование активности или экспрессии гена синтеза дитерпена увеличивает содержание сложного эфира сахарозы в растениях табака. Не желая быть связанными какой-либо теорией, полагают, что уменьшенное применение углерода для получения дитерпенов приводит к увеличению производства сложного эфира сахарозы. Преимущественно способы согласно настоящему изобретению позволяют получать растения табака с предпочтительным химическим составом «турецкого типа», которые представляют собой растения с высоким содержанием биомассы. Преимущественно изобретение обеспечивает желаемый химический состав дыма и химический состав, полезный для продуктов, нагреваемых, но не горючи. Теперь изобретение будет описано только в качестве примера со ссылкой на следующие фигуры и Примеры.
ПРИМЕРЫ
Пример 1. Получение трансгенных конструкций GW1, GW2, GW3, GW5.
Авторы изобретения стремились проанализировать вторичный метаболизм трихом растения и оценить влияние генов, экспрессируемых трихомом, на содержание сложного эфира сахарозы с применением посттранскрипционных стратегий сайленсинга генов (PTGS). Функции гена циклазы 2 (CYC2), CBTol-циклазы и терпен-синтазы 3-8 оценивали с применением RNAi. Способы получения конструкции и регенерации растений подробно описаны в Wang and Wagner 2003 (см. выше, которая включена в данное описание ссылкой) вместе с традиционными методами молекулярного клонирования (Sambrook et al.1989 Molecular cloning: a laboratory manual, Cold Spring Harbor Laboratory Press, New York).
Трансгенная конструкция GW1 - ингибирует экспрессию гена циклазы 2 (CYC2)
Эта ДНК-конструкция ddRNAi была разработана на основе гена циклазы 2 (CYC2). Полная кодирующая последовательность мРНК CYC2 обозначена как AF401234. Геномная последовательность обозначена как AY495694. Конструкция, используемая для создания растений GW1, состоит из: 5'-смыслового фрагмента с 54 по 716-й нуклеотид из последовательности AF401234; частичного фрагмента GUS A в виде петли-шпильки (с 787 по 1812-й нуклеотид), обозначение GUS A соответствует AF502128; и обратной комплементарной последовательности смыслового 3'-фрагмента CYC2. Конструкция содержит нуклеотиды 54-716 из мРНК CYC2 (AF401234) в смысловой ориентации. Эта последовательность соответствует положениям нуклеотидов в геномной последовательности (AY495694): нуклеотиды с 1 по 25, нуклеотиды с 26 по 271 (экзон 1), нуклеотиды с 1253 по 1529 (экзон 2) и первые 115 нуклеотидов 3 экзона (нуклеотиды с 2366 по 2480).). Плазмида pKYLX71-35S2 представляет собой бинарный вектор, используемый для генной конструкции (Wang et al. 2001, см. выше). Последовательность вставки конструкции GW1 изложена в SEQ ID No. 5.
Трансгенная конструкция GW2 - ингибирует экспрессию гена терпен-синтазы 3-8
Эта ДНК-конструкция ddRNAi была разработана на основе первичного транскрипта миРНК168 (EU549055.1; GI: 171195398) (от 5'127 до 535-3'), где 196-216 были заменены на 21 нуклеотид от нуклеотида 1497 в обратной комплементарной последовательности исходной последовательности (Genbank: AY528645, соответствующей положениям нуклеотидов, как показано в SEQ ID NO. 3), и нуклеотиды 279-299 были заменены на 21 нуклеотид из исходной цепи в прямой ориентации, но где 3 основания были модифицированы, когда использовалась область 1497-1517. Модуль, фланкированный фрагментом по сайтам рестрикции 5'-HindIII и 3'-EcoRI, был вставлен между промотором 2X35S и терминатором 35S. Расширенный модуль, который содержал 3X35S пром- miRNAi 3-8 и 35S-терминатор, вводили в сайт множественного клонирования (MCS) бинарного вектора pCAMBIA2300. Рекомбинантный вектор (KmR) вставляли в A. tumefaciens, GV3101 (RifR, GmR). Селекцию колоний проводили по трем антибиотикам (Km, Rif, Gen). Трансформацию Agrobacterium подтверждали ПЦР плазмиды. Последовательность вставки конструкции GW2 представлена в SEQ ID No. 7.
Трансгенная конструкция GW3 - ингибирует экспрессию гена CBTol-циклазы
Эта ДНК-конструкция ddRNAi была разработана на основе гена циклазы N. tabacum (AY049090). Этот двухцепочечный модуль RNAi был собран следующим образом: - частичная последовательность из гена циклазы N. tabacum (AY049090), от 5' 2854 до 4175-3' SEQ ID No. 1 в прямой и обратной ориентации, состоящая из частичного экзона 4, полного интрона 4, экзона 5, интрона 5, экзона 6, интрона 6 и частичного экзона 7. Спейсер между прямым и обратным участками представлял собой частичный ген GUS A, от 5'-786-1816-3'. Плазмида pCAMBIA2300 представляла собой бинарный вектор, используемый для этой генной конструкции. Последовательность вставки конструкции GW3 представлена в SEQ ID No. 6.
Трансгенная конструкция GW5 - ингибирует экспрессию гена терпен-синтазы 3-8
Эта ДНК-конструкция ddRNAi была разработана на основе первичного транскрипта миРНК168 (EU549055.1; GI: 171195398) (от 5'127 до 535-3'), где 196-216 были заменены на 21 нуклеотид от нуклеотида 884 в обратной комплементарной последовательности к исходной последовательности (Genbank: AY528645, соответствующей положениям нуклеотидов, как показано в SEQ ID NO. 3), и нуклеотиды 279-299 были заменены на 21 нуклеотид из исходной цепи в прямой ориентации, но где 4 основания смысловой цепи были модифицированы, когда использовалась область 884-904. Модуль, фланкированный фрагментом по сайтам рестрикции 5'-HindIII и 3'-EcoRI, был вставлен между промотором 2X35S и терминатором 35S. Расширенный модуль, который содержал 3X35S пром-miRNAi 3-8 и 35S-терминатор, вводили в сайт множественного клонирования (MCS) бинарного вектора pCAMBIA2300. Рекомбинантный вектор (KmR) вставляли в A. tumefaciens, GV3101 (RifR, GmR). Селекцию колоний проводили по трем антибиотикам (Km, Rif, Gen). Трансформация Agrobacterium была подтверждена плазмидной ПЦР. Последовательность вставки конструкции GW5 приведена в SEQ ID No. 11.
Пример 2. Агротрансформация и регенерация T.I. 1068
Семена N. tabacum TI 1068 были получены из коллекции семян KTRDC и стерилизованы поверхностно 70% EtOH в течение 1 мин, затем 5% (об./об.) Хлороксом, а затем трижды промыты стерильной водой. Растения выращивали in vitro в контейнерах PLANTCON® (MP Biomedicals, LLC) и использовали в качестве запаса эксплантов для трансформации. Все конструкции были введены в сорт табака T.I. 1068 путем трансформации, опосредованной Agrobacterium, и присутствие трансгенов было подтверждено с помощью ПЦР. Трансформацию A. tumefaciens осуществляли, по существу, как описано в Horsch et al. в Horsch RB, Fry JE, Hoffmann NL, Eichholtz D, Rogers SG, Fraley RT (1985) Science 227: 1229–1231, которая включена в данное описание ссылкой с некоторыми модификациями. Agrobacterium выращивали в течение ночи на среде LB, содержащей 50 мг/л Km, 50 мг/л Rif и 35 мг/л Gm (GW1, GW2,). Для GW3 Km был заменен на Hyg (55 мг/л).
Нарезанные кусочки листьев (1 см 2) от стерильно выращенных растений (возраст 45 дней) инокулировали бактериями (1 × 10 8 КОЕ/мл) в течение 2 дней в темноте, затем промокали досуха и переносили в среду MS с добавлением витаминов Gamborg B-5 (Sigma-Aldrich Co. LTD, Ирвин, Великобритания), 3% сахарозы, 1 мг/л BAP и 0,05 мг/л NAA с добавлением 200 мг/л Km (или 55 мг/л Hyg для GW3) и 400 мг цефотаксима. Примерно через месяц, небольшие проростки были сформированы на краях кусочков и после одного пассажа на той же самой среде, проростки переносили в ту же самую среду, но гормоны были удалены, и антибиотики были уменьшены в два раза. Укорененные растения переносили в удобренную Pro-Mix (Premier Horticulture Inc., Канада) в теплице. Индивидуальные трансформанты анализировали с помощью GC-MS, и те, в которых присутствовали желаемые изменения, высевали самостоятельно для дальнейшего анализа T1-поколения.
Пример 3. Анализ зеленых листьев контрольных и трансгенных линий в полевых условиях.
Подготовка линий для полевых испытаний и дизайн полевых испытаний
Семена из линий GW1 и GW2, полученные в результате самоопыления растений T3 или T2, стерилизовали поверхностно и проращивали in vitro на среде MS с добавлением витаминов Gamborg B-5 (Sigma-Aldrich Co. LTD, Ирвин, Великобритания), 3 % сахарозы и Km (200 мг/л). Семена GW3 стерилизовали поверхностно и проращивали на среде, содержащей Hyg (55 мг/л). Через месяц проростки переносили в плавающие лотки в теплице (удобренные Pro-Mix (Premier Horticulture Inc., Канада)) и выращивали в течение 7-8 недель, а затем пересаживали на поле. Использовали стандартные полевые практики, а орошение использовалось по мере необходимости. Уборка и сбор продукта, обрабатываемого дымовой и воздушной сушкой (как для Берли) было завершено через около 4 с половиной месяца. Дымовая сушка включала: 48 часов колоризация, 85-100°F, относительная влажность 94%; 24 часа завядание, 100°F до -120°F, до 54% относительной влажности; 30 часов сушки листьев, 120°F-135°F, 40% относительной влажности; 40 часов сушки стебля, 135°F - 168°F, до 22% относительной влажности.
Измерение компонентов экссудата как производных TMS с помощью GC-MS
Для измерения зеленых листьев с середины листовой пластинки среднего листа каждого растения вырезали два диска (диаметром 2 см). Диски промывали ацетонитрилом (5 мл) в течение 30 секунд. Смывы концентрировали с помощью вакуумного роторного испарителя с получением маслянистого остатка. Остаток дериватизировали с образованием сложных эфиров триметилсилила (TMS), по существу, как описано Severson et al. 1985 г. (см. выше ссылкой) следующим образом: промывные растворы ацетонитрила выпаривали досуха, растворяли в 1 мл CHCl3, переносили в 1,5 мл ампулы GC и сушили в потоке N2 при 40°C. Дериватизированные образцы растворяли в 1 мл диметилформамида (DMF) и 50 мкл BSTFA [бис (триметилсилил) трифторацетамида] и 24 мкг цембрена в качестве внутреннего стандартного цембрена добавляли в каждый флакон. Образцы дериватизировали при 70°C в течение 45 мин, охлаждали до комнатной температуры и анализировали.
Производные TMS разделяли и анализировали с помощью GC-MS (HP6890 GC, оснащенный HP5973 MS и автоматическим дозатором), используя 30,0 m капиллярную колонку Agilent 19091J-413 с толщиной пленки 0,25 мкм и диаметром 0,25 мм. Гелий использовали в качестве газа-носителя с постоянным расходом 1,8 мл/мин и температурой впрыска 250°C. Программа для работы с производными сложного эфира TMS была следующей: начальная температура 180°C в течение 2,0 мин; скорость 8°C в минуту до 280°C, выдерживать 9,5 мин. Общее время выполнения составило 24,0 мин. Элюированные соединения идентифицировали по времени их удерживания и по сравнению профилей MS со стандартами. Дитерпеноиды элюируются при: от цис-абиенола до βCBT-диола, от 6 до 8 мин; продукты окисления дитерпеноидов - от 8 до 13,5 мин; эфиры сахарозы, от 17 до 22,5 мин. Для измерения тканей, высушенных дымовой и воздушной сушкой, кусочки типичных листьев размером 6,14 × 6,14 см (эквивалент 12 листовых дисков из зеленых тканей) вырезали из сухой ткани и промывали ацетонитрилом. На линию готовили от десяти до шестнадцати повторных образцов и контролировали воздушную и дымовую сушку с применением тех же листьев, которые использовались для анализа зеленых листьев (проколы листьев). Подготовка образцов для химического анализа в качестве производных TMS была такой, как описано выше, и для ацильных групп сложных эфиров сахарозы, как показано ниже.
Были записаны интегрированные площади пиков всех соединений и внутреннего стандарта, и относительное количество каждого из них было рассчитано как % от общего количества. Многочисленные пики типов сложных эфиров сахарозы объединяли для получения суммарной площади пиков.
Среднее количество листьев на растение, зеленая масса листьев на растение и общая масса экссудата на контрольное растение по сравнению с трансгенными линиями представлены в таблице 1 ниже. Данные представляют собой средние значения по 3 растениям каждой линии через 19 дней после прищипывания (прищипывание относится к удалению цветов табака).
Результаты GC-MS для конструкций GW2 и GW5 (T0) и их T1 и T2 поколений были идентичными; обе показали очень высокий уровень лабдендиола и низкий уровень CBT-диолов и цис-абиенола. Чтобы избежать избыточности, полевые работы проводились с применением только конструкции GW2.
Таблица 1
В Таблице 1 показано, что среднее количество листьев на растение, зеленая масса листьев на растение были одинаковыми между тестированными трансгенными линиями и были аналогичны контрольному растению.
Анализ зеленых листьев полевых контрольных и трансгенных линий
Химический анализ экссудата зеленых листьев был проведен сразу после прищипывания и показал, что для каждой линии химический состав экссудата в полевых условиях был аналогичен химическому составу, наблюдаемому для растений в теплице, и что изменчивость между растениями в химическом составе составляла около 25-30% для контроля и каждой линии в полевых условиях (см. Фигуру 3). Каждая точка на графике представляет данные одного растения. Данные, представленные на Фигуре 3, демонстрируют, что:
- Линии GW1 и GW3 обогащены цис-абиенолом
- Линия GW2 обогащена лабден-диолом
Измерение ацильного состава сложного эфира сахарозы
Используемые методы модифицированы из Severson et al. 1985 (см. выше ссылкой). К 5-10 мг образцов камеди в стеклянной пробирке Corex объемом 30 мл добавляют 0,5 мл КОН (1 М) в 80% МеОН и оставляют на ночь при 22°C до омыления. Образцы затем сушили в атмосфере N2 с последующим добавлением 1 мл н-BuOH и 3 капель концентрированной H2SO4. Затем образцы нагревали при 110°C в течение 1 часа, после чего компоненты распределяли между гексаном и водой (по 1,5 мл каждый) с применением импульсного вихревого смесителя (10 импульсов, 1,5 секунды каждый). Было сделано шесть-семь экстракций (пока водная фаза не стала нейтральной к лакмусовой бумаге). Водная фаза содержала K2SO4, H2SO4 и сахара, а гексановая фаза представляла собой бутиловые эфиры ацильных групп. Гексан переносили в 1,5 мл флакон с ГХ и анализировали с помощью GC-MS с применением той же колонки, что и для TMS. Газом-носителем был гелий с постоянным потоком 1,8 мл/мин и температурой впрыска 250°C. Программа для работы с производными бутилового эфира была следующей: начальная температура 90°C в течение 3,0 мин; скорость 3°C в минуту до 160°C, держать в течение 2 минут, скорость 15°C в минуту до 250°C, выдерживать 5 минут. Общее время прогона составило 39,33 мин.
Данные на Фигуре 4 ясно показывают, что линии GW1 и GW3 (которые содержат высокий цис-абиенол) также демонстрируют повышенный выход сложного эфира сахарозы по сравнению с контролем. Каждая точка на графике представляет данные одного растения. Ацильный состав не изменился, поэтому эти линии похожи на турецкий или восточный табак в том, что присутствуют высокие уровни цис-абиенола и высокого предшественника 3-метил валериановой кислоты, но преимущественно они производятся с помощью относительно высокой биомассы растений.
На Фигуре 4 показано, что:
- В линиях с высоким содержанием цис-абиенола (GW1 и GW3) эфиры сахарозы были значительно обогащены по сравнению с контролем.
- В линии GW2 (высокое содержание лабдендиола) содержание сложных эфиров сахарозы также было повышено.
Пример 4. Компоненты экссудата в сыром табаке по сравнению с обработанным дымовой сушкой полевым табаком
Линии RNAi выращивали в поле, Т2 или Т3, признак стабильный. Уборку и сбор продукта, обработанного дымовой и воздушной сушкой (как для Берли) завершили через около 4 с половиной месяца. Дымовая сушка включала: 48 часов колоризация, от 85 до 100°F, относительная влажность 94%; 24 часа завядание, 100°F-120°F, до 54% относительной влажности; 30 часов сушки листьев, 120°F-135°F, 40% относительной влажности; 40 часов сушки стебля, 135°F-168°F, до 22% относительной влажности, как описано в примере 3.
Те же самые хроматограммы TMS, которые были получены для Фигур 3 и 4, использовали для пересчета экссудатного компонента на основе площади поверхности, выраженной в мкг/см2 в Таблицах 2 и 3 (сырой против высушенного дымовой сушкой и сырой против высушенного воздушной сушкой). Расчет проводили на основе сравнения с известным количеством внутреннего стандарта цембрена с предположением, что пиковый отклик идентичен для всех соединений. Количества соединений, показанные в таблице 2, представляют в среднем 20 зеленых растений для контроля и от 34 до 36 зеленых растений и от 10 до 14 высушенных растений для линий GW1-3 ± их соответствующие стандартные отклонения.
Таблица 2. Компоненты экссудата в зеленом табаке по сравнению с табаком, обработанным дымовой сушкой, урожай 2014 года.
*Содержание цис-абиенола в экстракте дисков 2-х листьев было низким. Количества рассчитывали от % суммарного экстракта зеленых листьев.
**Содержание a- и b-CBT-диолов из 2-х дисков зеленых листьев было низким. В данном документе количества рассчитывали от % суммарного экстракта зеленых листьев.
Данные, представленные в таблице 2, показывают, что дымовая сушка не увеличивает компонент экссудата в виде сложного эфира сахарозы в контрольных растениях, но увеличивает содержание сложного эфира сахарозы в трансгенных растениях, включающих конструкции GW1, GW2 или GW3. Дымовая сушка также увеличивала содержание LD в трансгенных растениях, включающих конструкции GW1, GW2 или GW3. Дымовая сушка увеличивала содержание цис-абиенола во всех табачных изделиях.
Данные демонстрируют, что дымовая сушка увеличивает содержание α-CBT-диола в компоненте эксудата трансгенных растений GW2 по сравнению с зеленым табаком. Содержание α-CBT-диола в контрольных растениях также увеличивается при дымовой сушке, но в меньшей степени. Данные также показывают, что дымовая сушка увеличивает содержание β-CBT-диола в компоненте эксудата трансгенных растений GW2 по сравнению с зеленым табаком. Содержание β-CBT-диола в контрольных растениях не зависит от дымовой сушки.
Продукты окисления были увеличены в трансгенных линиях.
Количества соединений, представленные в Таблице 3 (ниже), представляют в среднем 20 зеленых растений для контроля и от 34 до 36 зеленых растений и от 8 до 16 высушенных растений для линий GW1-3 ± их соответствующие стандартные отклонения.
Данные, представленные в Таблице 3 ниже, демонстрируют, что воздушная сушка не увеличивает компонент экссудата в виде сложного эфира сахарозы в контрольных растениях, но немного увеличивает содержание сложного эфира сахарозы в трансгенных растениях, содержащих конструкции GW1, GW2 или GW3. Содержание сложного эфира сахарозы стабильно в трансгенных линиях при воздушной сушке. Данные также демонстрируют, что дымовая сушка увеличивает содержание α-CBT-диола (который является второстепенным компонентом в трансгенных организмах) в экссудатном компоненте трансгенных растений GW2 по сравнению с зеленым табаком. Содержание α-CBT-диола в контрольных растениях не зависит от дымовой сушки.
Данные демонстрируют, что воздушная сушка увеличивает содержание β-CBT-диола в экссудатном компоненте трансгенных растений GW2 по сравнению с зеленым табаком. Содержание β-ХВТ-диола в контрольных растениях также увеличивается при дымовой сушке, но в меньшей степени. Воздушная сушка также увеличивала содержание LD в трансгенных растениях, содержащих конструкции GW1, GW2 или GW3. Воздушная сушка увеличивала содержание цис-абиенола в контроле и табаке, содержащем GW1 или GW3.
Продукты окисления увеличились в трансгенных линиях.
Таблица 3. Экссудатные компоненты в зеленом табаке по сравнению с табаком, обработанным воздушной сушкой, урожай 2014 года
*Содержание цис-абиенола в экстракте 2-х листовых дисков было низким. Количества рассчитывали от % суммарного экстракта зеленых листьев.
**Содержание a- и b-CBT-диолов из 2-х листовых дисков было низким. Количества рассчитывали от % суммарного экстракта зеленых листьев.
Пример 5. Анализы композиции дитерпеноидов и ацильной композиции сложного эфира сахарозы.
Композицию дитерпеноидов и ацильную композицию сложного эфира сахарозы измеряли, как описано в примере 3. Авторы изобретения сравнивали композицию дитерпеноидов и сложного эфира сахарозы в трансгенных линиях с контролем TI 1068 и с коммерческими турецким табаком, высушенным на солнце. Высушенный на солнце коммерческий табак был отправлен авторам изобретения в виде прессованного листа. Данные, представленные для линий трансгенного табака, представляют в среднем от 10 до 14 растений, в то время как контроль TI 1068 представляет в среднем более 16 растений.
Для сравнения были проанализированы следующие коммерческие турецкие типы:
A = Oriental, Поставщик: Socotab EOOD, урожай 2013
B = Турция-Самсун Урожай 2013, класс SMAL
C = Турция-IXMIR, урожай 2013, класс YZAL
См. Таблицу 4 Анализ состава экссудата на содержание дитерпеноидов.
Данные в Таблице 4 (ниже) демонстрируют, что трансгенные линии GW1, GW2 и GW3, обработанные дымовой сушкой, имеют более высокое содержание сложного эфира сахарозы по сравнению с контрольными и/или протестированными растениями Турции.
См. Таблицу 5 Анализ состава экссудата в отношении состава ацильной группы сложного эфира сахарозы.
Данные, представленные для линий трансгенного табака, представляют в среднем от 4 до 8 растений, в то время как контроль TI 1068 представляет в среднем более 10 растений. Данные в таблице 5 (ниже) показывают, что ацильный состав сложного эфира сахарозы трансгенных линий табака GW1, GW2 и GW3 аналогичен составу табака турецкого типа, то есть богат 3-Me валериановыми ацильными группами.
Таблица 4. Анализ состава экссудата на содержание дитерпеноидов
Таблица 5. Анализ состава экссудата в отношении композиции ацильных групп в сложном эфире сахарозы
Пример 6. Анализ химического состава поверхности GW3.
Были проведены анализы зеленого полевого, обработанного воздушной сушкой и обработанного дымовой сушкой образцов. Там, где это возможно, все было сделано на образцах из одних и тех же листьев путем маркировки и нумерации листьев в поле, когда были взяты листовые проколы для получения данных о зеленом полевом материале. После воздушной или дымовой сушки отбирали одинаковые пронумерованные листья, чтобы можно было непосредственно сравнивать образцы зеленый/воздушный и зеленый/дымовой. В приведенной ниже таблице показаны результаты зеленых образцов по сравнению с обработанными дымовой сушкой и зеленых по сравнению с обработанными воздушной сушкой. Данные были стандартизированы до микрограмма/см2 листа.
Таблица 6. Анализ химического состава поверхности GW3 зеленых полевых, обработанных воздушной сушкой и обработанных дымовой сушкой образцов
Компоненты эксудата: Полевые табаки - Зеленый против высушенного дымом- против высушенного воздухом
Числа в парантезах после «зеленый», «обработанный дымовой сушкой» или «обработанный воздушной сушкой» указывают, сколько независимых образцов было проанализировано и использовано для получения среднего значения. Например, для «контрольный зеленый (20)» означает 20 независимых образцов. Средние значения для них показаны со стандартным отклонением ниже, например, для контрольного зеленого (2), 6,695 ± 3,834.
Данные для «Зеленый» по отношению к «Обработанному дымовой сушкой» предназначены для линий: контроль и GW3 - высокое содержание цис-абиенола. Эти данные демонстрируют, что содержание цис-абиенола значительно уменьшалось при изменении состояния от зеленого до обработанного дымовой сушкой, и это цис-соединение, возможно, было разложено до определенных «продуктов окисления» в правой колонке для линии GW3 с высоким содержанием цис-абиенола. Эти данные также демонстрируют, что в GW3 содержание l-абдендиола увеличивалось при изменении состояния от зеленого до обработанного дымовой сушкой. Изменения были пропорциональными, например, когда цис-форма уменьшается (мкг/см2) при изменении состояния от зеленого до обработанного дымовой сушкой, лабдендиол увеличился пропорционально. Обратите внимание, что значения колонки со значениями продуктов окисления также значительно увеличились для GW3, при изменении состояния от зеленого до обработанного дымовой сушкой. Кроме того, CBT-диолы не сильно изменились при изменении состояния от зеленого до обработанного дымовой сушкой, а количества сложного эфира сахарозы не сильно изменились при изменении состояния от зеленого до обработанного дымовой сушкой в контроле; тогда как были значительно увеличены (+4,7) в линиях GW3. Отдельный анализ (не показан) продемонстрировал отсутствие качественного изменения состава ацильных групп при изменении состояния от зеленого до обработанного дымовой сушкой, как ожидалось для стабильного эфира сахарозы. Данные для изменения состояния от зеленого до обработанного воздушной сушкой показывают, что изменения от зеленого до обработанного воздушной сушкой меньше, чем наблюдаемые для изменения от зеленого до обработанного дымовой сушкой. Только окисленная форма, по-видимому, существенно увеличивается, вероятно, из-за разложения цис-абиенола.
Фигура 5 демонстрирует, что содержание сложного эфира сахарозы было обогащено в трансгенной линии GW-3 по сравнению с контролем. Содержание сложного эфира сахарозы представлено в процентах от общего количества основных экссудатов. Фигура 6 демонстрирует, что содержание цис-абиенола было обогащено в трансгенной линии GW-3 по сравнению с контролем. Содержание цис-абиенола представлено как процент от общего количества основных соединений экссудата.
Пример 7. Экссудатные компоненты полевых табаков урожая 2016, высушенных дымом и высушенных на воздухе
Линии RNAi выращивали в поле, Т2 или Т3, признак стабильный, и собирали. Растения, высушенные дымом, собирали за 3 ломки в 2016 году. В испытании 2016 года растения, высушенные на воздухе, перед анализом срезали с куском стебля, высушивали на воздухе и разделяли на 1/3 нижних листьев, 1/3 средних листьев и 1/3 верхних листьев. Это должно было имитировать общий подход, используемый для коммерческого производства.
Таблица 7. Компоненты экссудата в полевых табаках урожая 2016 года, высушенных дымовой сушкой
а) В 2016 году, в отличие от 2014 года, для дымовой сушки были взяты и проанализированы 3 ломки.
b) Каждое значение каждого праймирования составляет в среднем 3 повтора. Каждый повтор в среднем состоял из 20 растений.
с) В 2016 году, в отличие от 2014 года, зеленая ткань полевых растений не отслеживалась.
Заключение
Данные в Таблице 7 демонстрируют, что сложные эфиры сахарозы были обогащены в линиях GW1-3 по сравнению с контролем в полевом табаке урожая 2016 года, высушенном дымовой сушкой.
Таблица 8. Компоненты экссудата в полевых табаках урожая 2016, высушенных на воздушной сушкой.
a) В 2016 году, в отличие от 2014 года, верхние 1/3, средние 1/3 и нижние 1/3 листьев срезанного стебля, высушенных на воздухе растений были объединены и проанализированы отдельно.
b) Каждое значение каждого уровня в среднем составляет 20 листьев от 20 независимых растений.
c) В 2016 году, в отличие от 2014 года, зеленая ткань полевых растений не контролировалась.
Заключение
Данные в Таблице 8 демонстрируют, что сложные эфиры сахарозы были обогащены в линиях GW1-3 по сравнению с контролем в полевом табаке урожая 2016 года, обработанного воздушной сушкой.
Все публикации, упомянутые в приведенном выше описании, включены в него ссылкой. Различные модификации и вариации описанных способов и систем согласно настоящему изобретению будут очевидны для специалистов в данной области техники без отступления от объема и сущности настоящего изобретения. Хотя настоящее изобретение было описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что заявленное изобретение не должно чрезмерно ограничиваться такими конкретными вариантами осуществления. Действительно, различные модификации описанных способов осуществления изобретения, которые очевидны для специалистов в области биохимии и биотехнологии или в смежных областях, должны находиться в пределах объема следующей формулы изобретения.
--->
Список последовательностей
<110> University of Kentucky Research Foundation
<120> METHOD
<130> P108191RU
<140> EP 18706918.2
<141> 2018-02-06
<150> US 62/455850
<151> 2017-02-07
<160> 11
<170> PatentIn version 3.5
<210> 1
<211> 4624
<212> DNA
<213> Nicotiana tabacum
<400> 1
ccttatattt atccaccact tgagctactt tctataacca attaaagtaa gtccaattct 60
aacattgtat gctgtgctgc ccttattttt ggctacaaaa ctcgaaagca aaggaactag 120
aaaactcgtc tggcgagaga aagagagatg agtcaatcaa tttctccaat ctttcctcgc 180
tttgcaaaat ttcagtcgaa tatttggagg tgcagtactt ttgaactcag agttatacac 240
tcatcatatg cctctattgg agggaggaga aaagagagag aaagaagaat gaagcgagca 300
atgaatcctt cttcaagctc gcgtcatttg gcagattttc actcaaccat ttggggtgac 360
cattttctct cctacaattc tgaaataaca gtaggtcaca tacatatgta atcacatgct 420
tatattctat ttgaatttgt tatctaaatg tttaaaggaa taaagatgtt ataattttat 480
tagagacagg atcaagcatt taaacttgga aggtttaacc tttaagattg gtctttatcg 540
tacttttgaa attatgagtt tgaaatttaa tactccatcc gtctcaataa agaatgaata 600
ttttactatc tagggagtca aacaagattt tctcaacctt ttttttcgca aatgcatttt 660
aaaaattttg aaatttttaa ttgttgtgac ttacaactac cttcttatgt acttcctaaa 720
tgtgtaaatc tcattttcaa aaaatttacg gaatatatat tcgtcacatt gaaaatattt 780
aatttgaccc tcatactccg aaaaggttca aataaattga aacgaatgga atagtactat 840
tttgtaaaaa cttatgtaga ttttcactat atatctaata agtattcaaa actaataaat 900
aatgatcaac atatgcggat ctaagattta aattttttgg gttcaccttt aaggatctgt 960
tacaatttta gtagaatgtt accataaatt tgtgctccgt gaaatgtatt gagtcagatg 1020
aacctggtat tatacatgcg gatacgctcc tgatgctcaa tttctgcctg caggaaatta 1080
ctacccaaga gaaaaatgaa catgaaatgc taaaagaaat agttcggaaa atgttggtag 1140
aaactccaga taatagtaca caaaaactag tcttgattga cacaattcaa agattgggat 1200
tagcatatca tttcaatgat gagattgaaa actccattca aaacatcttt aatttgtctc 1260
aaaatagtga aaatgacaat gaacacaacc tttatgttgc tgctcttcgt tttcgacttg 1320
cgaggcaaca aggatattac atgtcttcag gtaccttaca tttctgccct ttcccgcaca 1380
gcttcatttt ttttcgttgt taaaggcagc tcggcgtata aaatatctcg tgtatacgca 1440
gggtcaggac ggaaccgccc ccaaggggtg taaagtatgc aacctaccct aatactaaat 1500
atctcgtgtt atacacaggg tcaggacaag tcgcacccaa gaggtgtaat gtaggcaact 1560
taccctaatg ctagcattag taactgattt tatggctcaa acacataaat tgtaggtcac 1620
acagtaacaa ctttatcgtt ctcaaagact cgccttcctc tttttttagt tatcgcacct 1680
tatttgttgc aaagaatagc aagtttcgag atctgcttct atataaaaaa cttctgtatt 1740
atactttttt attttgtcct tctgcttaaa aatagtaaaa aactataatg tggaaattgt 1800
aaatttctta actagctgtg aaatcaaata gttattatag gaatataatt tagactccac 1860
ttatggaaaa ccactgggtt gccgttgtta ttgtcaataa taacttgggg tacgatttac 1920
ttctttttcc atggctgtcc acgactatat ttctattaac caatgttgtg actatgcttt 1980
cccttgagtc gaggatctat tgataacagg ctcttcgatc tttaacaagg taaaagtaat 2040
gtctgcgtac acactctact ccgcagaccc acttgtagga tttcattgaa tttttttttt 2100
tgttgttgtt gttgtaataa cttagggttt agtttcttga tgctgatgaa attcagttct 2160
ttcaactata aacatggtgt tcaccagatg tgttcaagca attcactaac catgacggaa 2220
aattcaagga aaatcatatt aatgatattc aaggattact aagtttgtat gaagcaacac 2280
atatgagagt gcacgacgag gaaattctag aagaagctct tatctttacc accactcatc 2340
tcgagtccat gatcccaaat ttgagcaact cgcttaaggt acaagttact gaagcctcaa 2400
accaacctat tcgcaaaact ataccgaggg tgggatcaag gaaatacata tacatatatg 2460
aaaacattgg aacacataat gatttgcttg tgaaatttgc aaagttggac tttaacatgt 2520
tacaagagct tcatcgaaaa gagctcaacg agctaacaag gtacatctac tattcttatc 2580
attttcatag ttatggtaca gtcagatctc tctataaaat ccatccttta taacaacggt 2640
tcaccataac ggtcatgttt tctttagaac taatctttta tgttaccaaa aaatttcgaa 2700
acaattgaga ctattataga gatgtttgat ggtaactcgc gctaattaat aacacctaaa 2760
gtttaagtat gttaatgttg ttgtgttatc tatagctggt ggaaagaaat ggattttgca 2820
acaaatttcc aatatgcaaa gggcagattg gtagaagctt acttttggat ggtgggaata 2880
tattttgaac ctcaatatag tcgttcaaga agaatgataa cacaagtagt caacatgaac 2940
tccatcattg atgacactta tgatgctttt gcaacttttg atgagcttat gcttttcacg 3000
gatgcgatcc aaaggtaatc tttctataac aactgcattt gttctgataa ttttttaaga 3060
tgctatttga agtgttgtta tagagaaata tattatgaca acttagactt tgcagatggg 3120
atgtaggtgc catggattca ttaccggcat atttgagacc tatttatcaa ggccttctcg 3180
acgttttcaa tgaaatggaa gaagtaatgg ccaaagaagg taaagcagat cgcatctact 3240
atgcgaaaaa agaggtaatc cttgattaag ttacattaat tactacttaa taagttaatt 3300
aagtaaacca agttgtaggg aagaatcaca attttgaact attagtactt tttctgttac 3360
ttttttagat gaaaaagttg gtggcagcct attttaagga agttgaatgg ttgaatgcta 3420
actacattcc aaaatgtgag gagtatatga aaaatggagt tgtaagttgt accggtagat 3480
gtatggaaca atttgctttg gttgttatcg aggaaattat aacaaaagag gcttttgaat 3540
ggttggcaaa tgaacctttg attcctcgag ctgcatcaac aatctgtaga ttaatggatg 3600
atattgttga tcatgaatta agtataacaa tataatttcc attttatata acaattagtc 3660
atcctaattc acaaattttg tccctaaata catacaaaaa caactacaat aacagaaaca 3720
acatatccag tgtattccta tagtacgggt ctgggcagag agatgtgtat gaagatctta 3780
ccctatcttg tggaggtaga aaggttgttt cccgatagac cctcgactca aaaaaacatt 3840
tctcaatctg atttcgagtc taggtggcac ttttgcatga tataataaat agacatgctt 3900
gataaattac aacttcaatg agcacattta cataaagtga tttatggaac tttagaaacg 3960
aactgattaa aatggtaaaa tattgtataa tattaatgaa gatattgaaa tatattatgt 4020
gtaggttgaa caacaaagag gacatgttgc ttcatttgtt gagtactaca tgaaagaata 4080
tggaacttca aagcaagaag catatgttga gatgtggaaa aagatcacaa atgcgtggaa 4140
agacataaat aaggaactcc tgcgcgctac tgcagtacca atgtttgtcc tcgaacgaac 4200
tttagattat acaagattgg ttgatacatg tttcaaagat gatgatggtt acacaaatcc 4260
caaatccaaa gtgaaagaca tgattgcttt gttgtttgtc gaatctatcg acatatgatg 4320
atatataaca atgcagatgc accttcaacc gagtattcag agcaaatatg gaagcatttt 4380
gtatggttct gtatgaccta taggtcatat gttcgagtca tggaagcatc cattaatact 4440
tgcattaggg taggctgtct atatcatact tattagggtg cgacccttcc cctgactctg 4500
catgaatgca ggatgctttg tgcgctcagc tgcctttttt tactatttcg ctgtcagtta 4560
tgtttgagaa gggacaatac ttgatttgat tcatgcagtc ttgtctccag gtttgattcc 4620
tatg 4624
<210> 2
<211> 597
<212> PRT
<213> Nicotiana tabacum
<400> 2
Met Ser Gln Ser Ile Ser Pro Ile Phe Pro Arg Phe Ala Lys Phe Gln
1 5 10 15
Ser Asn Ile Trp Arg Cys Ser Thr Phe Glu Leu Arg Val Ile His Ser
20 25 30
Ser Tyr Ala Ser Ile Gly Gly Arg Arg Lys Glu Arg Glu Arg Arg Met
35 40 45
Lys Arg Ala Met Asn Pro Ser Ser Ser Ser Arg His Leu Ala Asp Phe
50 55 60
His Ser Thr Ile Trp Gly Asp His Phe Leu Ser Tyr Asn Ser Glu Ile
65 70 75 80
Thr Glu Ile Thr Thr Gln Glu Lys Asn Glu His Glu Met Leu Lys Glu
85 90 95
Ile Val Arg Lys Met Leu Val Glu Thr Pro Asp Asn Ser Thr Gln Lys
100 105 110
Leu Val Leu Ile Asp Thr Ile Gln Arg Leu Gly Leu Ala Tyr His Phe
115 120 125
Asn Asp Glu Ile Glu Asn Ser Ile Gln Asn Ile Phe Asn Leu Ser Gln
130 135 140
Asn Ser Glu Asn Asp Asn Glu His Asn Leu Tyr Val Ala Ala Leu Arg
145 150 155 160
Phe Arg Leu Ala Arg Gln Gln Gly Tyr Tyr Met Ser Ser Asp Val Phe
165 170 175
Lys Gln Phe Thr Asn His Asp Gly Lys Phe Lys Glu Asn His Ile Asn
180 185 190
Asp Ile Gln Gly Leu Leu Ser Leu Tyr Glu Ala Thr His Met Arg Val
195 200 205
His Asp Glu Glu Ile Leu Glu Glu Ala Leu Ile Phe Thr Thr Thr His
210 215 220
Leu Glu Ser Met Ile Pro Asn Leu Ser Asn Ser Leu Lys Val Gln Val
225 230 235 240
Thr Glu Ala Ser Asn Gln Pro Ile Arg Lys Thr Ile Pro Arg Val Gly
245 250 255
Ser Arg Lys Tyr Ile Tyr Ile Tyr Glu Asn Ile Gly Thr His Asn Asp
260 265 270
Leu Leu Val Lys Phe Ala Lys Leu Asp Phe Asn Met Leu Gln Glu Leu
275 280 285
His Arg Lys Glu Leu Asn Glu Leu Thr Ser Trp Trp Lys Glu Met Asp
290 295 300
Phe Ala Thr Asn Phe Gln Tyr Ala Lys Gly Arg Leu Val Glu Ala Tyr
305 310 315 320
Phe Trp Met Val Gly Ile Tyr Phe Glu Pro Gln Tyr Ser Arg Ser Arg
325 330 335
Arg Met Ile Thr Gln Val Val Asn Met Asn Ser Ile Ile Asp Asp Thr
340 345 350
Tyr Asp Ala Phe Ala Thr Phe Asp Glu Leu Met Leu Phe Thr Asp Ala
355 360 365
Ile Gln Arg Trp Asp Val Gly Ala Met Asp Ser Leu Pro Ala Tyr Leu
370 375 380
Arg Pro Ile Tyr Gln Gly Leu Leu Asp Val Phe Asn Glu Met Glu Glu
385 390 395 400
Val Met Ala Lys Glu Gly Lys Ala Asp Arg Ile Tyr Tyr Ala Lys Lys
405 410 415
Glu Met Lys Lys Leu Val Ala Ala Tyr Phe Lys Glu Val Glu Trp Leu
420 425 430
Asn Ala Asn Tyr Ile Pro Lys Cys Glu Glu Tyr Met Lys Asn Gly Val
435 440 445
Val Ser Cys Thr Gly Arg Cys Met Glu Gln Phe Ala Leu Val Val Ile
450 455 460
Glu Glu Ile Ile Thr Lys Glu Ala Phe Glu Trp Leu Ala Asn Glu Pro
465 470 475 480
Leu Ile Pro Arg Ala Ala Ser Thr Ile Cys Arg Leu Met Asp Asp Ile
485 490 495
Val Asp His Glu Val Glu Gln Gln Arg Gly His Val Ala Ser Phe Val
500 505 510
Glu Tyr Tyr Met Lys Glu Tyr Gly Thr Ser Lys Gln Glu Ala Tyr Val
515 520 525
Glu Met Trp Lys Lys Ile Thr Asn Ala Trp Lys Asp Ile Asn Lys Glu
530 535 540
Leu Leu Arg Ala Thr Ala Val Pro Met Phe Val Leu Glu Arg Thr Leu
545 550 555 560
Asp Tyr Thr Arg Leu Val Asp Thr Cys Phe Lys Asp Asp Asp Gly Tyr
565 570 575
Thr Asn Pro Lys Ser Lys Val Lys Asp Met Ile Ala Leu Leu Phe Val
580 585 590
Glu Ser Ile Asp Ile
595
<210> 3
<211> 2433
<212> DNA
<213> Nicotiana tabacum
<400> 3
tgcagccaca gtactgcttc atcaatggaa gaggcaaagg agagaataag ggaaacattt 60
ggaaaaatag agctatctcc ttcttcctat gacacagcat gggtagctat ggtcccttca 120
agatattcta tgaaccaacc atgttttcct cagtgcttag attggattct tgaaaatcaa 180
agagaagatg gatcttgggg cctaaatcct agccatccat tgcttgtaaa agactccctt 240
tcttccactc tagcatcttt gcttgccctt cgcaaatgga gaattggaga taaccaagtc 300
caaagaggcc ttggctttat tgaaacgcat ggttgggcag tcgataacaa ggatcagatt 360
tcacctttag gatttgaaat tatatttccc tgcatgacca actatgcaga gaaacttaat 420
ttggatctac ctttggatcc taaccttgta aatatgatgc tctgcgaacg tgaattaaca 480
attgaaagag ccttaaagaa tgaattcgag gggaatatgg caaatgtaga atattttgct 540
gaaggactcg gtgaatcatg tcattggaaa gagatgatgc ttcgtcagag acacaacggg 600
tcgctctttg attcaccagc cactactgca gctgccttga tttaccatca gtacgatgag 660
aaatgctttg ggtacttgaa ctcaatcttg aaactgcacg ataattgggt ccacactatt 720
tgccctacaa agatacattc aaatctcttc ttagttgatg cccttcaaaa tcttggagta 780
gatcggtatt ttaaaacaga agtcaaaaga gtactagatg aaatatacag gctttggcta 840
gaaaagaatg aagaaatttt ttcagacgtt gctcattgtg ccatggcgtt tcgactttta 900
cggatgaata actatgaagt ttcctcagaa gaacttgaag gatttgtcga ccaagaacat 960
ttctttacaa catcaagtgg gaaacttatg aatcacgttg caattctcga acttcaccga 1020
gcttcacagg tggctattca tgaaaggaaa gatcacattt tagataaaat aagtacttgg 1080
acaaggaatt ttatggagca aaaactcttg gacaagcaca tccctgatag gtcaaagaag 1140
gagatggaat ttgctatgag gaaattttat ggcacatttg atcgagtgga aactagacgt 1200
tacatcgagt catacaaaat ggacagtttt aagatcttaa aagcggctta caggtcttcc 1260
ggtattaaca acatagactt gctaaagttc tcagaacacg attttaactt gtgccaaacc 1320
cgacacaaag aagaacttca acagatgaaa aggtggttca cagattgcaa actcgaacaa 1380
gtaggattat cacaacagta cttatacact agttacttca taattgctgc tatactcttt 1440
gaacctgaat atgctgatgc tcgtctagca tatgcaaagt acgccataat aataacagcg 1500
gtggatgatt tcttcgattg ttttatttgc aaagaagaac tgcaaaacat catcgaatta 1560
gtagagagat gggagggata ctcaaccgtc ggattccgtt cagagagggt tagaattttc 1620
tttttggcac tttacaaaat ggtagaggaa attgcggcaa aggcggaaac taagcaaggt 1680
cgatgtgtca aagatcacct tattaacttg tggattgata tgttgaagtg tatgctagtg 1740
gaattggacc tttggaaaat taaatcaact accccaagca tagaggagta cttgtctgtt 1800
gcatgtgtaa ctattggtgt tccatgtttt gttctcacat cactatatct tcttggacca 1860
aaactgtcca aggatgtcat agaaagttct gaggtcagtg ccttatgcaa ttgtacagct 1920
gctgtggccc gattgattaa tgatatacac agttacaaga gagaacaagc agaaagttca 1980
acaaatatgg tatcaatatt aataacacaa agtcagggaa ctatctctga agaagaggct 2040
ataagacaga taaaggaaat gatggaaagt aagagaagag agttgctagg gatggttcta 2100
caaaataaag aaagccaatt gccacaagtg tgcaaggatc ttttttggac gacaatcaac 2160
gcagcagctt attctataca tacacatggg cgatgggtat cgcttcccag aggaattcaa 2220
gaaccatatc aacgatgtaa tttacaaacc actcaatcaa tattccccat aatatgcctt 2280
aaatctttta caatatgtta ctaatctttg gaacttggtt gtgatattat tagatgcatg 2340
gacgaattgt acttctttta tgttgtgcac aataatgtac aactgttact atgggaaaaa 2400
cttacttaca ctgctaaaaa aaaaaaaaaa aaa 2433
<210> 4
<211> 759
<212> PRT
<213> Nicotiana tabacum
<400> 4
Met Glu Glu Ala Lys Glu Arg Ile Arg Glu Thr Phe Gly Lys Ile Glu
1 5 10 15
Leu Ser Pro Ser Ser Tyr Asp Thr Ala Trp Val Ala Met Val Pro Ser
20 25 30
Arg Tyr Ser Met Asn Gln Pro Cys Phe Pro Gln Cys Leu Asp Trp Ile
35 40 45
Leu Glu Asn Gln Arg Glu Asp Gly Ser Trp Gly Leu Asn Pro Ser His
50 55 60
Pro Leu Leu Val Lys Asp Ser Leu Ser Ser Thr Leu Ala Ser Leu Leu
65 70 75 80
Ala Leu Arg Lys Trp Arg Ile Gly Asp Asn Gln Val Gln Arg Gly Leu
85 90 95
Gly Phe Ile Glu Thr His Gly Trp Ala Val Asp Asn Lys Asp Gln Ile
100 105 110
Ser Pro Leu Gly Phe Glu Ile Ile Phe Pro Cys Met Thr Asn Tyr Ala
115 120 125
Glu Lys Leu Asn Leu Asp Leu Pro Leu Asp Pro Asn Leu Val Asn Met
130 135 140
Met Leu Cys Glu Arg Glu Leu Thr Ile Glu Arg Ala Leu Lys Asn Glu
145 150 155 160
Phe Glu Gly Asn Met Ala Asn Val Glu Tyr Phe Ala Glu Gly Leu Gly
165 170 175
Glu Ser Cys His Trp Lys Glu Met Met Leu Arg Gln Arg His Asn Gly
180 185 190
Ser Leu Phe Asp Ser Pro Ala Thr Thr Ala Ala Ala Leu Ile Tyr His
195 200 205
Gln Tyr Asp Glu Lys Cys Phe Gly Tyr Leu Asn Ser Ile Leu Lys Leu
210 215 220
His Asp Asn Trp Val His Thr Ile Cys Pro Thr Lys Ile His Ser Asn
225 230 235 240
Leu Phe Leu Val Asp Ala Leu Gln Asn Leu Gly Val Asp Arg Tyr Phe
245 250 255
Lys Thr Glu Val Lys Arg Val Leu Asp Glu Ile Tyr Arg Leu Trp Leu
260 265 270
Glu Lys Asn Glu Glu Ile Phe Ser Asp Val Ala His Cys Ala Met Ala
275 280 285
Phe Arg Leu Leu Arg Met Asn Asn Tyr Glu Val Ser Ser Glu Glu Leu
290 295 300
Glu Gly Phe Val Asp Gln Glu His Phe Phe Thr Thr Ser Ser Gly Lys
305 310 315 320
Leu Met Asn His Val Ala Ile Leu Glu Leu His Arg Ala Ser Gln Val
325 330 335
Ala Ile His Glu Arg Lys Asp His Ile Leu Asp Lys Ile Ser Thr Trp
340 345 350
Thr Arg Asn Phe Met Glu Gln Lys Leu Leu Asp Lys His Ile Pro Asp
355 360 365
Arg Ser Lys Lys Glu Met Glu Phe Ala Met Arg Lys Phe Tyr Gly Thr
370 375 380
Phe Asp Arg Val Glu Thr Arg Arg Tyr Ile Glu Ser Tyr Lys Met Asp
385 390 395 400
Ser Phe Lys Ile Leu Lys Ala Ala Tyr Arg Ser Ser Gly Ile Asn Asn
405 410 415
Ile Asp Leu Leu Lys Phe Ser Glu His Asp Phe Asn Leu Cys Gln Thr
420 425 430
Arg His Lys Glu Glu Leu Gln Gln Met Lys Arg Trp Phe Thr Asp Cys
435 440 445
Lys Leu Glu Gln Val Gly Leu Ser Gln Gln Tyr Leu Tyr Thr Ser Tyr
450 455 460
Phe Ile Ile Ala Ala Ile Leu Phe Glu Pro Glu Tyr Ala Asp Ala Arg
465 470 475 480
Leu Ala Tyr Ala Lys Tyr Ala Ile Ile Ile Thr Ala Val Asp Asp Phe
485 490 495
Phe Asp Cys Phe Ile Cys Lys Glu Glu Leu Gln Asn Ile Ile Glu Leu
500 505 510
Val Glu Arg Trp Glu Gly Tyr Ser Thr Val Gly Phe Arg Ser Glu Arg
515 520 525
Val Arg Ile Phe Phe Leu Ala Leu Tyr Lys Met Val Glu Glu Ile Ala
530 535 540
Ala Lys Ala Glu Thr Lys Gln Gly Arg Cys Val Lys Asp His Leu Ile
545 550 555 560
Asn Leu Trp Ile Asp Met Leu Lys Cys Met Leu Val Glu Leu Asp Leu
565 570 575
Trp Lys Ile Lys Ser Thr Thr Pro Ser Ile Glu Glu Tyr Leu Ser Val
580 585 590
Ala Cys Val Thr Ile Gly Val Pro Cys Phe Val Leu Thr Ser Leu Tyr
595 600 605
Leu Leu Gly Pro Lys Leu Ser Lys Asp Val Ile Glu Ser Ser Glu Val
610 615 620
Ser Ala Leu Cys Asn Cys Thr Ala Ala Val Ala Arg Leu Ile Asn Asp
625 630 635 640
Ile His Ser Tyr Lys Arg Glu Gln Ala Glu Ser Ser Thr Asn Met Val
645 650 655
Ser Ile Leu Ile Thr Gln Ser Gln Gly Thr Ile Ser Glu Glu Glu Ala
660 665 670
Ile Arg Gln Ile Lys Glu Met Met Glu Ser Lys Arg Arg Glu Leu Leu
675 680 685
Gly Met Val Leu Gln Asn Lys Glu Ser Gln Leu Pro Gln Val Cys Lys
690 695 700
Asp Leu Phe Trp Thr Thr Ile Asn Ala Ala Ala Tyr Ser Ile His Thr
705 710 715 720
His Gly Arg Trp Val Ser Leu Pro Arg Gly Ile Gln Glu Pro Tyr Gln
725 730 735
Arg Cys Asn Leu Gln Thr Thr Gln Ser Ile Phe Pro Ile Ile Cys Leu
740 745 750
Lys Ser Phe Thr Ile Cys Tyr
755
<210> 5
<211> 2352
<212> DNA
<213> Artificial Sequence
<220>
<223> ddRNAi DNA construct (GW1) insert sequence
<400> 5
ctcgtttggc gagagaaaga gagagatgag tcaatcaatt tctccattaa tgttttctca 60
ctttgcaaaa tttcagtcga atatttggag atgcaatact tctcaactca gagttataca 120
ctcatcatat gcctcttttg gagggagaag aaaagagaga gtaagaagaa tgaatcgagc 180
aatggatctt tcttcaagct ctcgtcattt ggcagatttt ccctcaacaa tttggggtga 240
ccattttctc tcctacaatt ctgaaataac agaaattact acccaagaga aaaatgaaca 300
tgaaatgcta aaagaaatag ttcggaaaat gttggtagaa actccagata atagtacaca 360
aaaactagtc ttgattgaca caattcaaag attgggatta gcatatcatt tcaatgatga 420
gattgaaaac tccattcaaa acatctttaa tttgtctcaa aatagtgaag atgacgatga 480
acacaacctt tatgttgctg ctcttcgttt tcgacttgcg aggcaacaag gatattacat 540
gtcttcagat gtgttcaagc aattcactaa ccatgacgga aaattcaagg aaaatcatac 600
taatgatgtt caaggattat tgagtttgta tgaagcagca catatgagag tgcacgacga 660
ggagatatct acccgcttcg cgtcggcatc cggtcagtgg cagtgaaggg cgaacagttc 720
ctgattaacc acaaaccgtt ctactttact ggctttggtc gtcatgaaga tgcggacttg 780
cgtggcaaag gattcgataa cgtgctgatg gtgcacgacc acgcattaat ggactggatt 840
ggggccaact cctaccgtac ctcgcattac ccttacgctg aagagatgct cgactgggca 900
gatgaacatg gcatcgtggt gattgatgaa actgctgctg tcggctttaa cctctcttta 960
ggcattggtt tcgaagcggg caacaagccg aaagaactgt acagcgaaga ggcagtcaac 1020
ggggaaactc agcaagcgca cttacaggcg attaaagagc tgatagcgcg tgacaaaaac 1080
cacccaagcg tggtgatgtg gagtattgcc aacgaaccgg atacccgtcc gcaaggtgca 1140
cgggaatatt tcgcgccact ggcggaggca acgcgtaaac tcgacccgac gcgtccgatc 1200
acctgcgtca atgtaatgtt ctgcgacgct cacaccgata ccatcagcga tctctttgat 1260
gtgctgtgcc tgaaccgtta ttacggatgg tatgtccaaa gcggcgattt ggaaacggca 1320
gagaaggtac tggaaaaaga acttctggcc tggcaggaga aactgcatca gccgattatc 1380
atcaccgaat acggcgtgga tacgttagcc gggctgcact caatgtacac cgacatgtgg 1440
agtgaagagt atcagtgtgc atggctggat atgtatcacc gcgtctttga tcgcgtcagc 1500
gccgtcgtcg gtgaacaggt atggaatttc gccgattttg cgacctcgca aggcatattg 1560
cgcgttggcg gtaacaagaa agggatcttc actcgcgacc gcaaaccgaa gtcggcggct 1620
tttctgctgc aaaaacgctg gactggcatg aacttcggtg aaaaaccgca gcagggaggc 1680
aaacaatgat cctcgtcgtg cactctcata tgtgctgctt catacaaact caataatcct 1740
tgaacatcat tagtatgatt ttccttgaat tttccgtcat ggttagtgaa ttgcttgaac 1800
acatctgaag acatgtaata tccttgttgc ctcgcaagtc gaaaacgaag agcagcaaca 1860
taaaggttgt gttcatcgtc atcttcacta ttttgagaca aattaaagat gttttgaatg 1920
gagttttcaa tctcatcatt gaaatgatat gctaatccca atctttgaat tgtgtcaatc 1980
aagactagtt tttgtgtact attatctgga gtttctacca acattttccg aactatttct 2040
tttagcattt catgttcatt tttctcttgg gtagtaattt ctgttatttc agaattgtag 2100
gagagaaaat ggtcacccca aattgttgag ggaaaatctg ccaaatgacg agagcttgaa 2160
gaaagatcca ttgctcgatt cattcttctt actctctctt ttcttctccc tccaaaagag 2220
gcatatgatg agtgtataac tctgagttga gaagtattgc atctccaaat attcgactga 2280
aattttgcaa agtgagaaaa cattaatgga gaaattgatt gactcatctc tctctttctc 2340
tcgccaaacg ag 2352
<210> 6
<211> 3675
<212> DNA
<213> Artificial Sequence
<220>
<223> ddRNAi DNA construct (GW3) insert sequence
<400> 6
gaagcttact tttggatggt gggaatatat tttgaacctc aatatagtcg ttcaagaaga 60
atgataacac aagtagtcaa catgaactcc atcattgatg acacttatga tgcttttgca 120
acttttgatg agcttatgct tttcacggat gcgatccaaa ggtaatcttt ctataacaac 180
tgcatttgtt ctgataattt tttaagatgc tatttgaagt gttgttatag agaaatatat 240
tatgacaact tagactttgc agatgggatg taggtgccat ggattcatta ccggcatatt 300
tgagacctat ttatcaaggc cttctcgacg ttttcaatga aatggaagaa gtaatggcca 360
aagaaggtaa agcagatcgc atctactatg cgaaaaaaga ggtaatcctt gattaagtta 420
cattaattac tacttaataa gttaattaag taaaccaagt tgtagggaag aatcacaatt 480
ttgaactatt agtacttttt ctgttacttt tttagatgaa aaagttggtg gcagcctatt 540
ttaaggaagt tgaatggttg aatgctaact acattccaaa atgtgaggag tatatgaaaa 600
atggagttgt aagttgtacc ggtagatgta tggaacaatt tgctttggtt gttatcgagg 660
aaattataac aaaagaggct tttgaatggt tggcaaatga acctttgatt cctcgagctg 720
catcaacaat ctgtagatta atggatgata ttgttgatca tgaattaagt ataacaatat 780
aatttccatt ttatataaca attagtcatc ctaattcaca aattttgtcc ctaaatacat 840
acaaaaacaa ctacaataac agaaacaaca tatccagtgt attcctatag tacgggtctg 900
ggcagagaga tgtgtatgaa gatcttaccc tatcttgtgg aggtagaaag gttgtttccc 960
gatagaccct cgactcaaaa aaacatttct caatctgatt tcgagtctag gtggcacttt 1020
tgcatgatat aataaataga catgcttgat aaattacaac ttcaatgagc acatttacat 1080
aaagtgattt atggaacttt agaaacgaac tgattaaaat ggtaaaatat tgtataatat 1140
taatgaagat attgaaatat attatgtgta ggttgaacaa caaagaggac atgttgcttc 1200
atttgttgag tactacatga aagaatatgg aacttcaaag caagaagcat atgttgagat 1260
gtggaaaaag atcacaaatg cgtggaaaga cataaataag gaactcctgc gcgctactgc 1320
agtgatatct acccgcttcg cgtcggcatc cggtcagtgg cagtgaaggg cgaacagttc 1380
ctgattaacc acaaaccgtt ctactttact ggctttggtc gtcatgaaga tgcggacttg 1440
cgtggcaaag gattcgataa cgtgctgatg gtgcacgacc acgcattaat ggactggatt 1500
ggggccaact cctaccgtac ctcgcattac ccttacgctg aagagatgct cgactgggca 1560
gatgaacatg gcatcgtggt gattgatgaa actgctgctg tcggctttaa cctctcttta 1620
ggcattggtt tcgaagcggg caacaagccg aaagaactgt acagcgaaga ggcagtcaac 1680
ggggaaactc agcaagcgca cttacaggcg attaaagagc tgatagcgcg tgacaaaaac 1740
cacccaagcg tggtgatgtg gagtattgcc aacgaaccgg atacccgtcc gcaaggtgca 1800
cgggaatatt tcgcgccact ggcggaggca acgcgtaaac tcgacccgac gcgtccgatc 1860
acctgcgtca atgtaatgtt ctgcgacgct cacaccgata ccatcagcga tctctttgat 1920
gtgctgtgcc tgaaccgtta ttacggatgg tatgtccaaa gcggcgattt ggaaacggca 1980
gagaaggtac tggaaaaaga acttctggcc tggcaggaga aactgcatca gccgattatc 2040
atcaccgaat acggcgtgga tacgttagcc gggctgcact caatgtacac cgacatgtgg 2100
agtgaagagt atcagtgtgc atggctggat atgtatcacc gcgtctttga tcgcgtcagc 2160
gccgtcgtcg gtgaacaggt atggaatttc gccgattttg cgacctcgca aggcatattg 2220
cgcgttggcg gtaacaagaa agggatcttc actcgcgacc gcaaaccgaa gtcggcggct 2280
tttctgctgc aaaaacgctg gactggcatg aacttcggtg aaaaaccgca gcagggaggc 2340
aaacaatgaa ttcctgcagt agcgcgcagg agttccttat ttatgtcttt ccacgcattt 2400
gtgatctttt tccacatctc aacatatgct tcttgctttg aagttccata ttctttcatg 2460
tagtactcaa caaatgaagc aacatgtcct ctttgttgtt caacctacac ataatatatt 2520
tcaatatctt cattaatatt atacaatatt ttaccatttt aatcagttcg tttctaaagt 2580
tccataaatc actttatgta aatgtgctca ttgaagttgt aatttatcaa gcatgtctat 2640
ttattatatc atgcaaaagt gccacctaga ctcgaaatca gattgagaaa tgtttttttg 2700
agtcgagggt ctatcgggaa acaacctttc tacctccaca agatagggta agatcttcat 2760
acacatctct ctgcccagac ccgtactata ggaatacact ggatatgttg tttctgttat 2820
tgtagttgtt tttgtatgta tttagggaca aaatttgtga attaggatga ctaattgtta 2880
tataaaatgg aaattatatt gttatactta attcatgatc aacaatatca tccattaatc 2940
tacagattgt tgatgcagct cgaggaatca aaggttcatt tgccaaccat tcaaaagcct 3000
cttttgttat aatttcctcg ataacaacca aagcaaattg ttccatacat ctaccggtac 3060
aacttacaac tccatttttc atatactcct cacattttgg aatgtagtta gcattcaacc 3120
attcaacttc cttaaaatag gctgccacca actttttcat ctaaaaaagt aacagaaaaa 3180
gtactaatag ttcaaaattg tgattcttcc ctacaacttg gtttacttaa ttaacttatt 3240
aagtagtaat taatgtaact taatcaagga ttacctcttt tttcgcatag tagatgcgat 3300
ctgctttacc ttctttggcc attacttctt ccatttcatt gaaaacgtcg agaaggcctt 3360
gataaatagg tctcaaatat gccggtaatg aatccatggc acctacatcc catctgcaaa 3420
gtctaagttg tcataatata tttctctata acaacacttc aaatagcatc ttaaaaaatt 3480
atcagaacaa atgcagttgt tatagaaaga ttacctttgg atcgcatccg tgaaaagcat 3540
aagctcatca aaagttgcaa aagcatcata agtgtcatca atgatggagt tcatgttgac 3600
tacttgtgtt atcattcttc ttgaacgact atattgaggt tcaaaatata ttcccaccat 3660
ccaaaagtaa gcttc 3675
<210> 7
<211> 416
<212> DNA
<213> Artificial Sequence
<220>
<223> ddRNAi DNA construct (GW2) insert sequence
<400> 7
cttccctctt ctctttcttc ataaccctgt ctaaagggat tattatgata gtagagtctc 60
accatcgggc tcggattccg taaaagtcga aacgccacca attcggctga cacagcctcg 120
tgacatttaa atctttattg gtttgtgagc agggattgga ggcgttttga cttttagggg 180
atcggatcct cgaggtgtaa aaaaactcgt aaatcctatc agatctggaa gatttctacg 240
cttctccttc tttatattcg ttttcttatg ctttttattt ttgatataac ctagaaaaag 300
gctttttata tctttgaatc cgaaattgtt tgttttagaa tattgtatat ctgattttta 360
tcccttttta tatttgaatg ttctttagtc tccttttgtt tgcccaaatg ttgaat 416
<210> 8
<211> 4531
<212> DNA
<213> Nicotiana tabacum
<400> 8
ctcgtttggc gagagaaaga gagagatgag tcaatcaatt tctccattaa tctgttctca 60
ctttgcgaaa tttcagtcga atatttggag atgcaatact tctcaactca gagttataca 120
ctcatcatat gcctcttttg gagggagaag aaaagagaga gtaagaagaa tgaatcgagc 180
aatggatctt tcttcaagct ctcgtcattt ggcagatttt ccctcaacaa tttggggtga 240
ccattttctc tcctacaatt ctgaaataac agtaagtcac aatcccatgc ttatattcta 300
ccgtcaaact tctctataac aactccattt gttgcgatcg ttttttattg ttatagtgaa 360
gtgttgttat agagattata tattataata taacatagta atcggttccg agaaaacttg 420
attctttagt gaatgactgt tatatgggga tgttatagag aggtctgact gtgttaccta 480
aatgaataat gaagatgtta taattttatt agagatggat gaagcattta aacatgaaag 540
attcaagctt taagattggt ctttatcgta cttttgaaat tatgagtttg gaatttaata 600
cttcctccgt ctcaaaaaag aatgaatctt ttactatatg gggtcaaata agattttctt 660
tggccatatt ttttgcaaat gcatttcaaa tattttgaat ttttaattgt gtgacttaca 720
atacctttta tgcagtttct aaatgtgtaa aatatatttc aaacttttaa agaatattat 780
gtcatcacat taaaaatatt gaacttgact ctcatgctcc gacaggttaa ataatgaaac 840
gaatggaata ttactatttt gtaaaaactt aggtaggttt tcactatata tttaataatg 900
tattcaaact cataaataat gatcaaaagg agctttgacg ggaggtcacg ggttcgagcc 960
gtggaaacaa cctcttgcaa aaatgtaagg taagactgca tacaatagac tcttgtggtc 1020
cgccccttct ccggaccccg cacatagcgg aagcttattg cactaggctg ccctttttag 1080
tgattaacat atgtggatct aggatttaaa ttttttgggt tcaaccttta aggatctgtt 1140
acaattttag tagaatgtta cacataaatt tgtgctccgt gaatgtattg agtcagatga 1200
acctggtatt atacatgcgg atacgctcct gatgctcaat ttctgcctgc aggaaattac 1260
tacccaagag aaaaatgaac atgaaatgct aaaagaaata gttcggaaaa tgttggtaga 1320
aactccagat aatagtacac aaaaactagt cttgattgac acaattcaaa gattgggatt 1380
agcatatcat ttcaatgatg agattgaaaa ctccattcaa aacatcttta atttgtctca 1440
aaatagtgaa gatgacgatg aacacaacct ttatgttgct gctcttcgtt ttcgacttgc 1500
gaggcaacaa ggatattaca tgtcttcagg taccttacat ttctgccctt tcccgcacag 1560
cttcattttt tttcgttgtt aaaagacagt tcggcgcata aaatatctca tgtatacgcc 1620
agggtcagga cgaaccgccc ccaaggggtg taaagtatgc aacttaccct aatactaaat 1680
atctcgtgta tacacagggt caggacaagt cgcacccaag gggtgtaatg tagacaactt 1740
atcctaatgc tattagtaac tgattttatg gctcgaacac ataaattata ggtcacacag 1800
taacaacttt accgttgctc aaagactcgc cttcctcttt ttttagttat cgcaccttat 1860
ttgtgcagag aatagcaagt ttcgagatct gcttctatat agaagacttc tgtattatac 1920
ttttttattt tgtccttctg cttaaaaata gtaaaaaact atagtgtgga aattgtaaat 1980
ttcttaacta gctgtgaaat caaatagtta ttataggaat attatttaag actccactta 2040
tggaaaacca ctgggttgtt gttgttattg tcaataataa cttggggtac gatttacttc 2100
tttttccatg gcttgtccac gactatattt ctattaacaa tgttgtgact atgctttctt 2160
tgagtcgagg gtctattgat aacaggctct cgatctttac aaggtaaaag taatgtctgc 2220
gtacaccact ctactccgca gactccactt gtaggatttc actgaatatt ttttgttgtt 2280
gttgttgttg taataactta gggtttaatt tcttgatgct aatgaaattc atttctttca 2340
aaatataaac atggtgttca accagatgtg ttcaagcaat tcactaacca tgacggaaaa 2400
ttcaaggaaa atcatactaa tgatgttcaa ggattattga gtttgtatga agcagcacat 2460
atgagagtgc acgacgagga aattctagaa gaagctctta tctttaccac gactcatctc 2520
gagtccgtga tcccgaattt gagcaactcg cttaaggtac aagttactga agccttaagc 2580
catcctattc gcaaagctat accaagggtg ggagcaagga aatacataca catatatgaa 2640
aacattggaa cacataatga tttacttttg aaatttgcaa agttggactt caacatgtta 2700
caaaagcttc atcgaaaaga gcttaacgag ctaacaaggt acatctacta ttcttgtcat 2760
cttcataatt atggtacaat cagacctctc tctataaaat acatccttta taacaacagt 2820
tcactataac ggtcaagttt tctttaaaat caatgtttta tgttaccaaa ttattttgaa 2880
agaaatgtga ctattataga gaggtttgac tgtaactcgc gctaattaat aacacctaaa 2940
gtttaagtat gttaatgctg ttatgatatc tatagctggt ggaaagattt ggatcgtgca 3000
aacaaatttc catatgcaaa ggacagatta gtagaagctt acttttggac ggtgggaata 3060
tattttgaac ctcaatatag tcgttcaaga agtttggtaa caaaagtagt caaaatgaac 3120
tccattattg atgacactta tgatgcttat gcaacttttg atgagcttgt gcttttcacg 3180
gatgcgatcc aaaggtaaaa ttatatataa taaaatcttt ctataacaac gtcatttatt 3240
ctgatatttt tttaagatgc tatagtgaag tattgttata tagaaatata ttatgacaac 3300
ttagactttg cagatgggac gaaggtgcca tggatttatt accgacatat ctgagaccta 3360
tttatcaagg ccttctcgac gttttcaatg aaatggaaga agtattggcc aaagaaggta 3420
aagcagatca catctactat gcgaaaaaag aggtaatcct tgattaagtt acattaatta 3480
ctacttaata gttaattaag taaaccaagt tgtagggaag aatcgcaatt ttgaactatt 3540
agtacatttt ctgttacttt tttagatgaa aaaggtggcg gaagtctatt ttaaggaagc 3600
tgaatggttg aatgctaact acattccaaa atgcgaggag tatatgaaaa atggacttgt 3660
aagctctacc ggtccgatgt atggaataat ttctttggtt gttatggagg aaattataac 3720
aaaagaggct tttgaatggt tgacaaatga acctttgatt cttcgagctg catcaacaat 3780
ttgtagatta atggatgata tggctgatca tgaagtaagt ataacaatat aattttcatt 3840
ttatataaca atagccatcg taattcgcga attttgtccc taaatacaat acaaaaacaa 3900
ctacaataac aaaaacaaca tatccaatat attcctacag tacgggtcta ggaagagaga 3960
tgtgtacgca gatcttaccc taccttatag aggtagaaat gttgttcccg atagaccctc 4020
gactcaaaaa aagcatttct cagtctgatt tcgagtctag gtggcacttt tgcatgataa 4080
aataaataga catgcttgat aaattacaac ttcaatgatc acatttactt aaactgaatt 4140
atggaacttt agaaacggct gattaaaatg gtaaaatatt gtataataat gaagaaattg 4200
aaatatatta tgttgtaggt tgaacaacaa agaggacatg ttgcttcatt tgttgagtgc 4260
tacatgaaag aatatggagt ttcaaagcaa gaaacatatg ttgagatgcg gaaaaaaatc 4320
acaaatgcgt ggaaagatat aaataaggaa ctcttgcgcc ctactgcagt accaatgttt 4380
atcctcgaac gatctttaaa tttttcaaga ttggccgata catttttgaa agatgatgat 4440
ggatacacaa atcccaaatc caaagttaaa gacttgattg cttcgttgtt tgtcgaatct 4500
gtcgacatat gattatatat aacaatgcag a 4531
<210> 9
<211> 2251
<212> DNA
<213> Nicotiana tabacum
<400> 9
gctgcctttt ttttttttgg gctacaaaac tctaaagtaa aggaactaga aaactcgttt 60
ggcgagagaa agagagagat gagtcaatca atttctccat taatgttttc tcactttgca 120
aaatttcagt cgaatatttg gagatgcaat acttctcaac tcagagttat acactcatca 180
tatgcctctt ttggagggag aagaaaagag agagtaagaa gaatgaatcg agcaatggat 240
ctttcttcaa gctctcgtca tttggcagat tttccctcaa caatttgggg tgaccatttt 300
ctctcctaca attctgaaat aacagaaatt actacccaag agaaaaatga acatgaaatg 360
ctaaaagaaa tagttcggaa aatgttggta gaaactccag ataatagtac acaaaaacta 420
gtcttgattg acacaattca aagattggga ttagcatatc atttcaatga tgagattgaa 480
aactccattc aaaacatctt taatttgtct caaaatagtg aagatgacga tgaacacaac 540
ctttatgttg ctgctcttcg ttttcgactt gcgaggcaac aaggatatta catgtcttca 600
gatgtgttca agcaattcac taaccatgac ggaaaattca aggaaaatca tactaatgat 660
gttcaaggat tattgagttt gtatgaagca gcacatatga gagtgcacga cgaggaaatt 720
ctagaagaag ctcttatctt taccacgact catctcgagt ccgtgatccc gaatttgagc 780
aactcgctta aggtacaagt tactgaagcc ttaagccatc ctattcgcaa agctatacca 840
agggtgggag caaggaaata catacacata tatgaaaaca ttggaacaca taatgattta 900
cttttgaaat ttgcaaagtt ggacttcaac atgttacaaa agcttcatcg aaaagagctt 960
aacgagctaa caagctggtg gaaagatttg gatcgtgcaa acaaatttcc atatgcaaag 1020
gacagattag tagaagctta cttttggacg gtgggaatat attttgaacc tcaatatagt 1080
cgttcaagaa gtttggtaac aaaagtagtc aaaatgaact ccattattga tgacacttat 1140
gatgcttatg caacttttga tgagcttgtg cttttcacgg atgcgatcca aagatgggac 1200
gaaggtgcca tggatttatt accgacatat ctgagaccta tttatcaagg ccttctcgac 1260
gttttcaatg aaatggaaga agtattggcc aaagaaggta aagcagatca catttactat 1320
gcgaaaaaag agatgaaaaa ggtggcggaa gtctatttta aggaagctga atggttgaat 1380
gctaactaca ttccaaaatg cgaggagtat atgaaacatg gacttgtaag ctctaccggt 1440
ccgatgtatg gaataatttc tttggttgtt atggaggaaa ttataacaaa agaggctttt 1500
gaatggttga caaatgaacc tttgattctt cgacctgcat caacaatttg tagattaatg 1560
gatgatatgg ctgatcatga agttgaacaa caaagaggac atgttgcttc atttgttgag 1620
tgctacatga aagaatatgg agtttcaaag caagaagcat atgttgagat gcggaaaaaa 1680
atcacaaatg cgtggaaaga tataaataag gaactcttgc gccctactgc agtaccaatg 1740
tttatcctcg aacgatcttt aaatttttca agattggccg atacattttt gaaagatgat 1800
gatggataca caaatcccaa atccaaagtt aaagacttga ttgcttcgtt gtttgtcgaa 1860
tctgtcgaca tatgattata tataacaatg cagacacacc ttcaaagctg agtatttgga 1920
gcaaatatgg aagcattttg tattgtccat gtaaccctat aagtcacgtg tttgggcaat 1980
ggcaacattt actaatattt gcattatggt aggttgttta catcacacct attgggggcg 2040
acccttccta aaacctgaca tgaatgtgtg atgctttgtg cacctggcgg ctcattttta 2100
ctatttcact gttacaactt atttggacgg ttgttaacct attgaatcat gtagtattgt 2160
tacttaaata caatgtttat tttaattatt acttaaattt tattctatca tatcgttaaa 2220
tccatcatta cgtaacaaaa aaaaaaaaaa a 2251
<210> 10
<211> 598
<212> PRT
<213> Nicotiana tabacum
<400> 10
Met Ser Gln Ser Ile Ser Pro Leu Met Phe Ser His Phe Ala Lys Phe
1 5 10 15
Gln Ser Asn Ile Trp Arg Cys Asn Thr Ser Gln Leu Arg Val Ile His
20 25 30
Ser Ser Tyr Ala Ser Phe Gly Gly Arg Arg Lys Glu Arg Val Arg Arg
35 40 45
Met Asn Arg Ala Met Asp Leu Ser Ser Ser Ser Arg His Leu Ala Asp
50 55 60
Phe Pro Ser Thr Ile Trp Gly Asp His Phe Leu Ser Tyr Asn Ser Glu
65 70 75 80
Ile Thr Glu Ile Thr Thr Gln Glu Lys Asn Glu His Glu Met Leu Lys
85 90 95
Glu Ile Val Arg Lys Met Leu Val Glu Thr Pro Asp Asn Ser Thr Gln
100 105 110
Lys Leu Val Leu Ile Asp Thr Ile Gln Arg Leu Gly Leu Ala Tyr His
115 120 125
Phe Asn Asp Glu Ile Glu Asn Ser Ile Gln Asn Ile Phe Asn Leu Ser
130 135 140
Gln Asn Ser Glu Asp Asp Asp Glu His Asn Leu Tyr Val Ala Ala Leu
145 150 155 160
Arg Phe Arg Leu Ala Arg Gln Gln Gly Tyr Tyr Met Ser Ser Asp Val
165 170 175
Phe Lys Gln Phe Thr Asn His Asp Gly Lys Phe Lys Glu Asn His Thr
180 185 190
Asn Asp Val Gln Gly Leu Leu Ser Leu Tyr Glu Ala Ala His Met Arg
195 200 205
Val His Asp Glu Glu Ile Leu Glu Glu Ala Leu Ile Phe Thr Thr Thr
210 215 220
His Leu Glu Ser Val Ile Pro Asn Leu Ser Asn Ser Leu Lys Val Gln
225 230 235 240
Val Thr Glu Ala Leu Ser His Pro Ile Arg Lys Ala Ile Pro Arg Val
245 250 255
Gly Ala Arg Lys Tyr Ile His Ile Tyr Glu Asn Ile Gly Thr His Asn
260 265 270
Asp Leu Leu Leu Lys Phe Ala Lys Leu Asp Phe Asn Met Leu Gln Lys
275 280 285
Leu His Arg Lys Glu Leu Asn Glu Leu Thr Ser Trp Trp Lys Asp Leu
290 295 300
Asp Arg Ala Asn Lys Phe Pro Tyr Ala Lys Asp Arg Leu Val Glu Ala
305 310 315 320
Tyr Phe Trp Thr Val Gly Ile Tyr Phe Glu Pro Gln Tyr Ser Arg Ser
325 330 335
Arg Ser Leu Val Thr Lys Val Val Lys Met Asn Ser Ile Ile Asp Asp
340 345 350
Thr Tyr Asp Ala Tyr Ala Thr Phe Asp Glu Leu Val Leu Phe Thr Asp
355 360 365
Ala Ile Gln Arg Trp Asp Glu Gly Ala Met Asp Leu Leu Pro Thr Tyr
370 375 380
Leu Arg Pro Ile Tyr Gln Gly Leu Leu Asp Val Phe Asn Glu Met Glu
385 390 395 400
Glu Val Leu Ala Lys Glu Gly Lys Ala Asp His Ile Tyr Tyr Ala Lys
405 410 415
Lys Glu Met Lys Lys Val Ala Glu Val Tyr Phe Lys Glu Ala Glu Trp
420 425 430
Leu Asn Ala Asn Tyr Ile Pro Lys Cys Glu Glu Tyr Met Lys His Gly
435 440 445
Leu Val Ser Ser Thr Gly Pro Met Tyr Gly Ile Ile Ser Leu Val Val
450 455 460
Met Glu Glu Ile Ile Thr Lys Glu Ala Phe Glu Trp Leu Thr Asn Glu
465 470 475 480
Pro Leu Ile Leu Arg Pro Ala Ser Thr Ile Cys Arg Leu Met Asp Asp
485 490 495
Met Ala Asp His Glu Val Glu Gln Gln Arg Gly His Val Ala Ser Phe
500 505 510
Val Glu Cys Tyr Met Lys Glu Tyr Gly Val Ser Lys Gln Glu Ala Tyr
515 520 525
Val Glu Met Arg Lys Lys Ile Thr Asn Ala Trp Lys Asp Ile Asn Lys
530 535 540
Glu Leu Leu Arg Pro Thr Ala Val Pro Met Phe Ile Leu Glu Arg Ser
545 550 555 560
Leu Asn Phe Ser Arg Leu Ala Asp Thr Phe Leu Lys Asp Asp Asp Gly
565 570 575
Tyr Thr Asn Pro Lys Ser Lys Val Lys Asp Leu Ile Ala Ser Leu Phe
580 585 590
Val Glu Ser Val Asp Ile
595
<210> 11
<211> 416
<212> DNA
<213> Artificial Sequence
<220>
<223> ddRNAi DNA construct (GW5) insert sequence
<400> 11
cttccctctt ctctttcttc ataaccctgt ctaaagggat tattatgata gtagagtctc 60
accatcgggc tcggattcga agaaatcatc caccgctcca attcggctga cacagcctcg 120
tgacatttaa atctttattg gtttgtgagc agggattgga gcggtggagg atttcttagt 180
atcggatcct cgaggtgtaa aaaaactcgt aaatcctatc agatctggaa gatttctacg 240
cttctccttc tttatattcg ttttcttatg ctttttattt ttgatataac ctagaaaaag 300
gctttttata tctttgaatc cgaaattgtt tgttttagaa tattgtatat ctgattttta 360
tcccttttta tatttgaatg ttctttagtc tccttttgtt tgcccaaatg ttgaat 416
<---
Реферат
Изобретение относится к области биохимии, в частности к способу увеличения содержания сложного эфира сахарозы в растении табака. Также раскрыты применение полинуклеотида в указанном способе; применение указанного табачного продукта для производства табачного экстракта или в устройстве для нагрева табака; способ получения растения с помощь указанного способа; способ приготовления табачного экстракта. Раскрыты модифицированное растение или его часть, содержащее повышенное количество сложного эфира сахарозы, изделие табачной промышленности, бездымный табачный продукт. Изобретение позволяет получать растения табака с увеличенным содержанием сложного эфира сахарозы. 27 н. и 13 з.п. ф-лы, 17 ил., 10 табл., 7 пр.
Комментарии