Растения со сниженным содержанием аспарагина - RU2742725C2
Код документа: RU2742725C2
Чертежи




















Описание
ОБЛАСТЬ ТЕХНИКИ
В настоящем изобретении раскрыты полинуклеотидные последовательности генов, кодирующих аспарагинсинтетазу из Nicotianatabacum, и варианты, гомологи и фрагменты указанных последовательностей. Также раскрыты полипептидные последовательности, кодируемые указанными последовательностями, и варианты, гомологи и фрагменты данных последовательностей. Также раскрыта модификация экспрессии одного или нескольких генов или активности белка или белков, кодируемых указанными генами, для модулирования уровней аспарагина в растении.
УРОВЕНЬ ТЕХНИКИ
Акриламид представляет собой химическое соединение формулы C3H5NO (название согласно ИЮПАК, Международному союзу теоретической и прикладной химии, проп-2-енамид), и потенциальная токсичность данного соединения вызывает озабоченность. Источником акриламида в аэрозоле курительных изделий, содержащих табак, может являться аминокислота аспарагин, которая может присутствовать в табачном материале, используемом для производства указанных курительных изделий. Вследствие этого может быть желательно снизить уровни определенных аминокислот, таких как аспарагин, в растениях, в частоности, в тех растениях, которые используют для производства табака.
В публикации US2013/0068240 описаны генетически модифицированные растения табака и табачные продукты, в которых снижен уровень по меньшей мере одной аминокислоты, в результате чего после нагревания и/или горения в табаке образуются сниженные уровни соединения, образующегося из по меньшей мере одной аминокислоты, по сравнению с немодифицированным родительским растением табака. Был описан мутагенез семян с применением этилметансульфоната (ЭМС) для получения мутантных линий табака со сниженными уровнями аспарагина и сниженными уровнями акриламида в сигаретном дыме. Различные мутанты ЭМС описаны в публикации US2013/0068240. В листьях верхней части стебля после паровой обработки (томления) три мутанта ЭМС, а именно FC Up 10NH-5, FC Up 10NH-18 и FC-Up 10NH-23, продемонстрировали: (i) снижение уровня аспарагина на 71%, 30% и 62%, соответственно; (ii) снижение уровня акриламида в табачном дыме на 47%, 42% и 44%, соответственно; и (iii) снижение на 38%, повышение на 111% и снижение на 19% уровня никотина в табаке верхней части стебля после паровой обработки, соответственно. Таким образом, наибольшее снижение уровня аспарагина и акриламида, составляющее 71% и 47%, соответственно, было достигнуто в случае мутанта FC Up 10NH-5. Однако в данном мутанте наблюдалось снижение уровней никотина на 38% по сравнению с контролем. В листьях средней части стебля после паровой обработки три мутанта ЭМС, а именно FC Mid10NH-5, FC Up 10NH-18 и FC-Up 10NH-23, продемонстрировали: (i) снижение на 6%, повышение на 531% и снижение на 29% уровня аспарагина, соответственно; (ii) снижение на 19%, повышение на 140% и повышение на 200% уровня акриламида в табачном дыме, соответственно; и (iii) снижение на 36%, повышение на 113% и снижение на 19% уровня никотина в табаке средней части стебля после паровой обработки, соответственно. Таким образом, наибольшее снижение уровня акриламида, составляющее 19%, было достигнуто в случае FC Up 10NH-5, однако уровень аспарагина был снижен всего лишь на 6%, и уровень никотина был снижен на 36%.
Наибольшее снижение уровня аспарагина и акриламида, составляющее 71% и 47%, соответственно, было обнаружено в листьях верхней части стебля FC Up 10NH-5 после паровой обработки. Однако в данном мутанте наблюдалось снижение уровней никотина на 38% по сравнению с контролем. В листьях средней части стебля FC Up 10NH-5 после паровой обработки уровень акриламида был снижен лишь на 19%, уровень аспарагина был снижен лишь на 6%, и вновь наблюдалось снижение уровней никотина (составляющее 36%) по сравнению с контролем.
Снижение уровня никотина является нежелательным, поскольку потребитель табачного продукта будет вынужден употреблять больше табака для получения того же уровня никотина. Также из результатов, представленных в публикации US2013/0068240, очевидно, что уровни аспарагина, акриламида и никотина широко и произвольным образом варьируют среди различных мутантов ЭМС.
Желательно обнаружить альтернативные способы снижения уровней аспарагина в растении и посредством этого минимизировать количество акриламида в аэрозоле, образованном из нагретой или сгоревшей части растения. Также желательно найти способы дополнительного снижения количества акриламида до уровней ниже уровней, раскрытых в публикации US2013/0068240. Также желательно сохранить уровни никотина в растении. Также желательно разработать способы, которые позволят стабильно получать растения с данными свойствами. Настоящее изобретение направлено на удовлетворение этой потребности.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Было идентифицировано шесть генов полноразмерной аспарагинсинтетазы в Nicotiana tabacum, которые называют NtASN1-S (SEQ ID NO: 1), NtASN1-T (SEQ ID NO: 3), NtASN3-S (SEQ ID NO: 9), NtASN3-T (SEQ ID NO: 11), NtASN5-S (SEQ ID NO: 5) и NtASN5-T (SEQ ID NO: 7). Было обнаружено, что гены ASN3-S и ASN3-T исключительно слабо экспрессируются в ходе обработки, тогда как неожиданно было обнаружено, что NtASN1-S, NtASN1-T, NtASN5-S и NtASN5-T в высокой степени экспрессируются в ходе обработки, например, в течение первых 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 часов обработки. Предпочтительно, гены ASN3-S или ASN3-T или белки, кодируемые указанными генами, не применяют в настоящем изобретении. В данном документе показано, что снижение экспрессии NtASN1 и/или NtASN5 или активности белка или белков, кодируемых указанными генами, способствует снижению уровней аспарагина в обработанных листьях - таких как обработанный лист средней-нижней части стебля. Это, в свою очередь, приводит к снижению уровней акриламида в аэрозоле, который образуется после нагревания или сгорания обработанного листа. В отличие от результатов, полученных в публикации US2013/0068240, снижение экспрессии NtASN1 или NtASN5 не толькоприводит к снижению уровня аспарагина в листьях, но также стабильно приводит к снижению уровня акриламида в аэрозоле. Стабильное снижение уровней аспарагина и акриламида сопровождается уровнями никотина в обработанных листьях, которые являются приблизительно эквивалентными, если не большими, чем уровни в контрольном растении. Предпочтительно, в некоторых вариантах реализации настоящего изобретения можно получить более низкие значения, соответствующие приблизительно 89% снижению уровня аспарагина и приблизительно 70% снижению уровня акриламида, с по существу минимальным влиянием или отсутствием влияния на содержание никотина.
Без ограничения какой-либо тиорией, также определили, что акриламид, образующийся в аэрозоле табака, зависит от пула аспарагина, который образуется в ходе обработки листьев. Соответственно, уровни акриламида в аэрозоле можно модулировать (например, снижать) посредством снижения пула аспарагина, который образуется в ходе обработки или сушки, посредством модулирования (например, снижения) экспрессии или активности аспарагинсинтетазы, которая активна в течение ранней фазы обработки или сушки. Описаны растения, которые характеризуются сниженным содержанием аспарагина и демонстрируют сниженную экспрессию или функцию аспарагинсинтетазы. В вариантах реализации этого достигают с применением РНК-интерференции или мутации последовательностей, кодирующих аспарагинсинтетазу, описанных в данном документе, которые экспрессируются в ходе обработки или сушки. Снижение уровней аспарагина снижает количество акриламида, присутствующее в аэрозоле. Применительно к табаку, это приводит к меньшим количествам акриламида, присутствующим в аэрозоле, который вдыхает потребитель курительного изделия, включая нагретый или сгоревший табачный продукт. Другие преимущества настоящего изобретения описаны в данном документе.
АСПЕКТЫ И ВАРИАНТЫ РЕАЛИЗАЦИИ ИЗОБРЕТЕНИЯ
Аспекты и варианты реализации настоящего изобретения изложены в прилагаемой формуле изобретения.
В одном аспекте предложено мутантное, неприродное или трансгенное растение или его часть, которое характеризуется сниженной экспрессией или активностью аспарагинсинтетазы, причем указанная аспарагинсинтетаза содержит, состоит из или состоит по существу из: (i) полинуклеотидной последовательности, содержащей, состоящей из или состоящей по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 72% или по меньшей мере 90% с SEQ ID NO:1 или идентичностью последовательности по меньшей мере 72% с SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7; или (ii) полипептида, кодируемого полинуклеотидом, описанным в (i); или (iii) полипептида, который характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8; причем экспрессия или активность аспарагинсинтетазы, изложенной в (i), (ii) или (iii), снижена по сравнению с контрольным растением.
Предпочтительно, растение содержит по меньшей мере одно генетическое изменение в регуляторной области или в кодирующей последовательности полинуклеотида, кодирующего аспарагинсинтетазу.
Предпочтительно, сниженная экспрессия или активность аспарагинсинтетазы обеспечивает снижение уровня аспарагина в обработанном или высушенном листе, полученном из мутантного, неприродного или трансгенного растения или части указанного растения, по сравнению с уровнем аспарагина в обработанном или высушенном листе, полученном из контрольного растения, причем, предпочтительно, уровень аспарагина снижен по меньшей мере приблизительно на 17% по сравнению с контрольным растением.
Предпочтительно, сниженная экспрессия или активность аспарагинсинтетазы обеспечивает снижение уровня акриламида в аэрозоле, который образуется после нагревания или сгорания обработанного или высушенного листа, полученного из мутантного, неприродного или трансгенного растения или части указанного растения, по сравнению с обработанным или высушенным листом, полученным из контрольного растения, причем, предпочтительно, уровень акриламида в аэрозоле снижен по меньшей мере на 20% по сравнению с обработанным или высушенным листом контрольного растения.
Предпочтительно, уровень аспарагина в обработанном или высушенном листе, полученном из мутантного, неприродного или трансгенного растения или части указанного растения, является по меньшей мере приблизительно на 22% меньшим, и количество акриламида в аэрозоле, который образуется после нагревания или сгорания обработанного или высушенного листа, полученного из мутантного, неприродного или трансгенного растения или части указанного растения, является по меньшей мере на 24% меньшим, чем в обработанном или высушенном листе и аэрозоле из контрольного растения; причем, предпочтительно, уровень аспарагина в обработанном или высушенном листе, полученном из мутантного, неприродного или трансгенного растения или части указанного растения, является по меньшей мере приблизительно на 44% меньшим, и количество акриламида в аэрозоле, который образуется после нагревания или сгорания обработанного или высушенного листа, полученного из мутантного, неприродного или трансгенного растения или части указанного растения, является по меньшей мере на 66% ниже по сравнению с обработанным или высушенным листом и аэрозолем из контрольного растения; причем, предпочтительно, уровень аспарагина в обработанном или высушенном листе, полученном из мутантного, неприродного или трансгенного растения или части указанного растения, является по меньшей мере приблизительно на 70% меньшим, и количество акриламида в аэрозоле, который образуется после нагревания или сгорания обработанного или высушенного листа, полученного из мутантного, неприродного или трансгенного растения или части указанного растения, является по меньшей мере приблизительно на 88% ниже по сравнению с обработанным или высушенным листом и аэрозолем из контрольного растения.
Предпочтительно, уровень никотина в обработанном или высушенном листе из мутантного, неприродного или трансгенного растения или части указанного растения является по существу таким же, как и уровень никотина в обработанном или высушенном листе контрольного растения.
Предпочтительно, образование глутамина, аспарагиновой кислоты и глутаминовой кислоты повышено в обработанном или высушенном листе, полученном из мутантного, неприродного или трансгенного растения или части указанного растения, по сравнению с обработанным или высушенным листом контрольного растения.
Предпочтительно, растение выбрано из группы, состоящей из табака или чая (Camellia sinensis).
Предпочтительно, растение табака представляет собой растение табака Nicotiana tabacum, предпочтительно, растение табака Nicotiana tabacum типа Берли.
Предпочтительно, растение содержит по меньшей мере одну мутацию в каждой копии полинуклеотидной последовательности, кодирующей аспарагинсинтетазу, изложенную в (i), (ii) или (iii), предпочтительно, стоп-мутацию или мутации и/или фрагмент или фрагменты гена, которые препятствуют трансляции РНК-транскрипта, кодирующего аспарагинсинтетазу.
Предпочтительно, нуклеотидная последовательность аспарагинсинтетазы содержит нуклеотидную последовательность, кодирующую стоп-мутацию в положении, эквивалентном расположению нуклеотидной последовательности, кодирующей стоп-мутацию, в SEQ ID No. 19, 21 и 23, или причем аспарагинсинтетаза содержит нуклеотидную последовательность, кодирующую стоп-кодон, в положении, эквивалентном расположению стоп-кодона в SEQ ID No. 20, 22 и 24.
Предпочтительно, аспарагинсинтетаза содержит нуклеотидную последовательность, которая выбрана из группы, состоящей из SEQ ID No. 19, 21 и 23, или кодируется нуклеотидной последовательностью, которая выбрана из группы, состоящей из SEQ ID No. 19, 21 и 23, или содержит, состоит из или состоит по существу из полипептидной последовательности, представленной в SEQ ID No. 20, 22 и 24.
Предпочтительно, количество биомассы листа из мутантного, неприродного или трансгенного растения или части указанного растения является по существу таким же, как и количество биомассы листа из контрольного растения.
Предпочтительно, лист обрабатывают посредством воздушной обработки, причем, предпочтительно, лист, обработанный посредством воздушной обработки, является обработанным на солнце или обработанным посредством огневой обработки.
Предпочтительно, лист высушивают посредством воздушной сушки, причем, предпочтительно, лист, высушенным посредством воздушной сушки, является обработанным на солнце или высушенным посредством огневой сушки.
В другом аспекте предложен растительный материал или обработанный или высушенный растительный материал из растения согласно настоящему изобретению.
В другом аспекте предложен растительный продукт, содержащий по меньшей мере часть растения или растительного материала согласно настоящему изобретению.
В другом аспекте предложен способ получения растительного материала со сниженными уровнями аспарагина и сниженными уровнями акриламида в аэрозоле, полученном из указанного растительного материала, по сравнению с растительным материалом из контрольного растения, причем указанный способ включает следующие стадии: (a) обеспечение растения или части указанного растения, которое содержит полинуклеотид, содержащий, состоящий из или состоящий по существу из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% или по меньшей мере 90% с SEQ ID NO:1 или идентичностью последовательности по меньшей мере 72% с SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7; (b) снижение экспрессии полинуклеотида или активности белка, кодируемого указанным полинуклеотидом, в растении или части указанного растения; (c) сбор растительного материала из растения или части указанного растения; (d) обработку или сушку растительного материала; (e) необязательно, измерение уровней аспарагина в растении или части указанного растения и/или измерение уровней акриламида в аэрозоле, полученном из растения или части указанного растения; и (f) получение обработанного или высушенного растительного материала со сниженными уровнями аспарагина и сниженными уровнями акриламида в аэрозоле, полученном из указанного растительного материала, причем, предпочтительно, уровень никотина является по существу таким же, как и уровень никотина в контрольном растении; причем, предпочтительно, образование глутамина, аспарагиновой кислоты и глутаминовой кислоты повышено по сравнению с обработанным или высушенным листом контрольного растения.
Предпочтительно, растительный материал обрабатывают или высушивают в течение по меньшей мере приблизительно 3 дней после сбора.
Предпочтительно, растительный материал обрабатывают посредством воздушной обработки, причем, предпочтительно, обработанный посредством воздушной обработки лист является обработанным на солнце или обработанным посредством огневой обработки.
Предпочтительно, растительный материал высушивают посредством воздушной сушки или сушки на солнце, или высушивают посредством огневой сушки.
Также раскрыт растительный материал, полученный или получаемый посредством данного способа.
В другом аспекте предложен способ идентификации одного или нескольких генетических изменений в растении, которые коррелируют со сниженными уровнями аспарагина в обработанном или высушенном растительном материале из растения и сниженными уровнями акриламида в аэрозоле, полученном из обработанного или высушенного растения или части указанного растения, по сравнению с контрольным растением, которое не содержит одного или нескольких генетических изменений, причем указанный способ включает следующие стадии: (a) идентификацию обработанного или высушенного растения со сниженными уровнями аспарагина в растительном материале из растения и сниженными уровнями акриламида в аэрозоле, полученном из обработанного или высушенного растительного материала, по сравнению с растительным материалом из контрольного растения; (b) обеспечение образца нуклеиновой кислоты из растения, идентифицированного на стадии (a); и (c) идентификацию в образце нуклеиновой кислоты со стадии (b) одного или нескольких генетических изменений в полинуклеотидной последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% или по меньшей мере 90% с SEQ ID NO:1 или идентичностью последовательности по меньшей мере 72% с SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7.
В другом аспекте предложен способ получения растительного материала со сниженными уровнями аспарагина и сниженными уровнями акриламида в аэрозоле, полученном из обработанного или высушенного растительного материала, по сравнению с контрольным растением, причем указанный способ включает следующие стадии: (a) обеспечение растения или растительного материала согласно настоящему изобретению; (b) сбор растительного материала из растения; (c) обработку или сушку растительного материала в течение периода времени; и (d) получение обработанного или высушенного растительного материала, который содержит сниженные уровни аспарагина и сниженные уровни акриламида в аэрозоле, полученном из обработанного или высушенного растительного материала, причем, предпочтительно, уровень никотина является по существу таким же, как и уровень никотина в контрольном растении; причем, предпочтительно, образование глутамина, аспарагиновой кислоты и глутаминовой кислоты повышено по сравнению с контрольным растением.
В другом аспекте предложена выделенная полинуклеотидная последовательность, содержащая, состоящая из или состоящая по существу из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% или по меньшей мере 90% с SEQ ID NO:1 или идентичностью последовательности по меньшей мере 72% с SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или содержащая, состоящая из или состоящая по существу из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID No 19, 21 и 23, при условии, что последовательность, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID No 19, 21 и 23, содержит нуклеотидную последовательность, кодирующую стоп-кодон, в положении, эквивалентном расположению стоп-кодона в SEQ ID No. 19, 21 и 23, причем, предпочтительно, указанный выделенный полинуклеотид представляет собой синтетический полинуклеотид или кДНК.
В другом аспекте предложен выделенный полипептид, кодируемый полинуклеотидом, описанным в данном документе.
В другом аспекте предложен выделенный полипептид, содержащий, состоящий из или состоящий по существу из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO: 20 или SEQ ID NO: 22 или SEQ ID NO: 24, или содержащий, состоящий из или состоящий по существу из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO: 20 или SEQ ID NO: 22 или SEQ ID NO: 24, при условии, что последовательность, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO: 20 или SEQ ID NO: 22 или SEQ ID NO: 24, содержит стоп-кодон в положении, эквивалентном расположению стоп-кодона в SEQ ID NO: 20 или SEQ ID NO: 22 или SEQ ID NO: 24, причем, предпочтительно, указанный выделенный полинуклеотид представляет собой синтетический полинуклеотид или кДНК.
В другом аспекте предложена конструкция, вектор или вектор экспрессии, содержащий выделенный полинуклеотид, описанный в данном документе.
В другом аспекте предложена клетка мутантного, неприродного или трансгенного растения из растения, описанного в данном документе.
В другом аспекте предложен обработанный или высушенный растительный материал, содержащий клетку, описанную в данном документе.
В другом аспекте предложен табачный продукт или курительное изделие, содержащее растительный материал, описанный в данном документе.
Также раскрыты комбинации одного или нескольких изложенных вариантов реализации.
НЕКОТОРЫЕ ПРЕИМУЩЕСТВА
В данном документе показано, что аспарагин активно образуется в ходе обработки. Было показано, что данное накопление аспарагина приводит к образованию акриламида в аэрозоле, который образуется, когда обработанный растительный материал нагревают или сжигают. Предпочтительно, NtASN1-S, NtASN1-T, NtASN5-S и NtASN5-T в высокой степени экспрессируются в ходе обработки, в особенности, с самого начала обработки. Снижение экспрессии одного или нескольких из данных генов может привести к более низким уровням акриламида в аэрозоле, поскольку уровень аспарагина может быть снижен в течение процесса обработки.
Наблюдается ограниченное влияние на уровни никотина в модифицированных растениях, описанных в данном документе, которое является желательным, когда модифицированные растения предполагается использовать для получения растений табака.
Способы, описанные в данном документе, последовательно приводят к получению растений, в которых снижено накопление аспарагина в ходе обработки или сушки, а также снижено образование акриламида в аэрозоле.
Наблюдается ограниченное влияние на биомассу листа и фенотип, что характеризуется преимуществом при использовании описанных в данном документе модифицированных растений для коммерческих целей.
Настоящее изобретение позволяет создавать не модифицированные генетическим способом растения, которые могут являться более пригодными для потребителей.
Настоящее изобретение не ограничено применением мутантных растений ЭМС, как в публикации US2013/0068240.Мутантное растение ЭМС может характеризоваться меньшим потенциалом передавать улучшенные свойства урожаю после селекции. После начала селекции желаемая характеристика или характеристики мутантного растения ЭМС могут быть утрачены по различным причинам. Например, может потребоваться несколько мутаций, мутация может являться доминантной или рецессивной, и идентификация точковой мутации в гене-мишени может являться труднодостижимой. Напротив, в настоящем изобретении используют применение определенных генов, с которыми можно специфично манипулировать и/или которые можно нокаутировать для получения растений с желаемым фенотипом. Настоящее изобретение можно применять в отношении других разновидностей или сельскохозяйственных культур табака, и посредством этого предлагать желаемое решение для снижения по меньшей мере уровня акриламида.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фигуре 1 представлено содержание аспартата (Asp), глутамина (Glu), аспарагина (Asn) и суммарных свободных аминокислот (АК) в 3 различных табаках Берли, которые культивировали на одном поле (среднее из 5 повторов для участка) и обрабатывали одновременно. Три Берли представляли собой TN90 (A), Бенкет A1 (B) и Кентукки 14 (C).
На фигуре 2 представлены зависимости от времени процесса изменения уровня аспарагина, глутамина и аспартата в ходе обработки листьев Берли.
На фигуре 3 представлен путь активности аспарагинсинтетазы.
На фигуре 4 представлена корреляция, существующая между присутствием аспарагина в отдельных сортах табака Вирджиния (FC), Ориентал (OR), Кастури (KS) или Берли (BU) и в композиции, содержащей смесь Вирджинии (FC), Кастури (KS) и Ориентал (OR) (обозначена RRP).
На фигуре 5 представлены результаты, полученные, когда швейцарский Берли Стелла выращивали в Швейцарии и обрабатывали либо в амбаре для воздушной обработки в течение 10 недель (классическая сельскохозяйственная методика, BU-AC), либо в печи для паровой обработки, точно как же, как в случае швейцарского табака Вирджиния (BU-FC). После обработки листовую пластинку измельчали и проводили определение аспарагина (A). Готовили гомогенизированный порошок табачного материала (меньше или равно 200 микрон). Гомогенизированный табачный материал использовали в изделии, генерирующем аэрозоль, по существу как описано в публикации WO2013098405, для применения с устройством, генерирующим аэрозоль. Изделие, генерирующее аэрозоль, выкуривали с применением режима выкуривания согласно Министерству здравоохранения Канады для определения содержания акриламида в аэрозоле (B).
На фигуре 6 представлено филогенетическое дерево аминокислотных последовательностей, предсказанных на основании генов табака, соответствующих NtASN1-S, NtASN1-T, NtASN3-S, NtASN3-T, NtASN5-S, NtASN5-T, по сравнению с белками арабидопсиса AtASN1 (At3g47340), AtASN2 (At5g65010) и AtASN3 (At5g10240) и белками томата Solyc06g007180 и Solyc04g055200.
На фигуре 7 представлена экспрессия генов ASN в ходе обработки табака швейцарский Берли (Стелла), выращенного в Швейцарии в 2010 году. ARN выделяли из образцов листа (расположенных в средней части стебля), отобранных в зависимости от времени в поле (-4 ч) и в амбарах для воздушной обработки до 96 ч обработки. Экспрессию генов на специфичных зондах определяли посредством использования микроматричного анализа с чипами Affymetrix Tobarray.
На фигуре 8 представлена экспрессия генов ASN в табаке швейцарский Берли (Стелла, A и C) и швейцарская Вирджиния (Брайт, ITB 683, B и B) через 48 ч обработки.
На фигуре 9 представлена послеуборочная экспрессия ASN1, ASN5 (A) и ASN3 (B) через 60 ч воздушной обработки в 4 различных листьях Берли (расположенных в нижней части стебля), выращенного на поле Пайерн в 2013 году.
На фигуре 10 представлена экспрессия ASN1, ASN5 (левый чертеж) и ASN3 (правый чертеж) в 8 тканях, отобранных от зрелого табака Берли, выращенного в поле (Незрелый цветок (Незрел._цветок; Лист из нижней, _НС, средней,_СС и верхней, _ВС, части стебля; Лепесток, Корень, Чашелистик и Стебель).
На фигуре 11 представлено содержание аспарагина в растениях ASN1-RNAi, выращенных в теплице, под контролем промотора MMV или E4 и уровни транскрипта ASN1, определенные через 3 дня обработки. Количество аспарагина определяли в листьях, расположенных в средней-нижней части стебля, от 15 растений от каждого события трансформации (A). Относительную экспрессию транскрипта ASN1 оценивали методом кПЦР (количественной полимеразной цепной реакции) с применением РНК, отобранной через 3 дня обработки, в 15 растениях от каждого события трансформации (B).
На фигуре 12 представлено содержание глутамина, аспартата, глутамата и никотина в растениях ASN1-RNAi, выращенных в теплице, под контролем промотора MMV или E4. Количество Gln (A), Asp (B), Glu (C) и никотина (D) измеряли в листьях, расположенных в средней-нижней части стебля, от 15 растений от каждого события трансформации.
На фигуре 13 представлено содержание аспарагина в растениях ASN5-RNAi, выращенных в теплице, под контролем промотора MMV или E4 и уровни транскрипта ASN5, определенные через 3 дня обработки. Количество аспарагина определяли в листьях, расположенных в средней-нижней части стебля, от 15 растений от каждого события трансформации (A). Относительную экспрессию транскрипта ASN5 оценивали методом кПЦР (количественной полимеразной цепной реакции) с применением РНК, отобранной через 3 дня обработки, в 15 растениях от каждого события трансформации (B).
На фигуре 14 представлено содержание глутамина, аспартата, глутамата и никотина в растениях ASN5-RNAi, выращенных в теплице, под контролем промоторов MMV или E4. Количество Gln (A), Asp (B), Glu (C) и никотина (D) измеряли в листьях, расположенных в средней-нижней части стебля, от 15 растений от каждого события трансформации.
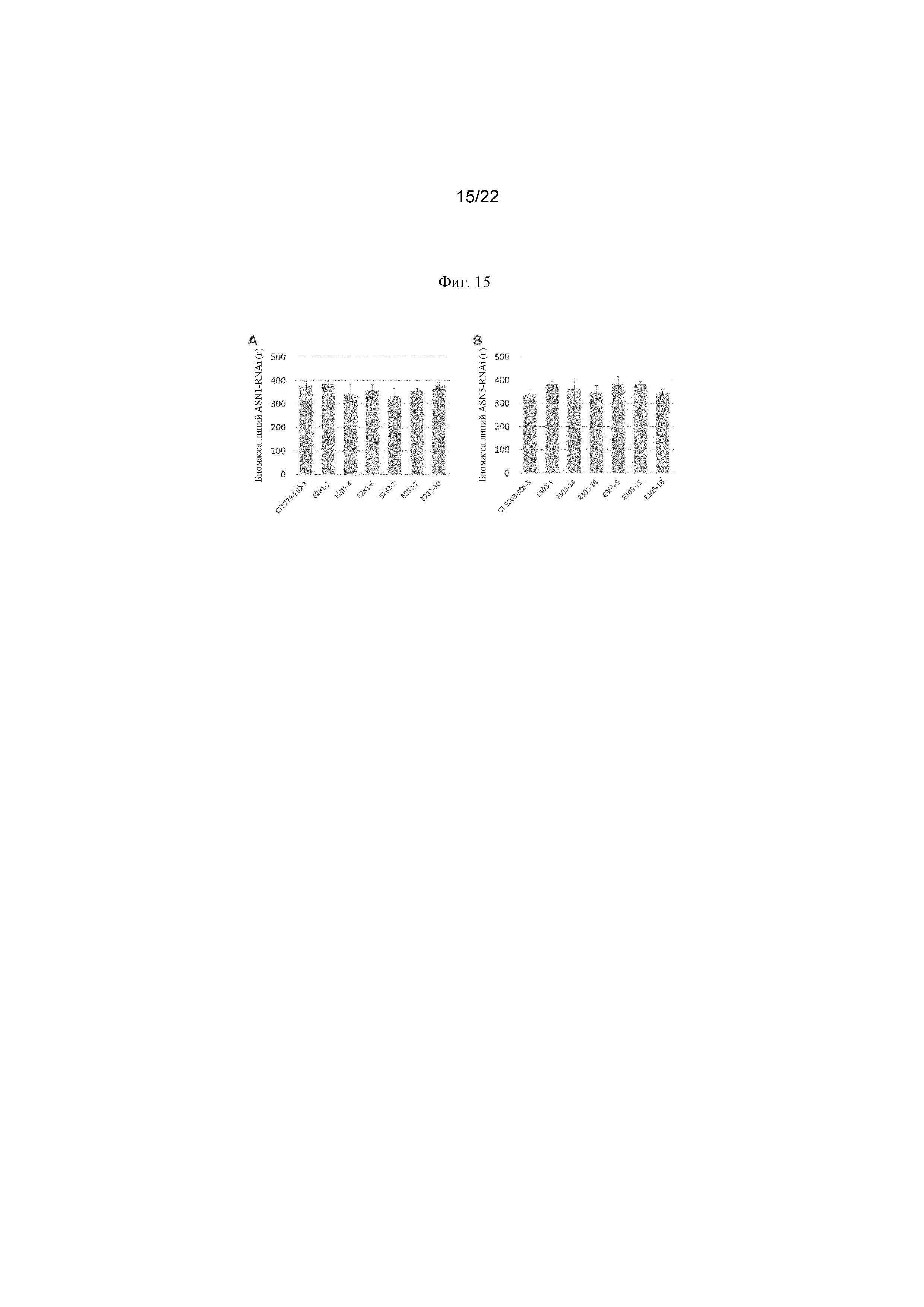
На фигуре 15 представлена биомасса листа линий ASN1- и ASN5-RNA. Отбирали четыре листа в средней части стебля от 4 растений от каждого события трансформации, выбранных случайным образом и взвешенных. Показаны среднее значение массы и стандартное отклонение.
На фигуре 16 представлена экспрессия CYP82E4 в течение ранних этапов воздушной обработки швейцарского Берли (Стелла). Транскрипты определяли с применением чипов Tobarray Affymetrix на основе специфичного зонда NtPMIa1g2e2_st для цитохром P450-монооксигеназы CYP82E4.
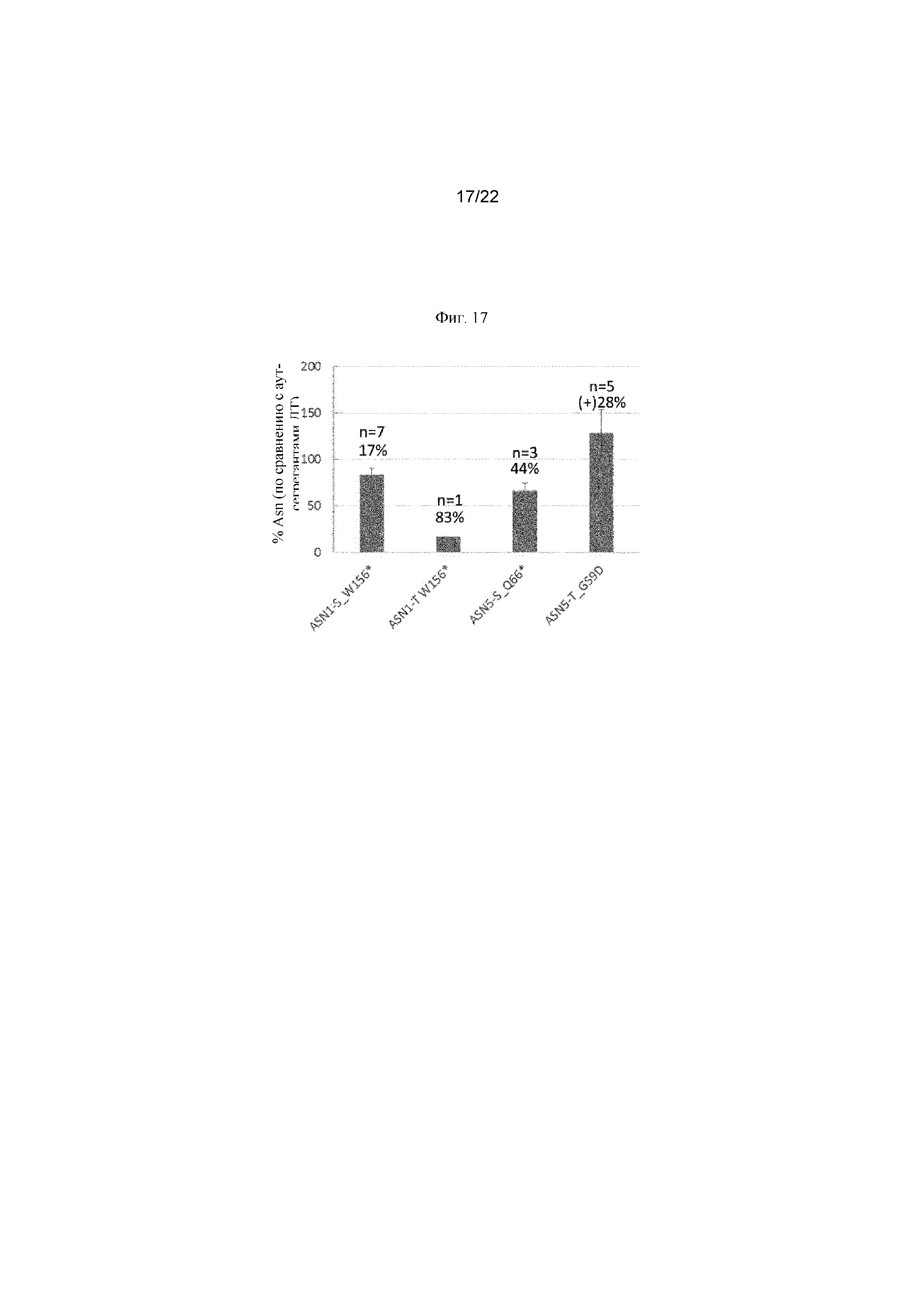
На фигуре 17 представлен процент снижения уровня аспарагина в вариантах ASN. Процентные значения рассчитывали по сравнению с аут-сегрегантами ДТ (дикого типа) для каждой линии варианта.
На фигуре 18 представлена биомасса обработанных листьев (4), отобранных в средней-нижней части стебля.
На фигуре 19 представлено определение акриламида в аэрозоле, полученном в результате нагревания изделия, образующего аэрозоль, как описано в публикации WO2013/098405, изготовленного из ASN1-RNAi, ASN5-RNAi и соответствующих контрольных листьев указанных растений. Аспарагин определяли в измельченном порошке из ASN1-RNAi, ASN5-RNAi и контрольных листьев (см. фигуру11 и фигуру13) после перемешивания с 50% табаком Вирджиния (A, C). Смесь табака преобразовывали в формованный лист и нагревали в изделии, образующем аэрозоль. Затем в аэрозоле определяли акриламид (B, D).
На фигуре 20 представлено определение акриламида в дыме горючей сигареты, изготовленной из резаного наполнителя ASN1-RNAi и соответствующего контроля листовой пластинки средней части стебля. Аспарагин (чертеж A) определяли в резаном наполнителе, полученном из ASN1-RNAi и контрольных листьев. Горючие сигареты выкуривали, и в дыме анализировали акриламид.
На фигуре 21 представлено определение акриламида в аэрозоле из изделия, генерирующего аэрозоль, изготовленного из листьев ASN1-S_стоп-мутанта и контроля (аут-сегрегант ДТ). Аспарагин определяли в измельченном порошке из ASN1-S_стоп-мутанта и контрольных листьев после перемешивания с 50% табаком Вирджиния (A). Смесь табака преобразовывали в формованный лист, и аэрозоль получали из изделия, генерирующего аэрозоль. Затем в аэрозоле определяли акриламид (B).
На фигуре 22 представлены примеры векторов, использованных для получения растений ASN-RNAi.
ОПРЕДЕЛЕНИЯ
Техническим терминам и выражениям, используемым в пределах объема данной заявки, следует придавать значение, которое обычно применяется к ним в данной области биологии растений и молекулярной биологии. Все нижеследующие определения терминов применяют ко всему содержанию данной заявки. Слово «содержащий» не исключает другие элементы или стадии, а формы единственного числа не исключают множественное. Одна стадия может выполнять функции нескольких признаков, изложенных в формуле изобретения. Термины «приблизительно», «по существу» и «приблизительно» в контексте данного цифрового значения или диапазона относятся к значению или диапазону, которые находятся в пределах 20%, в пределах 10%, или в пределах 5%, 4%, 3%, 2% или 1% от данного значения или диапазона.
Термин «выделенный» относится к любому объекту, который взят из его естественной среды, но этот термин не подразумевает какой-либо степени очистки.
«Вектор экспрессии» представляет собой средство доставки нуклеиновой кислоты, которое содержит комбинацию компонентов нуклеиновой кислоты для обеспечения экспрессии нуклеиновой кислоты. Подходящие векторы экспрессии включают в себя эписомы, способные к внехромосомной репликации, такие как кольцевые плазмиды на основе двухнитевой нуклеиновой кислоты; линеаризованные плазмиды на основе двухнитевой нуклеиновой кислоты и другие функционально эквивалентные векторы экспрессии любого происхождения. Вектор экспрессии содержит по меньшей мере промотор, расположенный выше по последовательности и функционально связанный с нуклеиновой кислотой, конструкциями нуклеиновых кислот или конъюгатом нуклеиновой кислоты, как определено ниже.
Термин «конструкция» относится к двухнитевому фрагменту рекомбинантной нуклеиновой кислоты, содержащему один или несколько полинуклеотидов. Конструкция содержит «матричную цепь», основания которой спарены с комплементарной «смысловой» или «кодирующей» цепью. Данная конструкция может быть встроена в вектор в двух возможных ориентациях, либо в той же (или смысловой) ориентации, либо в обратной (или антисмысловой) ориентации по отношению к ориентации промотора, расположенного в векторе, таком как вектор экспрессии.
«Вектор» относится к средству доставки нуклеиновой кислоты, которое содержит комбинацию компонентов нуклеиновой кислоты для обеспечения переноса нуклеиновой кислоты, конструкций нуклеиновых кислот и конъюгатов нуклеиновых кислот и тому подобного. Подходящие векторы включают в себя эписомы, способные к внехромосомной репликации, такие как кольцевые плазмиды на основе двухнитевой нуклеиновой кислоты; линеаризованные плазмиды на основе двухнитевой нуклеиновой кислоты и другие векторы любого происхождения.
«Промотор» относится к элементу/последовательности нуклеиновой кислоты, как правило, расположенным выше по цепи и функционально связанным с фрагментом двухнитевой ДНК. Промоторы могут быть получены целиком из областей вблизи нативного представляющего интерес гена, или могут состоять из разных элементов, полученных из различных нативных промоторов или сегментов синтетической ДНК.
Термины «гомология, идентичность или сходство» относятся к степени сходства последовательностей между двумя полипептидами или между двумя молекулами нуклеиновых кислот, сравниваемых путем выравнивания последовательностей. Степень гомологии между двумя отдельными сравниваемыми последовательностями нуклеиновых кислот зависит от количества идентичных или совпадающих нуклеотидов в сопоставимых положениях. Процент идентичности может быть определен путем визуального осмотра и математических расчетов. В качестве альтернативы, процент идентичности двух последовательностей нуклеиновых кислот можно определить путем сравнения информации о последовательностях с использованием компьютерной программы, такой как ClustalW, BLAST, FASTA или Smith-Waterman. Процент идентичности двух последовательностей может принимать различные значения в зависимости от: (i) метода, используемого для выравнивания последовательностей, например, ClustalW, BLAST, FASTA, Smith-Waterman (используемых в различных программах), или структурного выравнивания из 3D-сравнения; и (ii) параметров, используемых способом выравнивания, например, локального по сравнению с глобальным выравниванием, используемой матрицы замен пар (например, BLOSUM62, PAM250, Gonnet и т.д.), и штрафа за пропуск, например, функциональной формы и констант. После проведения выравнивания существуют различные способы подсчета процента идентичности между двумя последовательностями. Например, можно разделить количество идентичностей на: (i) длину самой короткой последовательности; (ii) длину выравнивания; (iii) среднюю длину последовательности; (iv) количество положений без пропуска; или (iv) количество эквивалентных положений, исключая выступающие концы. Более того, следует понимать, что процент идентичности также является строго зависимым от длины. Вследствие этого чем более короткой является пара последовательностей, тем большую идентичность последовательностей, возникшую случайно, можно ожидать. Популярная программа для множественного выравнивания ClustalW (Nucleic Acids Research (1994) 22, 4673-4680; Nucleic Acids Research (1997), 24, 4876-4882) является подходящим способом получения множественных выравниваний полипептидов или полинуклеотидов. Подходящие параметры для ClustalW могут являться следующими: Для выравниваний полинуклеотидов: Штраф за открытие пропуска=15,0, штраф за удлинение пропуска=6,66, и матрица=Identity. Для выравниваний полипептидов: Штраф за открытие пропуска=10,0, штраф за удлинение пропуска=0,2, и матрица=Gonnet. Для выравниваний ДНК и белка: ENDGAP=-1 и GAPDIST=4. Специалистам в данной области техники известно, что может быть необходимо варьировать данные и другие параметры для оптимального выравнивания последовательности. Предпочтительно, затем проводят расчет процента идентичностей на основании такого выравнивания в виде (N/T), где N представляет собой количество положений, в которых в последовательностях присутствует идентичный остаток, и T представляет собой общее количество сравниваемых положений, включая пропуски, но исключая выступающие концы.
«Вариант» означает по существу аналогичную последовательность. Вариант может обладать аналогичной функцией или по существу аналогичной функцией относительно последовательности дикого типа. В случае аспарагинсинтетазы аналогичная функция представляет собой по меньшей мере приблизительно 50%, 60%, 70%, 80% или 90% от функции фермента дикого типа по преобразованию аспартата в аспарагин из в одинаковых условиях. В случае аспарагинсинтетазы по существу аналогичная функция представляет собой по меньшей мере приблизительно 90%, 95%, 96%, 97%, 98% или 99% от функции фермента дикого типа по преобразованию аспартата в аспарагин в одинаковых условиях. Варианты могут содержать одну или несколько благоприятных мутаций, которые приводят к получению фермента, характеризующегося сниженным уровнем активности аспарагинсинтетазы по сравнению с полипептидом дикого типа. Варианты могут содержать одну или несколько благоприятных мутаций, которые приводят к нокауту активности аспарагинсинтетазы указанных вариантов (то есть, 100% ингибированию и, таким образом, нефункциональному полипептиду). Иллюстративный вариант ASN1-S дикого типа представляет собой ASN1-S_W156* (SEQ ID No 19 и 20), содержащий благоприятную стоп-мутацию, которая приводит к снижению уровня аспарагина приблизительно на 17% по сравнению с полипептидом ASN1-S дикого типа. Иллюстративный вариант ASN1-T дикого типа представляет собой ASN1-T_W156*(SEQ ID No 21 и 22), содержащий благоприятную стоп-мутацию, которая приводит к снижению уровня аспарагина приблизительно на 83% по сравнению с полипептидом ASN1-S дикого типа. Иллюстративный вариант ASN5-S дикого типа представляет собой ASN5-S_Q66* (SEQ ID No 23 и 24), содержащий благоприятную стоп-мутацию, которая приводит к снижению уровня аспарагина приблизительно на 44% по сравнению с полипептидом ASN1-S дикого типа.
Термин «растение» относится к любому растению или части растения на любой стадии его жизненного цикла или развития, а также к его потомству. В одном варианте реализации растение представляет собой «растение табака», которое относится к растению, принадлежащему к роду Nicotiana. Предпочтительные виды растения табака описаны в данном документе. Предпочтительно, растение представляет собой мутантное, неприродное или трансгенное растение, в котором экспрессию одного или нескольких генов или активность одного или нескольких белков модулируют по сравнению с контрольным растением. Предпочтительно, изменение, которое делает растение мутантным, не встречающимся в природе или трансгенным растением, приводит к модуляции экспрессии одного или нескольких генов или модуляции активности одного или нескольких белков. В некоторых вариантах реализации изменение представляет собой генетическое изменение или генетическую модификацию.
«Части растения» включают клетки растения, протопласты растений, тканевые культуры клеток растений, из которых можно регенерировать целое растение, каллюсы растений, корневища растений и клетки растений, которые являются интактными в растениях или частях растений, таких как зародыши, пыльца, пыльники, семяпочки, семена, листья, цветки, стебли, ветви, плод, корни, кончики корней и т. п. Потомство, варианты и мутанты регенерированных растений также включены в объем настоящего раскрытия при условии, что они содержат введенные полинуклеотиды, описанные в данном документе. Листья растений являются в особенности предпочтительными для применения в настоящем изобретении.
«Клетка растения» относится к структурной и физиологической единице растения. Клетка растения может находиться в виде протопласта без клеточной стенки, выделенной отдельной клетки или культивируемой клетки, или может представлять собой часть более высокоорганизованной единицы, такой как без ограничения растительная ткань, орган растения или целое растение.
Термин «растительный материал» относится к любой твердой, жидкой или газообразной композиции, или их комбинации, получаемой из растения, включая биомассу, листья, стебли, корни, цветки или части цветка, плоды, пыльцу, яйцеклетки, зиготы, семена, черенки, секреты, экстракты, клеточные или тканевые культуры, или любые другие части или продукты растения. В одном варианте реализации растительный материал содержит или состоит из биомассы, стеблей, семян или листьев. В другом варианте реализации растительный материал содержит или состоит из листьев.
Термин «сорт» относится к популяции из растений, которые обладают постоянными характеристиками, отделяющими их от других растений того же вида. Сорт имеет один или несколько отличительных признаков, и дополнительно характеризуется очень небольшой общей изменчивостью индивидуумов в пределах такой разновидности. Сорт зачастую является предметом коммерческих продаж.
Термин «линия» или «селекционная линия» в этом документе обозначает группу растений, которые используют при селекции растений. Линия отличается от разновидности, поскольку демонстрирует небольшую вариабельность между особями по одному или нескольким представляющим интерес признакам, хотя может присутствовать некоторая вариабельность между особями по другим признакам.
Термин «неприродный» в этом документе описывает объект (например, полинуклеотид, генетическую мутацию, полипептид, растение и клетку растения и растительный материал), который не образован природой или не существует в природе. Такие неприродные объекты или искусственные объекты можно создать, синтезировать, произвести, модифицировать, подвергнуть вмешательству или манипуляции способами, описанными в данном документе, или которые известны из уровня техники. Такие неприродные объекты или искусственные объекты можно создать, синтезировать, произвести, модифицировать, подвергнуть вмешательству или манипуляции человеком. Таким образом, в качестве примера, неприродное растение, неприродную клетку растения или неприродный растительный материал можно создать с применением методик манипуляции с генами, например, с использованием антисмысловой РНК, интерферирующей РНК, мегануклеазы и т.п. В качестве дополнительного примера, неприродное растение, неприродную клетку растения или неприродный растительный материал можно создать с использованием интрогрессии или путем переноса одной или нескольких генетических мутаций (например, одного или нескольких полиморфизмов) от первого растения или клетки растения во второе растение или клетку растения (которые сами по себе могут быть встречающимися в природе), так что полученное растение, клетка растения или растительный материал или их потомство содержит генетическую структуру (например, геном, хромосому или ее сегмент), которая не образована естественным путем или которая не существует в природе. Полученное растение, клетка растения или растительный материал, таким образом, являются искусственными или неприродными. Соответственно, искусственные или неприродные растение или клетку растения можно создать путем модификации генетической последовательности в первом встречающемся в природе растении или клетке растения, даже если полученная генетическая последовательность встречается в природе во втором растении или клетке растения, генетический фон которых отличается от такового первого растения или клетки растения.
Термин «модулирование» может относиться к снижению, ингибированию, повышению или иному влиянию на экспрессию или активность полипептида. Этот термин может также относиться к снижению, ингибированию, повышению или иному влиянию на активность гена, кодирующего полипептид, которое может включать в себя, помимо прочего, модулирование транскрипционной активности.
Термин «снижение» или «сниженный» в этом документе относится к снижению от приблизительно 10% до приблизительно 99%, или снижению, составляющему по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99%, или по меньшей мере 100% или более от количества или активности, такой как, помимо прочего, полипептидная активность, транскрипционная активность и экспрессия белка.
Термин «ингибировать» или «ингибированный» в этом документе относится к снижению от приблизительно 98% до приблизительно 100%, или снижению, составляющему по меньшей мере 98%, по меньшей мере 99%, но в особенности 100% от количества или активности, такой как, помимо прочего, полипептидная активность, транскрипционная активность и экспрессия белка.
Трансформация клетки может быть стабильной или временной. Термин «временная трансформация» или «временно трансформированный» или варианты указанного термина означают введение одного или нескольких экзогенных полинуклеотидов в клетку при отсутствии встраивания экзогенного полинуклеотида в геном клетки-хоязина. Напротив, термин «стабильная трансформация» или «стабильно трансформированный» означает введение и интеграцию одного или нескольких экзогенных полинуклеотидов в геном клетки. Термин «стабильный трансформант» означает клетку, в которую стабильно интегрировали один или несколько экзогенных полинуклеотидов в геномную ДНК или ДНК органелл. Следует понимать, что организм или его клетка, трансформированная нуклеиновыми кислотами, конструкциями и/или векторами согласно настоящему изобретению, могут являться временно, а также стабильно трансформированными. В некоторых вариантах реализации стабильная трансформация является предпочтительной.
Термин «повышение» или «повышенный» в этом документе относится к повышению от приблизительно 5% до приблизительно 99%, или повышению, составляющему по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98%, по меньшей мере 99%, или по меньшей мере 100% или более от количества или активности, такой как, помимо прочего, полипептидная активность, транскрипционная активность и экспрессия белка.
Термин «по существу» в этом документе и при использовании в контексте количества означает, что количество составляет по меньшей мере приблизительно 10%, по меньшей мере приблизительно 9%, по меньшей мере приблизительно 8%, по меньшей мере приблизительно 7%, по меньшей мере приблизительно 6%, по меньшей мере приблизительно 5%, по меньшей мере приблизительно 4%, по меньшей мере приблизительно 3%, по меньшей мере приблизительно 2%, по меньшей мере приблизительно 1% или по меньшей мере приблизительно 0,1% от количества, с которым его сравнивают.
Термин «контроль» в контексте контрольного растения или контрольной клетки растения и т.п. означает растение или клетку растения, в которых экспрессия или активность гена или белка, представляющего интерес, не были модулированы, и поэтому данный контроль может обеспечить сравнение или эталон относительно растения или клетки растения, в которых экспрессия или активность фермента были модифицированы. Таким образом, в контексте настоящего изобретения контроль не будет содержать по меньшей мере одно генетическое изменение, которое снижает экспрессию или активность аспарагинсинтетазы. Контрольное растение или клетка растения может содержать пустой вектор. Контрольное растение или клетка растения может соответствовать растению дикого типа или клетке растения дикого типа и т.п. Во всех таких случаях исследуемое растение и контрольное растение культивируют и собирают с использованием одинаковых протоколов для целей сравнения. Изменения уровней, соотношений, активности или распределения генов или полипептидов, описанных в настоящей заявке, или изменения фенотипа растения, в особенности, снижение накопления аспарагина и/или снижение накопления акриламида, и/или повышение накопления глутамина, и/или повышение накопления аспарагиновой кислоты, и/или повышение накопления глутаминовой кислоты, можно измерять посредством сравнения растения, о котором идет речь, с контрольным растением, причем, предпочтительно, растение, о котором идет речь, и контрольное растение культивировали и/или собирали с применением одинаковых протоколов. Контрольное растение может обеспечивать эталонную точку для измерения изменений фенотипа исследуемого растения. Показатель изменений в фенотипе можно измерять в любой момент времени для растения, в том числе, во время развития растения, старения или после обработки. Показатель изменений фенотипа можно измерять у растений, выращиваемых в любых условиях, в том числе у растений, выращиваемых в вегетационной камере, теплице или в поле. Изменения фенотипа можно измерять посредством измерения содержания аспарагина и/или содержания глутамина, и/или содержания аспарагиновой кислоты, и/или содержания глутаминовой кислоты (как описано в публикации Moldoveanu (2005)), и/или содержания акриламида ( как описано в публикациях Papousek et al., 2014 и/или Onoa et al., 2003, и/или United States Environmental Protection Agency Method 8032A - Acrylamide by Gas Chromatography, Revision 1, December 1996), и/или содержание никотина (с помощью Cooperation Centre for Scientific Research Relative to Tobacco Recommended (CORESTA) Method Number 62, Determination of nicotine in tobacco and tobacco products by gas chromatographic analysis (February 2005)) можно контролировать до и/или в течение и/или после обработки или сушки с применением способов, хорошо известных в данной области техники. Анализ свободных аминокислот проводят с применением способа на основе материалов Moldoveanu (2005).
ПОДРОБНОЕ ОПИСАНИЕ
В одном варианте реализации предложен выделенный полинуклеотид, содержащий, состоящий из или состоящий по существу из полинуклеотидной последовательности, которая характеризуется идентичностью последовательности по меньшей мере 60% с любой из последовательностей, описанных в данном документе, включая любой из полинуклеотидов, представленных в перечне последовательностей. Предпочтительно, выделенный полинуклеотид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% с указанной последовательностью. Более предпочтительно, выделенный полинуклеотид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% с указанной последовательностью. Выравнивания последовательностей, описанных в данном документе, показаны в таблице 1.
В другом варианте реализации предложен выделенный полинуклеотид, содержащий, состоящий из или состоящий по существу из полинуклеотидной последовательности, которая характеризуется идентичностью последовательности по меньшей мере 60% с SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11. Предпочтительно, выделенный полинуклеотид SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11 содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 78% 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11. Более предпочтительно, выделенный полинуклеотид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% с SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11.
Более предпочтительно, выделенный полинуклеотид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% с SEQ ID NO:1.
Более предпочтительно, выделенный полинуклеотид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% с SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11.
В другом варианте реализации предложен выделенный полинуклеотид, содержащий, состоящий из или состоящий по существу из полинуклеотидной последовательности, которая характеризуется идентичностью последовательности по меньшей мере 60% с SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23, при условии, что указанный выделенный полинуклеотид содержит мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23.
Предпочтительно, выделенный полинуклеотид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 78% 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23. Более предпочтительно, выделенный полинуклеотид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% или 100% с SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23.
В другом варианте реализации предложен полинуклеотид, содержащий, состоящий из или состоящий по существу из полинуклеотида с существенной гомологией (то есть, сходством последовательности) или существенной идентичностью с SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11.
В другом варианте реализации предложен полинуклеотид, содержащий, состоящий из или состоящий по существу из полинуклеотида с существенной гомологией (то есть, сходством последовательности) или существенной идентичностью с SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23, при условии, что указанный выделенный полинуклеотид содержит мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23.
В другом варианте реализации предложены варианты полинуклеотида, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% с последовательностью SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11. Предпочтительно, предложены варианты полинуклеотида, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% с последовательностью SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11.
Предпочтительно, предложены варианты полинуклеотида, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% с последовательностью SEQ ID NO:1.
Предпочтительно, предложены варианты полинуклеотида, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% с последовательностью SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11.
В другом варианте реализации предложены варианты полинуклеотида, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% с последовательностью SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23, при условии, что указанные варианты полинуклеотида содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23. Предпочтительно, предложены варианты полинуклеотида, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% с последовательностью SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23, при условии, что указанные варианты полинуклеотида содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23. В другом варианте реализации предусмотрены фрагменты SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11 с существенной гомологией (то есть, сходством последовательности) или существенной идентичностью с указанными последовательностями, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 78%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с соответствующими фрагментами SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11.
В другом варианте реализации предложены фрагменты SEQ ID NO:1 с существенной гомологией (то есть, сходством последовательности) или существенной идентичностью с указанной последовательностью, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с соответствующим фрагментом SEQ ID NO:1.
В другом варианте реализации предложены фрагменты SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11 с существенной гомологией (то есть, сходством последовательности) или существенной идентичностью с указанными последовательностями, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 78%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с соответствующими фрагментами SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11.
Предпочтительно, предложены фрагменты вариантов полинуклеотида SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11 с существенной гомологией (то есть, сходством последовательности) или существенной идентичностью с указанными последовательностями, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% с соответствующими фрагментами SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11.
Предпочтительно, предложены фрагменты вариантов полинуклеотида SEQ ID NO:1 с существенной гомологией (то есть, сходством последовательности) или существенной идентичностью с указанной последовательностью, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% с соответствующими фрагментами SEQ ID NO:1.
Предпочтительно, предложены фрагменты вариантов полинуклеотида SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11 с существенной гомологией (то есть, сходством последовательности) или существенной идентичностью с указанными последовательностями, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% с соответствующими фрагментами SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11.
В другом варианте реализации предложены фрагменты полинуклеотида SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23 с существенной гомологией (то есть, сходством последовательности) или существенной идентичностью с указанными последовательностями, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 78%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с соответствующими фрагментами SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23, при условии, что указанные фрагменты содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23. Предпочтительно, предложены фрагменты вариантов полинуклеотида SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23 с существенной гомологией (то есть, сходством последовательности) или существенной идентичностью с указанными последовательностями, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 72%, 73%, 74%, 75%, 78%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8% или 99,9% с соответствующими фрагментами SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23, при условии, что указанные фрагменты содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23.
В другом варианте реализации предложены полинуклеотиды, содержащие достаточную или существенную степень идентичности или сходства с SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7, или SEQ ID NO:9, или SEQ ID NO:11, которые кодируют полипептид, функционирующий в качестве аспарагинсинтетазы. Предпочтительно, полинуклеотид или полинуклеотиды, описанные в данном документе, кодируют белок с активностью аспарагинсинтетазы, которая составляет по меньшей мере приблизительно 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100% или более от активности белка, изложенного в SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:12. Чтобы определить, функционирует ли полипептид в качестве аспарагинсинтетазы, можно использовать анализ, описанный в публикации Romagni & Dayan (2000) J Agric Food Chem. May;48(5):1692-6. В другом варианте реализации предложены полинуклеотиды, содержащие достаточную или существенную степень идентичности или сходства с SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23, которые кодируют полипептид, функционирующий в качестве аспарагинсинтетазы, при условии, что указанные полинуклеотиды содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23. Предпочтительно, полинуклеотид или полинуклеотиды, описанные в данном документе, кодируют белок с активностью аспарагинсинтетазы, которая составляет по меньшей мере приблизительно 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100% или более от активности белка, изложенного в SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23.
В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:1, и SEQ ID NO:3, и SEQ ID NO:5, и SEQ ID NO:7.
В некоторых вариантах реализации модулируют экспрессию полинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 90% с SEQ ID NO:1.
В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:3, и SEQ ID NO:5, и SEQ ID NO:7.
В некоторых вариантах реализациимодулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7.
В некоторых вариантах реализациимодулируют экспрессию полинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 90% с SEQ ID NO:1.
В некоторых вариантах реализациимодулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7.
В некоторых вариантах реализациимодулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:1 и SEQ ID NO:3. В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 90% с SEQ ID NO:1 и идентичностью последовательности по меньшей мере 72% с SEQ ID NO:3. В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:5 и SEQ ID NO:7. В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:1, и SEQ ID NO:3, и SEQ ID NO:5 и/или SEQ ID NO:7. В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 90% с SEQ ID NO:1 и идентичностью последовательности по меньшей мере 72% с SEQ ID NO:3, и SEQ ID NO:5, и/или SEQ ID NO:7. В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:1 и/или SEQ ID NO:3, и SEQ ID NO:5, и SEQ ID NO:7. В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 90% с SEQ ID NO:1 и/или идентичностью последовательности по меньшей мере 72% с SEQ ID NO:3, и SEQ ID NO:5 и SEQ ID NO:7.
В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:19, и SEQ ID NO:21, и SEQ ID NO:23, при условии, что указанные полинуклеотиды содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23.
В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23, при условии, что указанные полинуклеотиды содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19, или SEQ ID NO:21, или SEQ ID NO:23.
В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:19 и SEQ ID NO:21, при условии, что указанные полинуклеотиды содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19 и SEQ ID NO:21.
В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:21 и SEQ ID NO:23, при условии, что указанные полинуклеотиды содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:21 и SEQ ID NO:23.
В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:19, и SEQ ID NO:21 и/или SEQ ID NO:23, при условии, что указанные полинуклеотиды содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO:19, SEQ ID NO:21 и SEQ ID NO:23.
В некоторых вариантах реализации модулируют экспрессиюполинуклеотида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:19, и/или SEQ ID NO:21, и SEQ ID NO:23, при условии, что указанные полинуклеотиды содержат мутацию, кодирующую стоп-кодон, в положении, эквивалентном положению, представленному в SEQ ID NO: 19, SEQ ID NO:21 и SEQ ID NO:23.
Полинуклеотид, описанный в данном документе, может включать в себя полимер из нуклеотидов, который может быть немодифицированной или модифицированной дезоксирибонуклеиновой кислотой (ДНК) или рибонуклеиновой кислотой (РНК). Соответственно, полинуклеотид может быть без ограничения геномной ДНК, комплементарной ДНК (кДНК), мРНК или антисмысловой РНК или их фрагментом(ами). Кроме того, полинуклеотид может быть однонитевой или двухнитевой ДНК, ДНК, которая является смесью однонитевых и двухнитевых участков, гибридной молекулой, содержащей ДНК и РНК, или гибридной молекулой со смесью однонитевых и двухнитевых участков или их фрагментом(ами). Кроме того, полинуклеотид может содержать трехнитевые участки, содержащие ДНК, РНК или их обе, или их фрагмент(фрагменты). Полинуклеотид может содержать одно или несколько модифицированных оснований, таких как фосфоротиоаты, и может представлять собой пептидную нуклеиновую кислоту. Как правило, полинуклеотиды могут быть собраны из выделенных или клонированных фрагментов кДНК, геномной ДНК, олигонуклеотидов или отдельных нуклеотидов или комбинации вышеперечисленного. Хотя полинуклеотидные последовательности, описанные в данном документе, показаны в виде последовательностей ДНК, последовательности включают в себя соответствующие им последовательности РНК и комплементарные (например, полностью комплементарные) им последовательности ДНК или РНК, в том числе обратно комплементарные им последовательности.
Полинуклеотид, описанный в данном документе, обычно содержит фосфодиэфирные связи, хотя некоторые случаи включают полинуклеотидные аналоги, которые могут иметь альтернативные остовы, содержащие, например, фосфороамидатные, фосфоротиоатные, фосфородитиоатные или О-метилфосфороамидитные связи; и пептидные полинуклеотидные остовы и связи. Другие аналоги полинуклеотидов включают в себя полинуклеотиды с положительно заряженными остовами, неионными остовами и безрибозными остовами. Модификации рибозофосфатного остова можно делать по разным причинам, например, для повышения стабильности и периода полужизни таких молекул в физиологических средах или в качестве зондов на биочипе. Можно получать смеси природных полинуклеотидов и аналогов; в качестве альтернативы, можно получать смеси разных полинуклеотидных аналогов и смеси встречающихся в природе полинуклеотидов и аналогов.
Известно множество полинуклеотидных аналогов, включающие, например, фосфороамидатные, фосфоротиоатные, фосфородитиоатные или О-метилфосфороамидитные связи и пептидные полинуклеотидные остовы и связи. Другие аналоги полинуклеотидов включают в себя полинуклеотиды с положительно заряженными остовами, неионными остовами и безрибозными остовами. Полинуклеотиды, содержащие один или несколько карбоциклических сахаров, также включены.
Другие аналоги включают в себя пептидные полинуклеотиды, которые представляют собой пептидные полинуклеотидные аналоги. Эти остовы являются по существу неионными в нейтральных условиях в отличие от высокозаряженного фосфодиэфирного остова встречающихся в природе полинуклеотидов. Это может давать преимущества. Во-первых, пептидный полинуклеотидный остов может характеризоваться улучшенной кинетикой гибридизации. Пептидные полинуклеотиды характеризуются более значительными изменениями температуры плавления в случае несовпадающих пар оснований по сравнению с идеально совпадающими парами оснований. В случае внутреннего несовпадения ДНК и РНК, как правило, проявляют падение температуры плавления на 2-4°C. В случае неионного пептидного полинуклеотидного остова падение составляет ближе к 7-9°C. Аналогично, вследствие их неионной природы гибридизация оснований, прикрепленных к этим остовам, является относительно нечувствительной к концентрации солей. Кроме того, пептидные полинуклеотиды могут не разрушаться или разрушаться в меньшей степени клеточными ферментами и, таким образом, могут быть более стабильными.
В числе применений раскрытых полинуклеотидов и их фрагментов находится применение фрагментов в качестве зондов в анализах гибридизации нуклеиновых кислот или в качестве праймеров для применения в анализах амплификации нуклеиновых кислот. Такие фрагменты обычно содержат по меньшей мере приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 или более смежных нуклеотидов из последовательности ДНК. В других вариантах реализации фрагмент ДНК содержит по меньшей мере приблизительно 10, 15, 20, 30, 40, 50 или 60 или более смежных нуклеотидов из последовательности ДНК. Таким образом, в одном аспекте также предложен способ детектирования полинуклеотида, кодирующего белок с активностью аспарагинсинтетазы или кодирующего фермент аспарагинсинтетазу, причем указанный способ включает применение зондов или праймеров, или того и другого. Иллюстративные праймеры изложены в SEQ ID NO: 13-16. Соответственно, следующий аспект относится к олигонуклеотидному праймеру, содержащему, состоящему из или по существу состоящему из нуклеотидной последовательности с идентичностью последовательности по меньшей мере 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15 или SEQ ID NO: 16. Также раскрыты способ применения данных праймеров для детектирования полинуклеотида, кодирующего белок с активностью аспарагинсинтетазы, и варианты применения указанного белка.
Основные параметры, влияющие на выбор условий гибридизации для полинуклеотидов, и руководство для разработки подходящих условий описаны в публикации Sambrook, J., E. F. Fritsch, and T. Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.). Используя данные о генетическом коде в комбинации с аминокислотными последовательностями, описанными в данном документе, можно получить наборы вырожденных олигонуклеотидов. Такие олигонуклеотиды применимы в качестве праймеров, например, в полимеразных цепных реакциях (ПЦР), с помощью которых выделяют и амплифицируют фрагменты ДНК. В некоторых вариантах реализации вырожденные праймеры можно использовать в качестве зондов для генетических библиотек. Такие библиотеки будут включать в себя, помимо прочего, библиотеки кДНК, геномные библиотеки и даже электронные библиотеки меток экспрессируемых последовательностей или ДНК. Гомологичные последовательности, идентифицированные этим способом, будут затем использованы в качестве зондов для идентификации гомологов последовательностей, указанных в данном документе.
Кроме того, потенциальное применение имеют полинуклеотиды и олигонуклеотиды (например, праймеры или зонды), которые гибридизируются в условиях пониженной жесткости, как правило в условиях средней жесткости, и обычно в условиях высокой жесткости с полинуклеотидом(ами), описанным в данном документе. Основные параметры, влияющие на выбор условий гибридизации, и руководство для разработки подходящих условий могут с легкостью определить средние специалисты в данной области техники на основании, например, длины или состава оснований полинуклеотида. Один из путей достижения условий средней жесткости включает применение раствора для предварительной отмывки, содержащего 5x стандартный цитрат натрия, 0,5% додецилсульфата натрия, 1,0 мM этилендиаминтетрауксусной кислоты (pH 8,0), гибридизационного буфера с приблизительно 50% формамида, 6x стандартного цитрата натрия и температуры гибридизации при приблизительно 55°C (или других подобных гибридизационных растворов, таких как содержащий приблизительно 50% формамида при температуре гибридизации около 42°С) и условий отмывки при приблизительно 60°C с применением 0,5x стандартного цитрата натрия, 0,1% додецилсульфата натрия. Как правило, условия высокой жесткости определяются как условия гибридизации, как описано выше, но с отмывкой при приблизительно 68 С, с применением 0,2x стандартного цитрата натрия, 0,1% додецилсульфата натрия. SSPE (1x SSPE представляет собой 0,15 M хлорида натрия, 10 мМ фосфата натрия и 1,25 мМ этилендиаминтетрауксусной кислоты, рН 7,4) можно заменить стандартным цитратом натрия (1x стандартный цитрат натрия представляет собой 0,15 M хлорида натрия и 15 мМ цитрата натрия) в гибридизационных и отмывочных буферах; при этом отмывку выполняют в течение 15 минут после завершения гибридизации. Следует понимать, что температуру отмывки и концентрацию солей при отмывке можно регулировать по мере необходимости для достижения требуемой степени жесткости с применением основных принципов, которые управляют реакциями гибридизации и стабильностью дуплексов, как это известно специалистам в данной области техники и дополнительно описано ниже (см., например, Sambrook, J., E. F. Fritsch, and T. Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y). При гибридизации полинуклеотида с целевым полинуклеотидом с неизвестной последовательностью предполагается, что длина гибрида будет такой, как у гибридизирующегося полинуклеотида. Когда гибридизируют полинуклеотиды с известными последовательностями, длина гибрида может быть определена путем выравнивания последовательностей полинуклеотидов и идентификации участка или участков с оптимальной комплементарностью последовательности. Температура гибридизации для гибридов, которые, предположительно, составят менее чем 50 пар оснований в длину, должна быть на 5-10 C меньше, чем температура плавления гибрида, где температуру плавления определяют в соответствии со следующими уравнениями. Для гибридов, длина которых составляет менее 18 пар оснований, температура плавления (°С)=2(количество оснований А+Т)+4(количество оснований G+C). Для гибридов, длина которых составляет более 18 пар оснований, температура плавления (°C)=81,5+16,6(log10 [Na+])+0,41(% G+C)-(600/N), где N - количество оснований в гибриде и [Na+] - концентрация ионов натрия в гибридизационном буфере ([Na+] для 1x стандартного цитрата натрия=0,165 M). Как правило, каждый такой гибридизирующийся полинуклеотид имеет длину, которая составляет по меньшей мере 25% (обычно, по меньшей мере 50%, 60% или 70%, и наиболее часто по меньшей мере 80%) от длины полинуклеотида, с которым он гибридизируется, и характеризуется по меньшей мере 60% идентичностью последовательности (например, по меньшей мере 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100%) с полинуклеотидом, с которым он гибридизируется.
Как будет понятно специалисту в данной области техники, линейная ДНК имеет две возможные ориентации: направление 5'-3' и направление 3'-5'. Например, если эталонная последовательность расположена в направлении 5'-3', и если вторая последовательность расположена в направлении 5'-3' в той же полинуклеотидной молекуле/нити, то эталонная последовательность и вторая последовательность ориентированы в одном направлении или имеют одинаковую ориентацию. Как правило, промоторная последовательность и представляющий интерес ген, находящийся под контролем данного промотора, расположены в одинаковой ориентации. Однако если по отношению к эталонной последовательности, расположенной в направлении 5'-3', вторая последовательность расположена в направлении 3'-5' в той же полинуклеотидной молекуле/нити, тогда эталонная последовательность и вторая последовательность ориентированы в антисмысловом направлении или имеют антисмысловую ориентацию. Две последовательности, имеющие антисмысловые ориентации по отношению друг к другу, могут быть альтернативно описаны как имеющие одинаковую ориентацию, если эталонная последовательность (направление 5'-3') и обратно комплементарная последовательность эталонной последовательности (эталонной последовательности, расположенной в 5'-3') расположены в пределах одной полинуклеотидной молекулы/нити. Последовательности, изложенные в данном документе, показаны в направлении 5'-3'.
Рекомбинантные конструкции, предложенные в данном документе, могут быть использованы для трансформации растений или клеток растений для модулирования уровней экспрессии и/или активности белка. Рекомбинантная полинуклеотидная конструкция может содержать полинуклеотид, кодирующий один или несколько полинуклеотидов, описанных в данном документе, функционально связанных с регуляторным участком, подходящим для экспрессии полипептида. Таким образом, полинуклеотид может содержать кодирующую последовательность, которая кодирует полипептид, описанный в данном документе. Растения или клетки растений, в которых модулируют уровни экспрессии и/или активности белка, могут включать в себя мутантные, неприродные, трансгенные, созданные человеком или полученные с помощью методик генной инженерии растения или клетки растений. Предпочтительно, растение или клетка растения содержит геном, который был изменен путем стабильной интеграции рекомбинантной ДНК. Рекомбинантная ДНК включает ДНК, полученную с помощью методик генной инженерии и сконструирована вне клетки, и включает ДНК, содержащую встречающуюся в природе ДНК, или кДНК, или синтетическую ДНК. Растение может включать в себя растение, регенерированное из первоначально трансформированной клетки растения, и растения-потомки от более поздних поколений или скрещиваний трансформированного растения. Предпочтительно, модификация изменяет экспрессию или активность полинуклеотида или полипептида, описанного в данном документе, по сравнению с контрольным растением.
Полипептид, кодируемый рекомбинантным полинуклеотидом, может быть нативным полипептидом или может быть гетерологичным по отношению к клетке. В некоторых случаях рекомбинантная конструкция содержит полинуклеотид, который модулирует экспрессию, функционально связанный с регуляторным участком. Примеры подходящих регуляторных участков описаны в данном документе.
Также предложены векторы, содержащие рекомбинантные полинуклеотидные конструкции, такие как описанные в данном документе. Подходящие основы для векторов включают в себя, например, те, которые обычно используют в данной области техники, такие как плазмиды, вирусы, искусственные хромосомы, искусственные хромосомы бактерий, искусственные хромосомы дрожжей или искусственные хромосомы бактериофагов. Подходящие векторы экспрессии включают в себя без ограничения плазмиды и вирусные векторы, полученные из, например, бактериофага, бакуловирусов и ретровирусов. Многочисленные векторы и системы экспрессии являются коммерчески доступными. Векторы могут включать в себя, например, точки начала репликации, участки связывания с ядерным матриксом или маркеры. Маркерный ген может придавать клетке растения селектируемый фенотип. Например, маркер может придавать биоцидную устойчивость, такую как устойчивость к антибиотику (например, к канамицину, G418, блеомицину или гигромицину) или к гербициду (например, к глифосату, хлорсульфурону или фосфинотрицину). Кроме того, вектор экспрессии может включать в себя последовательность метки, предназначенную для облегчения манипуляций или выявления (например, очистки или локализации) экспрессированного полипептида. Последовательности меток, такие как последовательности люциферазы, бета-глюкуронидазы, зеленого флуоресцентного белка, глутатион S-трансферазы, полигистидина, c-myc или гемагглютинина, как правило, экспрессируются в виде слияния с кодируемым полипептидом. Такие метки могут быть вставлены в любом месте в пределах полипептида, в том числе на карбоксильном или амино-конце.
Растение или клетку растения можно трансформировать путем интегрирования рекомбинантного полинуклеотида в ее геном, чтобы они стали стабильно трансформированными. Растение или клетка растения, описанные в данном документе, могут быть стабильно трансформированными. Стабильно трансформированные клетки, как правило, сохраняют введенный полинуклеотид при каждом клеточном делении. Растение или клетка растения также могут быть временно трансформированными, так что рекомбинантный полинуклеотид не интегрируется в их геном. Как правило, временно трансформированные клетки утрачивают весь введенный рекомбинантный полинуклеотид или некоторую его часть при каждом клеточном делении, так что введенный рекомбинантный полинуклеотид невозможно выявить в дочерних клетках после прохождения достаточного количества клеточных делений. Также рассматривается использование редактирования генома.
В данной области техники существует целый ряд способов, доступных для трансформации клетки растения, каждый из которых охвачен данным документом, в том числе, биолистика, методики с применением генной пушки, Agrobacterium-опосредованная трансформация, опосредованная вирусным вектором трансформация и электропорация. Система Agrobacterium для интеграции чужеродной ДНК в хромосомы растений была тщательно изучена, модифицирована и использована для генной инженерии растений. Молекулы депротеинизированной рекомбинантной ДНК, содержащие последовательности ДНК, соответствующие исследуемому очищенному белку, функционально связанные в смысловой или антисмысловой ориентации с регуляторными последовательностями, соединяют с соответствующими последовательностями Т-ДНК с помощью общепринятых способов. Данные молекулы вводят в протопласты при помощи методики на основе полиэтиленгликоля или методики электропорации, каждая из которых является стандартной. В качестве альтернативы, такие векторы, содержащие молекулы рекомбинантной ДНК, кодирующие исследуемый очищенный белок, вводят в живые клетки Agrobacterium, которые затем переносят ДНК в клетки растения. Трансформацию с помощью депротеинизированной ДНК без сопутствующих векторных последовательностей Т-ДНК можно выполнять посредством слияния протопластов с ДНК-содержащими липосомами или посредством электропорации. Депротеинизированную ДНК без сопутствующих векторных последовательностей Т-ДНК также можно использовать для трансформации клеток с помощью инертных высокоскоростных микрочастиц.
Если клетку или культивируемую ткань используют в качестве реципиентной ткани для трансформации, растения можно регенерировать из трансформированных культур, при необходимости, с помощью методик, известных специалистам в данной области техники.
Выбор регуляторных участков, которые следует включить в рекомбинантную конструкцию, зависит от нескольких факторов, в том числе без ограничения от эффективности, селектируемости, индуцируемости, необходимого уровня экспрессии и предпочтительной экспрессии в определенных клетках или тканях. Обычным делом для специалиста в данной области техники является модулирование экспрессии кодирующей последовательности путем соответствующего выбора и размещения регуляторных участков по отношению к кодирующей последовательности. Аналогичным образом можно модулировать транскрипцию полинуклеотида. Некоторые подходящие регуляторные участки инициируют транскрипцию исключительно или преимущественно в определенных типах клеток. Способы идентификации и определения свойств регуляторных участков в геномной ДНК растений хорошо известны из уровня техники.
Примеры промоторов включают в себя тканеспецифичные промоторы, распознаваемые тканеспецифичными факторами, присутствующими в разных типах тканей или клеток (например, специфичные для корня промоторы, специфичные для побега промоторы, специфичные для ксилемы промоторы), или присутствующими на различных стадиях развития, или присутствующими в ответ на различные условия окружающей среды. Примеры промоторов включают в себя конститутивные промоторы, которые могут активироваться в большинстве типов клеток без необходимости в специфических индукторах. Примеры промоторов для контроля над образованием полипептида РНКи включают в себя промотор 35S вируса мозаики цветной капусты (CaMV/35S), SSU, OCS, lib4, usp, STLS1, B33, nos или промоторы убиквитина или фазеолина. Специалисты в данной области техники могут создавать множество вариантов рекомбинантных промоторов.
Тканеспецифичные промоторы представляют собой элементы управления транскрипцией, которые активны только в определенных клетках или тканях в определенное время в ходе развития растений, например, в вегетативных тканях или репродуктивных тканях. Тканеспецифичная экспрессия может иметь преимущество, например, когда предпочтительной является экспрессия полинуклеотидов в определенных тканях. Примеры тканеспецифичных промоторов под управлением развития включают в себя промоторы, которые могут инициировать транскрипцию только (или в основном только) в определенных тканях, таких как вегетативные ткани, например,корни или листья, или репродуктивные ткани, такие как плоды, семяпочки, семена, пыльца, тычинки, цветы или любая эмбриональная ткань. Промоторы, специфичные для репродуктивных тканей, могут быть, например, специфичными для пыльника, специфичными для семяпочки, специфичными для зародыша, специфичными для эндосперма, специфичными для интегумента, специфичными для семян и семенной кожуры, специфичными для пыльцы, специфичными для лепестка, специфичными для чашелистика или их комбинациями.
Примеры специфичных для листа промоторов включают в себя промотор гена пируватортофосфатдикиназы (PPDK) из C4-растения (маиса), промотор cab-m1Ca+2 из маиса, промотор родственных myb генов из Arabidopsis thaliana (Atmyb5), промоторы генов рибулозобифосфаткарбоксилазы (RBCS) (например, генов томата RBCS 1, RBCS2 и RBCS3A, экспрессируемых в листьях и выращенных на свету саженцах, RBCS1 и RBCS2, экспрессируемых в развивающихся плодах томата, или промотор гена рибулозобифосфаткарбоксилазы, экспрессируемого на высоких уровнях почти исключительно в клетках мезофилла листовых пластинок и листовых пазух).
Примеры специфичных для старения промоторов включают в себя промотор из томата, активный во время созревания плодов, старения и опадения листьев, промотор гена, кодирующего цистеиновую протеазу маиса, промотор гена 82E4 и промотор генов SAG.
В некоторых вариантах реализации промотор, специфичный для старения, является предпочтительным. Предпочтительно, промотор может представлять собой промотор E4. В публикации Plant Mol Biol. (2008) Mar;66(4):415-27 сообщалось, что промотор E4 является подходящим промотором, специфичным для старения, для табака с применением GUS в качестве репортерного гена. Другой иллюстративный промотор представляет собой промотор SAG12, описанный в публикации Plant Cell. (1999) Jun;11(6):1073-80.
Промоторы, специфичные к пыльнику, представляют собой следующие примеры. Можно выбрать промоторы для экспрессии предпочтительно в корнях, известные специалистам в данной области техники. Промоторы для экспрессии предпочтительно в семенах включают в себя как специфичные для семян промоторы (промоторы, активные в процессе развития семян, такие как промоторы запасных белков семян), так и промоторы прорастающих семян (промоторы, активные во время прорастания семян). Такие промоторы для экспрессии предпочтительно в семенах включают в себя без ограничения промотор гена Cim1 (цитокинин-индуцированный сигнал); cZ19B1 (19 кДa зеин маиса); milps (миоинозитол-1-фосфатсинтаза); mZE40-2, также известный как Zm-40; nuclc и celA (целлюлозосинтаза). Промотор гена гамма-зеина представляет собой специфичный для эндосперма промотор. Промотор гена Glob-1 представляет собой специфичный для зародыша промотор. В случае двудольных растений специфичные для семян промоторы включают в себя без ограничения промотор гена бета-фазеолина фасоли, гена напина, гена β-конглицинина, гена лектина сои, гена круциферина и т. п. В случае однодольных растений специфичные для семян промоторы включают в себя без ограничения промотор гена 15 кДа зеина маиса, промотор гена 22 кДа зеина, промотор гена 27 кДа зеина, промотор гена g-зеина, промотор гена 27 кДа гамма-зеина (такой как промотор gzw64A, см. номер доступа S78780 в Genbank), промотор гена waxy, промотор гена shrunken 1, промотор гена shrunken 2, промотор гена глобулина 1 (см. номер доступа L22344 в Genbank), промотор гена Itp2, промотор гена cim1, промоторы генов end1 и end2 маиса, промотор гена nuc1, промотор гена Zm40, промотор гена eep1 и eep2; промотор гена lec1, промотор гена тиоредоксина H; промотор гена mlip15, промотор гена PCNA2 и промотор гена shrunken-2.
Примеры индуцируемых промоторов включают в себя промоторы, реагирующие на нападение патогенов, анаэробные условия, повышенную температуру, свет, засуху, низкую температуру или высокую концентрацию солей. Индуцируемые патогеном промоторы включают в себя промоторы связанных с патогенезом белков (PR-белки), которые индуцируются после заражения патогеном (например, PR-белки, SAR-белки, бета-1,3-глюканаза, хитиназа).
В дополнение к растительным промоторам можно получить другие подходящие промоторы бактериального происхождения (например, промотор гена октопинсинтазы, промотор гена нопалинсинтазы и другие промоторы, полученные из Ti-плазмид), или их можно получить из вирусных промоторов (например, 35S и 19S РНК промоторы вируса мозаики цветной капусты (CaMV), конститутивные промоторы вируса табачной мозаики, промоторы 19S и 35S вируса мозаики цветной капусты (CaMV) или промотор 35S вируса мозаики норичника). В некоторых вариантах реализации промотор вируса мозаики мирабелис (Mirabilis Mosaic Virus, MMV) является предпочтительным. В некоторых вариантах реализации промотор 35S является предпочтительным.
В другом аспекте предложен выделенный полипептид, содержащий, состоящий из или по существу состоящий из полипептидной последовательности, которая характеризуется идентичностью последовательности по меньшей мере 60% с любой из полипептидных последовательностей, описанных в данном документе, включая любые полипептиды, представленные в перечне последовательностей. Предпочтительно, выделенный полипептид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 76%, 77%, 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с указанными последовательностями.
В одном варианте реализации выделенный полипептид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 76%, 77%, 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:12. В другом варианте реализации выделенный полипептид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:12.
В одном варианте реализации выделенный полипептид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 76%, 77%, 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24.
В другом варианте реализации выделенный полипептид содержит, состоит из или состоит по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24.В другом варианте реализации предложен вариант полипептида, содержащий, состоящий из или по существу состоящий из аминокислотной последовательности, кодируемой вариантом полинуклеотида с идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 76%, 77%, 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% с SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:12.
В другом варианте реализации предложен вариант полипептида, содержащий, состоящий из или по существу состоящий из аминокислотной последовательности, кодируемой вариантом полинуклеотида с идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 76%, 77%, 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% с SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24. В другом варианте реализации предложены фрагменты полипептида SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:12или фрагменты SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:12, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 76%, 77%, 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с соответствующими фрагментами SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:12.
В другом варианте реализации предложены фрагменты полипептида SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24 или фрагменты SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24, которые характеризуются идентичностью последовательности по меньшей мере приблизительно 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 75%, 76%, 77%, 78%, 79%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% с соответствующими фрагментами SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24. В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8. В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2, и SEQ ID NO:4, и SEQ ID NO:6, и SEQ ID NO:8. В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2 и SEQ ID NO:4. В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:6 и SEQ ID NO:8. В некоторых вариантах реализации активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2, и SEQ ID NO:4, и SEQ ID NO:6, и/или SEQ ID NO:8. В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2, и/или SEQ ID NO:4, и SEQ ID NO:6, и SEQ ID NO:8.
В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:20, или SEQ ID NO:22, или SEQ ID NO:24.
В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:20, и SEQ ID NO:22, и SEQ ID NO:24, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:20, и SEQ ID NO:22, и SEQ ID NO:24.
В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:20 и SEQ ID NO:22, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:20 и SEQ ID NO:22.
В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:22 и SEQ ID NO:24, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:22 и SEQ ID NO:24.
В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:20 и SEQ ID NO:24, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:20 и SEQ ID NO:24.
В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:20, и SEQ ID NO:22, и/или SEQ ID NO:24, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:20, и SEQ ID NO:22, и/или SEQ ID NO:24.
В некоторых вариантах реализации модулируют активность полипептида, содержащего, состоящего из или по существу состоящего из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:20, и/или SEQ ID NO:22, и SEQ ID NO:24, при условии, что указанный полипептид содержит стоп-кодон мутацию в положении, эквивалентном положению, представленному в SEQ ID NO:20, и/или SEQ ID NO:22, и SEQ ID NO:24.Полипептид может содержать фрагменты последовательностей, которые характеризуются достаточной или существенной степенью идентичности или сходства для функционирования в качестве аспарагинсинтетазы. Фрагменты полипептида или полипептидов, как правило, сохраняют некоторую степень активности или полную активность полноразмерной последовательности - например, по меньшей мере приблизительно 95%, 96%, 97%, 98%, 99%, 99,1%, 99,2%, 99,3%, 99,4%, 99,5%, 99,6%, 99,7%, 99,8%, 99,9% или 100% активности.
Как рассматривается в данном документе, полипептиды также включают в себя мутантов, полученных за счет введения изменений любого типа (например, вставок, делеций или замен аминокислот, изменений в сайтах гликозилирования, изменений, которые влияют на рефолдинг или изомеризацию, трехмерные структуры или состояния самоассоциации), при условии, что указанные мутанты все еще сохраняют некоторую степень или полную свою функцию или активность в качестве аспарагинсинтетазы. Предпочтительно, функцию или активность в качестве аспарагинсинтетазы модулируют, снижают или ингибируют. Предпочтительно, функцию или активность в качестве аспарагинсинтетазы ингибируют так, что активность аспарагинсинтетазы становится необнаруживаемой.
Полипептиды включают в себя варианты, полученные путем введения изменений любого типа (например, вставки, делеции или замены аминокислот, изменения структуры гликозилирования, изменения, которые влияют на рефолдинг или изомеризацию, трехмерную структуру или состояния самоассоциации), которые могут быть сконструированы преднамеренно или выделены естественным образом. Вариант может содержать изменения, которые обеспечивают молчащее изменение и приводят к образованию функционально эквивалентного белка. Преднамеренные аминокислотные замены можно осуществлять, исходя из сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и амфипатической природы остатков при условии, что сохраняется вторичная активность связывания вещества. Например, отрицательно заряженные аминокислоты включают в себя аспарагиновую кислоту и глутаминовую кислоту, положительно заряженные аминокислоты включают в себя лизин и аргинин, и аминокислоты с незаряженными полярными концевыми группами с подобными значениями гидрофильности включают в себя лейцин, изолейцин, валин, глицин, аланин, аспарагин, глутамин, серин, треонин, фенилаланин и тирозин. Консервативные замены можно осуществлять, например, в соответствии с приведенной ниже таблицей. Аминокислоты в одном и том же блоке во втором столбце и предпочтительно в одной и той же строке в третьем столбце можно заменять одна на другую:
Полипептид может быть зрелым белком или незрелым белком, или белком, полученным из незрелого белка. Полипептиды могут быть в линейной форме или циклизированы с использованием известных способов. Полипептиды, как правило, содержат по меньшей мере 10, по меньшей мере 20, по меньшей мере 30 или по меньшей мере 40, или по меньшей мере 50, или по меньшей мере 100, или по меньшей мере 200, или по меньшей мере 300, или по меньшей мере 400, или по меньшей мере 500, или по меньшей мере 600 непрерывных аминокислот.
Также раскрыт полипептид, кодируемый SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:12, или SEQ ID NO: 20, или SEQ ID NO:22, или SEQ ID NO:24, который характеризуется идентичностью последовательности 100% с указанными последовательностями, или полипептид, содержащий, состоящий из или состоящий по существу из последовательности, представленной в SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8, или SEQ ID NO:10, или SEQ ID NO:12, или SEQ ID NO: 20, или SEQ ID NO:22, или SEQ ID NO:24.
Полипептид можно получить путем культивирования трансформированных или рекомбинантных клеток-хозяев в условиях культивирования, подходящих для экспрессии полипептида. Полученный экспрессированный полипептид затем можно очистить из такой культуры с помощью известных способов очистки. Очистка полипептида может включать в себя использование аффинной колонки, содержащей средства, которые будут связываться с полипептидом; одну или несколько стадий с использованием колонки с пропусканием через такие аффинные смолы; одну или несколько стадий с участием хроматографии с гидрофобным взаимодействием или иммуноаффинную хроматографию. В качестве альтернативы, полипептид может быть экспрессирован в форме, которая будет облегчать очистку. Например, он может быть экспрессирован в виде полипептида слияния, например, с мальтозосвязывающим полипептидом, глутатион-5-трансферазой, гистидиновой меткой или тиоредоксином. Наборы для экспрессии и очистки полипептидов слияния являются коммерчески доступными. Полипептид можно пометить эпитопом и затем очищать с применением специфического антитела к такому эпитопу. Одну или несколько стадий жидкостной хроматографии, такой как обращенно-фазовая высокоэффективная жидкостная хроматография, можно использовать для дополнительной очистки полипептида. Некоторые или все из вышеуказанных стадий очистки в различных комбинациях можно использовать для получения практически однородного рекомбинантного полипептида. Очищенный таким образом полипептид может практически не содержать другие полипептиды и определен в данном документе как «практически очищенный полипептид»; такие очищенные полипептиды включают полипептиды, фрагменты, варианты и т. п. Экспрессию, выделение и очистку полипептидов и фрагментов можно выполнять при помощи любой подходящей методики, в том числе, помимо прочего, способами, описанными в данном документе.
Кроме того, можно использовать аффинную колонку, например, с моноклональным антителом против полипептидов, для аффинной очистки экспрессированных полипептидов. Эти полипептиды можно удалять из аффинной колонки с помощью общепринятых методик, например, в элюирующем буфере с высоким содержанием солей с последующим диализом в буфере с низким содержанием солей для использования, или посредством изменения рН или других компонентов в зависимости от используемой аффинной матрицы, или путем конкурентного удаления с помощью естественного субстрата аффинного фрагмента.
Раскрыты выделенные или практически очищенные полинуклеотиды или композиции белков. «Выделенный» или «очищенный» полинуклеотид или белок, или его биологически активная часть практически или, по сути, не содержит компоненты, которые обычно сопровождают полинуклеотид или белок или взаимодействуют с ними при нахождении в его естественном окружении. Таким образом, выделенный или очищенный полинуклеотид или белок по существу не содержит другой клеточный материал или культуральную среду при получении с помощью методик рекомбинации, или по существу не содержит химических соединений-предшественников или других химических соединений при получении путем химического синтеза. В оптимальном случае «выделенный» полинуклеотид не содержит последовательностей (оптимально, белок-кодирующих последовательностей), которые в естественных условиях фланкируют полинуклеотид (например, последовательностей, расположенных на 5'- и 3'-концах полинуклеотида) в геномной ДНК организма, из которого получен полинуклеотид. Например, в различных вариантах реализации выделенный полинуклеотид может содержать менее приблизительно 5 т. о., 4 т. о., 3 т. о., 2 т. о., 1 т. о., 0,5 т. о. или 0,1 т. о. нуклеотидной последовательности, которая в естественных условиях фланкирует полинуклеотид в геномной ДНК клетки, из которой получен полинуклеотид. Белок, который практически не содержит клеточный материал, включает препараты белка, характеризующегося содержанием контаминирующего белка, составляющим менее приблизительно 30%, 20%, 10%, 5% или 1% (из расчета на сухую массу).
Полипептид можно также получать с помощью известных общепринятых методик химического синтеза. Способы конструирования полипептидов или их фрагментов с помощью способов синтеза известны специалистам в данной области техники. Синтетически сконструированные полипептидные последовательности благодаря первичной, вторичной или третичной структуре или конформационным характеристикам, которые являются общими с нативными полипептидами, могут обладать общими с ними биологическими свойствами, включая биологическую активность.
Различия в генетическом фоне можно обнаружить по фенотипическим различиям или с помощью методик молекулярной биологии, известных в данной области техники, таких как секвенирование нуклеиновых кислот, определение наличия или отсутствия генетических маркеров (например, микросателлитных маркеров РНК).
Также предложены антитела, которые являются иммунореактивными в отношении полипептидов, описанных в данном документе. Полипептиды, фрагменты, варианты, полипептиды слияния и им подобные, изложенные в данном документе, можно использовать в качестве «иммуногенов» в получении антител, иммунореактивных по отношению к ним. Такие антитела могут специфически связываться с полипептидом посредством антигенсвязывающих сайтов антитела. Специфически связывающимися антителами являются такие антитела, которые будут специфически распознавать полипептид, гомологи и варианты и связываться с ними, а не с другими молекулами. В одном варианте реализации антитела являются специфичными в отношении полипептидов, характеризующихся аминокислотной последовательностью, изложенной в данном документе, и не взаимодействуют перекрестно с другими полипептидами.
Более конкретно, полипептиды, фрагмент, варианты, полипептиды слияния и им подобные содержат антигенные детерминанты или эпитопы, которые вызывают образование антител. Данные антигенные детерминанты или эпитопы могут быть линейными или конформационными (прерывистыми). Линейные эпитопы состоят из одного аминокислотного участка полипептида, тогда как конформационные или прерывистые эпитопы состоят из аминокислотных участков из различных областей полипептидной цепи, которые приводятся в непосредственную близость при сворачивании полипептида. Эпитопы можно идентифицировать любыми способами, известными из уровня техники. Кроме того, эпитопы из полипептидов можно использовать в качестве реагентов для исследования, в анализах и для очистки специфически связывающихся антител из веществ, таких как поликлональные сыворотки или супернатанты от культивируемых гибридом. Такие эпитопы или их варианты можно получать с использованием методик, известных в данной области техники, например, путем твердофазного синтеза, химического или ферментативного расщепления полипептида, или с помощью технологии рекомбинантной ДНК.
Как поликлональные, так и моноклональные антитела к полипептидам можно получать с помощью общепринятых методик. Клеточные линии гибридом, которые вырабатывают моноклональные антитела, специфичные в отношении полипептидов, также рассматриваются в данном документе. Такие гибридомы можно получать и идентифицировать с помощью общеизвестных методик. Для получения антител различных животных-хозяев можно иммунизировать путем введения полипептида, его фрагмента, варианта или мутантов. К примеру, такие животные-хозяева могут включать, помимо прочего, кроликов, мышей и крыс. Для усиления иммунологического ответа можно использовать различные адъюванты. В зависимости от вида хозяина такие адъюванты включают, помимо прочего, адъюванты Фрейнда (полный и неполный), минеральные гели, такие как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, полиолы плюроников, полианионы, пептиды, масляные эмульсии, гемоцианин фисуреллы, динитрофенол и потенциально пригодные в качестве адъювантов для человека BCG (бацилла Кальметта-Герена) и Corynebacterium parvum. Моноклональные антитела можно выделять с помощью общепринятых методик. Такие моноклональные антитела могут быть иммуноглобулинами любого класса, в том числе IgG, IgM, IgE, IgA, IgD, и любого их подкласса.
Антитела можно также использовать в анализах для выявления присутствия полипетидов или фрагментов либо in vitro, либо in vivo. Антитела можно также использовать для очистки полипептидов или фрагментов с помощью иммуноаффинной хроматографии.
Также раскрыты фрагменты полинуклеотидов и полипептидов, кодируемых указанными полинуклеотидами. Фрагменты полинуклеотида могут кодировать фрагменты белка, которые сохраняют биологическую активность нативного белка. В качестве альтернативы, фрагменты полинуклеотида, применимые в качестве зондов для гибридизации или ПЦР-праймеров, как правило, не кодируют фрагмент белка, сохраняющий биологическую активность. Кроме того, фрагменты раскрытых нуклеотидных последовательностей включают в себя такие фрагменты, которые могут быть собраны в рекомбинантные конструкции, как описано в данном документе. Фрагменты полинуклеотидной последовательности могут варьировать от по меньшей мере приблизительно 25 нуклеотидов, приблизительно 50 нуклеотидов, приблизительно 75 нуклеотидов, приблизительно 100 нуклеотидов, приблизительно 150 нуклеотидов, приблизительно 200 нуклеотидов, приблизительно 250 нуклеотидов, приблизительно 300 нуклеотидов, приблизительно 400 нуклеотидов, приблизительно 500 нуклеотидов, приблизительно 600 нуклеотидов, приблизительно 700 нуклеотидов, приблизительно 800 нуклеотидов, приблизительно 900 нуклеотидов, приблизительно 1000 нуклеотидов, приблизительно 1100 нуклеотидов, приблизительно 1200 нуклеотидов, приблизительно 1300 нуклеотидов или приблизительно 1500 нуклеотидов, приблизительно 2000 нуклеотидов, приблизительно 3000 нуклеотидов, приблизительно 4000 нуклеотидов, приблизительно 5000 нуклеотидов, приблизительно 6000 нуклеотидов, приблизительно 7000 нуклеотидов, приблизительно 8000 нуклеотидов, приблизительно 9000 нуклеотидов,
приблизительно 10000 нуклеотидов, приблизительно 15000 нуклеотидов, приблизительно 20000 нуклеотидов и до полноразмерного полинуклеотида, кодирующего полипептиды, описанные в данном документе. Фрагменты полипептидной последовательности могут варьировать от по меньшей мере приблизительно 25 аминокислот, приблизительно 50 аминокислот, приблизительно 75 аминокислот, приблизительно 100 аминокислот приблизительно 150 аминокислот, приблизительно 200 аминокислот, приблизительно 250 аминокислот, приблизительно 300 аминокислот, приблизительно 400 аминокислот, приблизительно 500 аминокислот, приблизительно 600 аминокислот и до полноразмерного полипептида, описанного в данном документе.
Следует понимать, что снижения экспрессии или активности аспарагинсинтетазы или аспарагинсинтетаз, описанных в настоящей заявке, можно достичь различными способами. В некоторых вариантах реализации снижение экспрессии аспарагинсинтетазы или аспарагинсинтетаз можно осуществить на геномном уровне и/или на уровне транскрипта с применением множества молекул, которые препятствуют транскрипции и/или трансляции, включая, без ограничения, молекулы антисмысловой, миРНК, рибозима или дезоксирибозима. Также можно использовать встраивание одной или нескольких мутаций в по меньшей мере один ген, включая делеции, вставки, сайт-специфичные мутации, нуклеазы «цинкового пальца» и т.п. Согласно другим вариантам реализации экспрессию можно заингибировать на уровне белка с применением антагонистов или ферментов, которые расщепляют полипептид, и т.п.
В одном аспекте описано мутантное растение или часть указанного растения, содержащего по меньшей мере одну мутацию и (i) полинуклеотид, содержащий, состоящий из или состоящий по существу из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7; (ii) полипептид, кодируемый полинуклеотидом, описанным в (i); (iii) полипептид, содержащий, состоящий из или состоящий по существу из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8; или (iv) конструкцию, вектор или вектор экспрессии, содержащий выделенный полинуклеотид, изложенный в (i), причем по меньшей мере одна мутация снижает экспрессию или активность аспарагинсинтетазы по сравнению с контрольным растением, которое не содержит по меньшей мере одну мутацию. В другом аспекте описано мутантное растение или часть указанного растения, содержащего по меньшей мере одну мутацию и (i) полинуклеотид, содержащий, состоящий из или состоящий по существу из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 90% с SEQ ID NO:1 или идентичностью последовательности по меньшей мере 72% с SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7; (ii) полипептид, кодируемый полинуклеотидом, описанным в (i); (iii) полипептид, содержащий, состоящий из или состоящий по существу из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8; или (iv) конструкцию, вектор или вектор экспрессии, содержащий выделенный полинуклеотид, изложенный в (i), причем по меньшей мере одна мутация снижает экспрессию или активность аспарагинсинтетазы по сравнению с контрольным растением, которое не содержит по меньшей мере одну мутацию.
Растение или клетка растения вследствие этого может содержать одну или несколько мутаций в NtASN1-S (SEQ ID NO: 1) и/или NtASN1-T (SEQ ID NO: 3), и/или NtASN3-S (SEQ ID NO: 9), и/или NtASN3-T (SEQ ID NO: 11), и/или NtASN5-S (SEQ ID NO: 5), и/или NtASN5-T (SEQ ID NO: 7), причем указанная мутация приводит к снижению экспрессии или снижению функции указанного гена или белка, кодируемого указанным геном. Растение или клетка растения может содержать одну или несколько мутаций в NtASN1-S (SEQ ID NO: 1) или NtASN1-T (SEQ ID NO: 3), или NtASN5-S (SEQ ID NO: 5), или NtASN5-T (SEQ ID NO: 7), как описано, причем указанная мутация приводит к снижению экспрессии или снижению функции указанного гена или белка, кодируемого указанным геном. Растение или клетка растения может содержать одну или несколько мутаций (например, нонсенс-мутации или т.п., как описано в данном документе) в NtASN1-S (SEQ ID NO: 1) и NtASN1-T (SEQ ID NO: 3) или NtASN5-S (SEQ ID NO: 5) и NtASN5-T (SEQ ID NO: 7), как описано, причем указанная мутация приводит к снижению экспрессии или снижению функции указанного гена или белка, кодируемого указанным геном. Экспрессию или функцию мутанта или мутантов можно модулировать, ингибировать или снижать. Помимо одной или нескольких мутаций, описанных в данном документе, мутантное растение или клетка растения могут содержать одну или несколько дополнительных мутаций в одном или нескольких других генах или полипептидах. В некоторых вариантах реализации мутанты могут содержать одну или несколько дополнительных мутаций в одном или нескольких других генах или полипептидах. В данном документе описаны мутанты NtASN1-S, NtASN1-T, NtASN5-S и ASN5-T. В некоторых вариантах реализации одна или несколько мутаций находятся в домене глутаминазы.
Три стоп-мутации идентифицированы в копиях ASN1-S (ASN1-S_W156* - SEQ ID No 19 и 20), ASN1-T (ASN1-T_W156* - SEQ ID No 21 и 22) и ASN5-S (ASN5-S_Q66* - SEQ ID No 23 и 24), но не в копии ASN5-T. Поскольку в ASN5-T не были идентифицированы стоп-мутации, мутация, отличная от стоп-мутации, была идентифицирована в качестве потенциального кандидата. Наблюдалась однозначная тенденция к снижению уровня аспарагина в ASN1-S_W156*, ASN1-T_W156* и ASN5-S_Q66* по сравнению с их соответствующими сегрегантами ДТ, но не для ASN5-T_G59D. Наибольшее снижение наблюдалось для ASN1-T_W156* (83%), затем для ASN5-S_Q66*(44%) и, наконец, для ASN1-S_W156* (17%). В линии ASN5-T_G59D наблюдалось незначительное повышение уровня аспарагина. Стоп-мутации в ASN1-S и ASN5-S не оказывали влияния на биомассу и размер растений. Согласно публикации Canales et al., 2012, мутации во всех генах Nt-ASN расположены в домене глутаминазы, которая высвобождает аммоний из глутамина, домене синтетазы, расположенном в Nt-ASN от аминокислоты R211 до D451 консенсусной области.
Указанные мутантное растение или клетка растения могут являться гетерозиготными или гомозиготными по мутации или мутациям. Указанные мутантное растение или клетка растения могут являться гетерозиготными по меньшей мере по одной мутации и гомозиготными по меньшей мере по одной отличающейся мутации. Предпочтительно, мутантное растение или клетка растения являются гомозиготными по мутации или мутациям.
Мутант может содержать мутацию (например, нонсенс-мутацию, которая приводит к появлению стоп-кодона) в положении, эквивалентном положению аминокислоты 156 SEQ ID NO: 2. Пример такого мутанта представляет собой ASN1-S_W156*, как описано в данном документе.
Мутант может содержать мутацию (например, нонсенс-мутацию, которая приводит к появлению стоп-кодона) в положении, эквивалентном положению аминокислоты 156 SEQ ID NO: 4. Пример такого мутанта представляет собой ASN1-T_W156*, как описано в данном документе.
Мутант может содержать мутацию (например, нонсенс-мутацию, которая приводит к появлению стоп-кодона) в положении, эквивалентном положению аминокислоты 66 SEQ ID NO: 6. Пример такого мутанта представляет собой ASN5-S_Q66*, как описано в данном документе.
Также предусмотрены комбинации данных мутаций. Указанные комбинации могут включать в себя комбинацию ASN1-S_W156* и ASN1-T_W156*; или комбинацию ASN1-S_W156* и ASN5-S_Q66*; или комбинацию ASN1-T_W156* и ASN5-S_Q66*; или комбинацию ASN1-S_W156*, ASN1-T_W156* и ASN5-S_Q66*.
В другом аспекте предложен способ снижения уровня аспарагина в растении или в растительном материале, полученном из растения, причем указанный способ включает введение в геном указанного растения одной или нескольких мутаций, которые снижают экспрессию по меньшей мере одного гена аспарагинсинтетазы, причем указанный по меньшей мере один ген аспарагинсинтетазы кодирует NtASN1-S, NtASN1-T, NtASN5-S или Nt ASN5-T. Предпочтительно, помимо мутации или мутаций в NtASN1-S, NtASN1-T, NtASN5-S или NtASN5-T, одна или несколько мутаций также могут быть введены в по меньшей мере один аллель по меньшей мере одного, двух или трех или более дополнительных генов аспарагинсинтетазы. Также предусмотрены мутации в NtASN1-S и NtASN1-T или NtASN5-S и NtASN5-T.
Также предложен способ идентификации растения со сниженными уровнями аспарагина, причем указанный способ включает осуществление скрининга образца нуклеиновой кислоты из растения, представляющего интерес, для детектирования присутствия одной или нескольких мутаций в NtASN1-S, NtASN1-T, NtASN5-S или ASN5-T. Предпочтительно, указанный способ также включает осуществление скрининга указанного образца нуклеиновой кислоты или другого образца нуклеиновой кислоты от указанного растения, представляющего интерес, для детектирования присутствия мутации в NtASN1-S, присутствия мутации в NtASN1-T, или присутствия мутации в NtASN5-S, или присутствия мутации в NtASN5-T.
Также раскрыты растение или клетка растения, которые являются гетерозиготными или гомозиготными по одной или нескольким мутациям в гене, кодирующем NtASN1-S, NtASN1-T, NtASN5-S или NtASN5-T, причем указанная мутация или мутации приводят к снижению экспрессии гена или снижению функции белка, кодируемого указанным геном.
В некоторых вариантах реализации благоприятную мутацию или мутации вводят в растение или клетку растения с использованием подхода мутагенеза, и введенную мутацию идентифицируют или подвергают отбору с применением способов, известных специалисту в данной области техники, таких как анализ методом Саузерн-блоттинга, секвенирование ДНК, ПЦР-анализ или фенотипический анализ. Мутации, которые влияют на экспрессию генов или которые нарушают функцию кодируемого белка, можно определять с применением способов, хорошо известных из уровня техники. Инсерционные мутации в экзонах гена, как правило, приводят к образованию нефункциональных мутантов. Мутации в консервативных остатках могут являться в особенности эффективными для ингибирования или снижения метаболической функции кодируемого белка.
Также раскрыты способы получения мутантных полинуклеотидов и полипептидов. Любое представляющее интерес растение, в том числе клетку растения или растительный материал, можно генетически модифицировать различными способами, с помощью которых, как известно, индуцируется мутагенез, в том числе сайт-направленный мутагенез, олигонуклеотид-направленный мутагенез, индуцированный химическими соединениями мутагенез, индуцированный ионизирующим излучением мутагенез, мутагенез с использованием модифицированных оснований, мутагенез с использованием содержащих пропуски ДНК-дуплексов, мутагенез с двухнитевыми разрывами, мутагенез с использованием штаммов хозяев с нарушенной репарацией, мутагенез посредством синтеза полного гена, ДНК-шаффлинг и другие эквивалентные способы.
Мутантные варианты полипептида можно использовать для создания мутантных, не встречающихся в природе или трансгенных растений (например, мутантных, не встречающихся в природе, трансгенных, созданных человеком растений или растений, полученных при помощи методик генной инженерии) или клеток растения, содержащих один или несколько мутантных вариантов полипептида. Предпочтительно мутантные варианты полипептида сохраняют активность немутировавшего полипептида. Активность мутантного варианта полипептида может быть выше, ниже или приблизительно такой же, как у немутировавшего полипептида.
Мутации в нуклеотидных последовательностях и полипептидах, описанных в данном документе, могут включать в себя внесенные человеком мутации или синтетические мутации, или мутации, созданные при помощи генной инженерии. Мутации в нуклеотидных последовательностях и полипептидах, описанных в данном документе, могут представлять собой мутации, которые получены или которые можно получить посредством способа, который включает этап манипуляции in vitro или in vivo. Мутации в нуклеотидных последовательностях и полипептидах, описанных в данном документе, могут представлять собой мутации, которые получены или которые можно получить посредством способа, который включает вмешательство человека. В качестве примера, способ может включать в себя мутагенез с применением экзогенно добавляемых химических соединений, таких как мутагенные, тератогенные или канцерогенные органические соединения, например, этилметансульфонат (ЭМС), которые приводят к случайным мутациям в генетическом материале. В качестве дополнительного примера способ может включать один или несколько этапов осуществления генной инженерии, таких как один или несколько этапов осуществления генной инженерии, которые описаны в данном документе, или их комбинации. В качестве дополнительного примера способ может включать в себя одну или несколько стадий скрещивания растений.
Активность одного или нескольких полипептидов аспарагинсинтетазы в растении снижается или ингибируется согласно настоящему изобретению, если активность по превращению является статистически меньшей, чем активность по превращению того же полипептида или полипептидов аспарагинсинтетазы в растении, которое не было модифицировано с целью ингибирования активности по превращению данного полипептида аспарагинсинтетазы, и которое культивировали и собирали с использованием тех же протоколов. Активность полипептида аспарагинсинтетазы в растении считается устраненной, если данная активность является необнаруживаемой с помощью аналитических способов, описанных в данном документе. Способы определения активности полипептида аспарагинсинтетазы описаны в данном документе.
Помимо мутагенеза, композиции, которые могут модулировать экспрессию или активность одного или нескольких полинуклеотидов или полипептидов, описанных в данном документе, включают в себя, без ограничения, специфичные в отношении последовательности полинуклеотиды, которые могут препятствовать транскрипции одного или нескольких эндогенных генов; специфичные в отношении последовательности полинуклеотиды, которые могут препятствовать трансляции РНК-транскриптов (например, двухнитевые РНК, миРНК, рибозимы); специфичные в отношении последовательности полипептиды, которые могут препятствовать стабильности одного или нескольких белков; специфичные в отношении последовательности полинуклеотиды, которые могут препятствовать ферментативной активности одного или нескольких белков или активности связывания одного или нескольких белков с субстратами или регуляторными белками; антитела, которые проявляют специфичность в отношении одного или нескольких белков; низкомолекулярные соединения, которые могут препятствовать стабильности одного или нескольких белков, или ферментативной активности одного или нескольких белков, или активности связывания одного или нескольких белков; белки «цинковые пальцы», которые связывают один или несколько полинуклеотидов; и мегануклеазы, которые характеризуются активностью в отношении одного или нескольких полинуклеотидов. Технологии редактирования гена, генетические технологии редактирования и технологии редактирование генома хорошо известны в данной области техники.
Один из способов редактирования гена предусматривает использование подобных активатору транскрипции эффекторных нуклеаз (TALEN), вызывающих двухнитевые разрывы, на которые клетка может отвечать с привлечением механизмов репарации. Негомологичное соединение концов вновь соединяет ДНК по обеим сторонам двухнитевого разрыва, если имеется незначительное перекрывание или вовсе отсутствует перекрывание последовательности для комплементарного связывания. Этот механизм репарации индуцирует ошибки в геноме посредством вставки или делеции, либо хромосомной перестройки. Любые такие ошибки могут превратить генные продукты, закодированные в этом месте, в нефункциональные. Другой способ редактирования гена предусматривает использование бактериальной системы CRISPR/Cas. Бактерии и архебактерии характеризуются хромосомными элементами, называемыми короткими палиндромными повторами, регулярно расположенными группами (CRISPR), которые являются частью адаптивной иммунной системы, защищающей от проникновения вирусной и плазмидной ДНК. В системах CRISPR II типа, CRISPR РНК (crRNA) функционирует с трансактивирующей crRNA (tracrRNA) и CRISPR-ассоциированными (Cas) белками с внесением двухнитевых разрывов в целевую ДНК. Целевое расщепление с помощью Cas9 требует спаривания оснований crRNA и tracrRNA, а также спаривания оснований crRNA и целевой ДНК. Целевое распознавание облегчается при наличии короткого мотива, называемого прилегающим к протоспейсеру мотивом (PAM), который соответствует последовательности NGG. Данную систему можно приспособить для редактирования генома. Cas9 обычно программируется двойной РНК, состоящей из crRNA и tracrRNA. Тем не менее, коровые компоненты этих РНК могут быть объединены в единую гибридную «направляющую РНК» для нацеливания Cas9. Использование некодирующей направляющей РНК для нацеливания на ДНК для сайт-специфического расщепления в перспективе является значительно более простым, чем существующие технологии, такие как TALEN. Использование стратегии CRISPR/Cas с перенацеливанием нуклеазного комплекса требует только введения новой последовательности РНК, и нет необходимости в переконструировании специфичности факторов транскрипции белка.
Технология применения антисмысловых олигонуклеотидов представляет собой другой хорошо известный способ, который можно использовать для модулирования экспрессии полипептида. Полинуклеотид гена, который подлежит репрессии, клонируют и функционально связывают с регуляторным участком и последовательностью терминации транскрипции, так что антисмысловая нить РНК транскрибируется. Затем рекомбинантной конструкцией трансформируют клетку растения и получают антисмысловую нить РНК. Полинуклеотид не обязательно является полной последовательностью гена, который подлежит репрессии, но, как правило, будет по существу комплементарен по меньшей мере части смысловой нити гена, который подлежит репрессии.
Полинуклеотид может быть транскрибирован в рибозим или каталитическую РНК, которые влияют на экспрессию мРНК. Рибозимы можно сконструировать для специфического спаривания практически с любой целевой РНК и расщепления фосфодиэфирного остова в определенном месте, тем самым функционально инактивируя целевую РНК. Гетерологичные полинуклеотиды могут кодировать рибозимы, сконструированные для расщепления конкретных мРНК-транскриптов, предотвращая таким образом экспрессию полипептида. Рибозимы типа ʺголовка молоткаʺ применимы для разрушения конкретных мРНК, хотя можно использовать различные рибозимы, которые расщепляют мРНК в последовательностях сайт-специфического распознавания. Рибозимы типа ʺголовка молоткаʺ расщепляют мРНК в местах, определяемых фланкирующими участками, которые формируют комплементарные пары оснований с целевой мРНК. Единственным требованием является то, что целевая РНК должна содержать нуклеотидную последовательность 5'-UG-3'. Конструирование и получение рибозимов типа ʺголовка молоткаʺ известно из уровня техники. Последовательности рибозимов типа ʺголовка молоткаʺ можно встроить в стабильную РНК, такую как транспортная РНК (тРНК) для повышения эффективности расщепления in vivo.
В одном варианте реализации специфичный к последовательности полинуклеотид, который может препятствовать трансляции РНК-транскрипта(ов), представляет собой интерферирующую РНК. РНК-интерференция или РНК-сайленсинг представляет собой эволюционно консервативный процесс, с помощью которого определенные мРНК могут намечаться для ферментативной деградации. Двухнитевую РНК (двухнитевая РНК) вводят в клетку или она образуется в клетке (например, вирус с двухнитевой РНК или полинуклеотиды интерферирующей РНК) для инициации пути РНК-интерференции. Двухцепочечную РНК можно преобразовать в несколько двойных спиралей малых интерферирующих РНК 21-23 п.о. в длину с помощью РНКазы III, которая представляет собой специфичную для двухцепочечной РНК эндонуклеазу. Затем малые интерферирующие РНК могут распознаваться индуцируемыми РНК комплексами сайленсинга, которые способствуют раскручиванию малых интерферирующих РНК с помощью АТФ-зависимого процесса. Раскрученная антисмысловая нить малой интерферирующей РНК направляет активированные индуцируемые РНК комплексы сайленсинга к целевой мРНК, содержащей последовательность, комплементарную антисмысловой нити малой интерферирующей РНК. Целевая мРНК и антисмысловая нить могут образовывать А-форму спирали, и большая бороздка А-формы спирали может распознаваться активированными индуцируемыми РНК комплексами сайленсинга. Целевая мРНК может расщепляться активированными индуцируемыми РНК комплексами сайленсинга в одном сайте, который определяется сайтом связывания 5'-конца нити малой интерферирующей РНК. Активированные индуцируемые РНК комплексы сайленсинга могут повторно использоваться для катализа еще одного события расщепления. Пример кодирующей последовательности из NtASN1-S, которую можно использовать для сайленсинга копий как NtASN1-S, так и NtASN1-T, представлен в SEQ ID NO: 17. Вследствие этого следующий аспект относится к нуклеотидной последовательности, представленной в SEQ ID NO: 17, или последовательности, которая характеризуется идентичностью последовательности по меньшей мере 72%, 73%, 74%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% с указанной последовательностью. Пример кодирующей последовательности из NtASN5-T, которую можно использовать для сайленсинга копий как NtASN1-S, так и NtASN1-T, представлен в SEQ ID NO: 18. Вследствие этого следующий аспект относится к нуклеотидной последовательности, представленной в SEQ ID NO: 17. Вследствие этого следующий аспект относится к нуклеотидной последовательности, представленной в SEQ ID NO: 18, или последовательности, которая характеризуется идентичностью последовательности по меньшей мере 72%, 73%, 74%, 75%, 80%, 85%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95% 96%, 97%, 98%, 99% с указанной последовательностью. Также предусмотрены способ сайленсинга генов с применением данных кодирующих последовательностей и варианты применения указанного способа.
Векторы экспрессии интерферирующей РНК могут содержать конструкции интерферирующей РНК, кодирующие полинуклеотиды интерферирующей РНК, которые проявляют активность РНК-интерференции при помощи снижения уровня экспрессии мРНК, пре-мРНК или родственных вариантов РНК. Иллюстративные конструкции представлены на фигуре 22. Векторы экспрессии могут содержать промотор, расположенный выше по цепи и функционально связанный с конструкцией интерферирующей РНК, как дополнительно описано в данном документе. Векторы экспрессии интерферирующей РНК могут содержать подходящий минимальный коровый промотор, представляющую интерес конструкцию интерферирующей РНК, расположенный выше по цепи (5') регуляторный участок, расположенный ниже по цепи (3') регуляторный участок, в том числе сигналы терминации транскрипции и полиаденилирования, и другие последовательности, известные специалистам в данной области техники, такие как различные селективные маркеры.
Полинуклеотиды можно получить в различных формах, в том числе в виде двухнитевых структур (то есть, двухнитевая молекула РНК, содержащая антисмысловую нить и комплементарную ей смысловую нить), двухнитевых шпилькообразных структур или однонитевых структур (то есть, молекула ssRNA, содержащая только антисмысловую нить). Структуры могут содержать вторичную структуру в виде двойной спирали, асимметричной двойной спирали, шпильки или асимметричной шпильки, имеющую самокомплементарную смысловую и антисмысловую нити. Двухнитевую интерферирующую РНК можно превратить ферментативным путем в двухнитевые малые интерферирующие РНК. Одну из нитей двойной спирали малой интерферирующей РНК можно отжечь с комплементарной последовательностью в целевой мРНК и родственных вариантах РНК. Двойные спирали малой интерферирующей РНК/мРНК распознаются РНК-индуцированными комплексами сайленсинга, которые могут расщеплять РНК в нескольких сайтах зависимым от последовательности образом, приводя в результате к разрушению целевой мРНК и родственных вариантов РНК.
Молекулы двухнитевой РНК могут включать в себя молекулы малой интерферирующей РНК, собранные из одного олигонуклеотида в структуре стебель-петля, где самокомплементарные смысловой и антисмысловой участки молекулы малой интерферирующей РНК соединены с помощью основанного на полинуклеотидах или не основанного на полинуклеотидах линкера (линкеров), а также кольцевую однонитевую РНК с двумя или более петлевыми структурами и стеблем, содержащую самокомплементарные смысловую и антисмысловую нити, где кольцевую РНК можно процессировать либо in vivo, либо in vitro с образованием активной молекулы малой интерферирующей РНК, способной опосредовать интерференции РНК.
Также предполагается использование молекул малой РНК, образующей шпильки. Они содержат специфичную антисмысловую последовательность в дополнение к обратно комплементарной (смысловой) последовательности, как правило, отделенной спейсером или последовательностью петли. Расщепление спейсера или петли обеспечивает молекулу однонитевой РНК и ее обратно комплементарную нить, так что их можно отжечь с образованием двухнитевой молекулы РНК (необязательно, с дополнительными стадиями обработки, которые могут привести в результате к добавлению или удалению одного, двух, трех или более нуклеотидов 3'-конца или 5'-конца одной или обеих нитей). Спейсер может иметь достаточную длину для обеспечения возможности комплементарного связывания антисмысловой и смысловой последовательностей и образования двухнитевой структуры (или стебля) до расщепления спейсера (и, необязательно, последующих стадий обработки, которые могут привести в результате к добавлению или удалению одного, двух, трех, четырех или более нуклеотидов с 3'-конца или с 5'-конца одной или обеих нитей). Спейсерная последовательность представляет собой, как правило, неродственную нуклеотидную последовательность, которая находится между двух комплементарных участков нуклеотидной последовательности, которая при комплементарном связывании в двухнитевой полинуклеотид содержит малую шпилечную РНК. Спейсерная последовательность обычно содержит от приблизительно 3 до приблизительно 100 нуклеотидов.
Любой представляющий интерес полинуклеотид РНК можно получить путем подбора подходящей композиции последовательности, размера петли и длины стебля для получения двойной спирали шпильки. Подходящий диапазон для конструирования длины стебля двойной спирали шпильки включает длину стебля по меньшей мере в приблизительно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 нуклеотидов, например, приблизительно 14-30 нуклеотидов, приблизительно 30-50 нуклеотидов, приблизительно 50-100 нуклеотидов, приблизительно 100-150 нуклеотидов, приблизительно 150-200 нуклеотидов, приблизительно 200-300 нуклеотидов, приблизительно 300-400 нуклеотидов, приблизительно 400-500 нуклеотидов, приблизительно 500-600 нуклеотидов и приблизительно 600-700 нуклеотидов. Подходящий диапазон для конструирования длины петли двойной спирали шпильки включает длину петли в приблизительно 4-25 нуклеотидов, приблизительно 25-50 нуклеотидов или длиннее, если длина стебля двойной спирали шпильки является значительной. В некоторых вариантах реализации молекула двухнитевой РНК или ssRNA составляет от приблизительно 15 до приблизительно 40 нуклеотидов в длину. В другом варианте реализации молекула малой интерферирующей РНК представляет собой молекулу двухнитевой РНК или ssRNA от приблизительно 15 до приблизительно 35 нуклеотидов в длину. В другом варианте реализации молекула малой интерферирующей РНК представляет собой молекулу двухнитевой РНК или ssRNA от приблизительно 17 до приблизительно 30 нуклеотидов в длину. В другом варианте реализации молекула малой интерферирующей РНК представляет собой молекулу двухнитевой РНК или ssRNA от приблизительно 19 до приблизительно 25 нуклеотидов в длину. В другом варианте реализации молекула малой интерферирующей РНК представляет собой молекулу двухцепочечной РНК или ssРНК от приблизительно 21 до приблизительно 23 нуклеотидов в длину. В некоторых вариантах реализации структуры в виде шпильки с участками двойной спирали длиннее, чем 21 нуклеотид, могут способствовать эффективному сайленсингу, управляемому малой интерферирующей РНК, вне зависимости от последовательности и длины петли. Примеры кодирующих последовательностей, используемых для РНК-интерференции, изложены в SEQ ID NO: 17 и 18.
Целевая последовательность мРНК составляет, как правило, от приблизительно 14 до приблизительно 50 нуклеотидов в длину. Целевую мРНК можно, таким образом, проверить на наличие участков от приблизительно 14 до приблизительно 50 нуклеотидов в длину, которые предпочтительно соответствуют одному или более из следующих критериев для целевой последовательности: соотношение A+T/G+С составляет от приблизительно 2:1 до приблизительно 1:2; динуклеотид AA или динуклеотид СA на 5'-конце целевой последовательности; последовательность из по меньшей мере 10 смежных нуклеотидов, уникальных для целевой мРНК (то есть, последовательность отсутствует в других последовательностях мРНК из этого же растения); и нет «полос» из более чем трех смежных гуаниновых (G) нуклеотидов или более чем трех смежных цитозиновых (C) нуклеотидов. Данные критерии можно оценивать с помощью различных методик, известных в данной области техники, например, компьютерные программы, такие как BLAST, можно использовать для поиска по общедоступным базам данных, чтобы определить, является ли выбранная целевая последовательность уникальной для целевой мРНК. В качестве альтернативы, можно выбрать целевую последовательность (и сконструировать последовательность малой интерферирующей РНК) с помощью коммерчески доступного компьютерного программного обеспечения (например, OligoEngine, Target Finder и Design Tool для малых интерферирующих РНК, которые являются коммерчески доступными).
В одном варианте реализации выбраны целевые последовательности мРНК, которые составляют от приблизительно 14 до приблизительно 30 нуклеотидов в длину и соответствуют одному или более критериям, указанным выше. В другом варианте реализации выбраны целевые последовательности, которые составляют от приблизительно 16 до приблизительно 30 нуклеотидов в длину и соответствуют одному или нескольким критериям, указанным выше. В дополнительном варианте реализации выбраны целевые последовательности, которые составляют от приблизительно 19 до приблизительно 30 нуклеотидов в длину и соответствуют одному или нескольким критериям, указанным выше. В другом варианте реализации выбраны целевые последовательности, которые составляют от приблизительно 19 до приблизительно 25 нуклеотидов в длину и соответствуют одному или нескольким критериям, указанным выше.
В иллюстративном варианте реализации молекулы малой интерферирующей РНК содержат специфическую антисмысловую последовательность, которая комплементарна по меньшей мере 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 или более смежным нуклеотидам из любой из описанных в данном документе полинуклеотидных последовательностей.
Специфическая антисмысловая последовательность, содержащаяся в молекуле малой интерферирующей РНК, может быть идентичной или практически идентичной комплементарной цепи целевой последовательности. В одном варианте реализации специфическая антисмысловая последовательность, содержащаяся в молекуле малой интерферирующей РНК, является по меньшей мере приблизительно на 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичной комплементарной цепи последовательности целевой мРНК. Способы определения идентичности последовательности известны в данной области техники, и ее можно определить, например, с помощью программы BLASTN программного обеспечения Computer Group (GCG) Университета штата Висконсин, или предоставленной на веб-сайте NCBI.
Специфическая антисмысловая последовательность молекул малой интерферирующей РНК может проявлять изменчивость в отношении различий (например, нуклеотидной замены, в том числе, транзиции или трансверсии) по одному, двум, трем, четырем или более нуклеотидам последовательности целевой мРНК. Если такие нуклеотидные замены находятся в антисмысловой цепи молекулы двухнитевой РНК, то комплементарный нуклеотид в смысловой цепи, с которым замещенный нуклеотид, как правило, должен образовывать водородные связи при спаривании оснований, может быть или может не быть соответственно замещенным. Молекулы двухнитевой РНК, в которых одна или несколько нуклеотидных замен происходят в смысловой последовательности, но не в антисмысловой нити, также предусмотрены. Если антисмысловая последовательность молекулы малой интерферирующей РНК содержит одно или несколько несовпадений между нуклеотидной последовательностью малой интерферирующей РНК и целевой нуклеотидной последовательностью, как описано выше, эти несоответствия могут быть обнаружены на 3'-конце, 5'-конце, или в центральной части антисмысловой последовательности.
В другом варианте реализации молекулы малой интерферирующей РНК содержат специфическую антисмысловую последовательность, которая способна селективно гибридизироваться в жестких условиях с частью встречающегося в природе целевого гена или целевой мРНК. Как известно специалистам в данной области техники, вариации жесткости условий гибридизации могут быть достигнуты путем изменения времени, температуры или концентрации растворов, используемых для стадий гибридизации и отмывания. Подходящие условия могут также частично зависеть от конкретных используемых нуклеотидных последовательностей, например, последовательности целевой мРНК или гена.
Одним из способов индукции сайленсинга двухнитевой РНК в растениях является трансформация генной конструкцией, производящей РНК, образующую шпильки (см. Smith et al. (2000) Nature, 407, 319-320). Такие конструкции содержат инвертированные участки последовательности целевого гена, отделенные соответствующим спейсером. Вставка функционального интронного участка растения в качестве спейсерного фрагмента дополнительно повышает эффективность индукции сайленсинга гена за счет выработки РНК, образующей шпильки, сплайсированной с интроном (Wesley et al. (2001) Plant J., 27, 581-590). Предпочтительно длина стебля составляет от приблизительно 50 нуклеотидов до приблизительно 1 тысячи нуклеотидов в длину. Способы получения РНК, образующей шпильки, сплайсированной с интроном, хорошо описаны в данной области техники (см., например, Bioscience, Biotechnology, and Biochemistry (2008) 72, 2, 615-617).
Молекулы интерферирующей РНК, имеющие двуспиральную или двухнитевую структуру, например, двухнитевая РНК или малая РНК, образующая шпильки, могут содержать тупые концы, или могут содержать липкие 3'- или 5'-концы. Используемый в данном документе термин «липкий конец» относится к неспаренному нуклеотиду или нуклеотидам, которые выступают из двухспиральной структуры, если 3'-конец одной нити РНК выходит за пределы 5'-конца другой нити (липкий 3'-конец), или наоборот (липкий 5'-конец). Нуклеотиды, представляющие собой липкие концы, могут быть рибонуклеотидами, дезоксирибонуклеотидами или их модифицированными версиями. В одном варианте реализации по меньшей мере одна нить молекулы интерферирующей РНК имеет липкий 3'-конец от приблизительно 1 до приблизительно 6 нуклеотидов в длину. В других вариантах реализации липкий 3'-конец составляет от приблизительно 1 до приблизительно 5 нуклеотидов, от приблизительно 1 до приблизительно 3 нуклеотидов и от приблизительно 2 до приблизительно 4 нуклеотидов в длину.
Если молекула интерферирующей РНК содержит липкий 3'-конец на одном конце молекулы, другой конец может быть с тупым концом, или также иметь липкий конец (5' или 3'). Если молекула интерферирующей РНК содержит липкий конец с обоих концов молекулы, длина липких концов может быть одинаковой или разной. В одном варианте реализации молекула интерферирующей РНК содержит липкие 3'-концы от приблизительно 1 до приблизительно 3 нуклеотидов в длину на обоих концах молекулы. В дополнительном варианте реализации молекула интерферирующей РНК представляет собой двухнитевую РНК, имеющую липкий 3'-конец из 2 нуклеотидов с обоих концов молекулы. В еще одном варианте реализации нуклеотиды, представляющие собой липкий конец интерферирующей РНК, являются динуклеотидами TT или динуклеотидами UU.
При определении процентной идентичности молекулы интерферирующей РНК, содержащей один или несколько липких концов, с последовательностью целевой мРНК, липкий конец(концы) может учитываться или не учитываться. Например, нуклеотиды с липкого 3'-конца и до 2 нуклеотидов с 5'- или 3'-конца двойной цепи могут быть модифицированы без существенной потери активности молекулы малой интерферирующей РНК.
Молекулы интерферирующей РНК могут содержать одну или несколько 5' или 3'-кэп-структур. Молекула интерферирующей РНК может содержать кэп-структуру на 3'-конце смысловой цепи, антисмысловой цепи или как смысловой, так и антисмысловой цепей; или на 5'-конце смысловой цепи, антисмысловой цепи или как смысловой, так и антисмысловой цепей молекулы интерферирующей РНК. В качестве альтернативы, молекула интерферирующей РНК может содержать кэп-структуру как на 3'-конце, так и на 5'-конце молекулы интерферирующей РНК. Термин «кэп-структура» относится к химической модификации, включенной с любого конца олигонуклеотида, которая защищает молекулу от разрушения эндонуклеазами и может также облегчать доставку или локализацию внутри клетки.
Другой модификацией, применяемой к молекулам интерферирующей РНК, является химическое связывание с молекулой интерферирующей РНК одного или нескольких фрагментов или конъюгатов, которые повышают активность, клеточное распределение, клеточный захват, биодоступность или стабильность молекулы интерферирующей РНК. Полинуклеотиды можно синтезировать или модифицировать с помощью способов, общепринятых в данной области техники. Химические модификации могут включать без ограничения 2'-модификации, введение неприродных оснований, ковалентное присоединение лиганда и замещение фосфатных связей тиофосфатными связями. В данном варианте реализации целостность двухспиральной структуры усиливают с помощью по меньшей мере одной и, как правило, двух химических связей. Химическое связывание может обеспечиваться с помощью любого из множества хорошо известных методик, например, путем введения ковалентных, ионных или водородных связей; гидрофобных взаимодействий, Ван-дер-Ваальсовых или стекинговых взаимодействий; посредством координации ионами металлов или благодаря использованию аналогов пуринов.
Нуклеотиды одной или обеих из двух одинарных цепей можно модифицировать для модулирования активации клеточных ферментов, таких как, например, без ограничения определенные нуклеазы. Методики для снижения или ингибирования активации клеточных ферментов известны в данной области техники и включают без ограничения 2'-аминомодификации, 2'-фтормодификации, 2'-алкилмодификации, модификации незаряженного каркаса, морфолиновые модификации, 2'-О-метилмодификации и фосфорамидат. Таким образом, по меньшей мере одну 2'-гидроксильную группу нуклеотидов двухнитевой РНК замещают химической группой. Также, по меньшей мере один нуклеотид можно модифицировать с образованием закрытого нуклеотида. Такой закрытый нуклеотид содержит метиленовый или этиленовый мостик, который соединяет 2'-кислород рибозы с 4'-углеродом рибозы. Введение закрытого нуклеотида в олигонуклеотид улучшает аффинность к комплементарным последовательностям и повышает температуру плавления на несколько градусов.
Можно конъюгировать лиганды с молекулой интерферирующей РНК, например, для повышения в отношении нее клеточной абсорбции. В некоторых вариантах реализации гидрофобный лиганд конъюгируют с молекулой для облегчения прямого проникновения через клеточную мембрану. Данные подходы использованы для облегчения проникновения в клетку антисмысловых олигонуклеотидов. В определенных случаях конъюгация катионного лиганда с олигонуклеотидами часто приводит к улучшенной устойчивости к нуклеазам. Типичные примеры катионных лигандов включают пропиламмоний и диметилпропиламоний. Антисмысловые олигонуклеотиды могут сохранять свою высокую аффинность связывания с мРНК, когда катионный лиганд распределен по всему олигонуклеотиду.
Молекулы и полинуклеотиды, описанные в данном документе, можно получить с помощью хорошо известных методик твердофазного синтеза. Любые другие средства для такого синтеза, известные в данной области техники, можно использовать дополнительно или в качестве альтернативы.
Различные варианты реализации направлены на векторы экспрессии, содержащие один или несколько полинуклеотидов или одну или несколько конструкций интерферирующей РНК, описанных в данном документе. Иллюстративные конструкции представлены на фигуре 21.
Различные варианты реализации направлены на векторы экспрессии, содержащие один или несколько полинуклеотидов или одну или более конструкций интерферирующей РНК, кодирующих один или несколько полинуклеотидов интерферирующей РНК, описанных в данном документе, которые способны к самоотжигу с образованием структуры в виде шпильки, в которых конструкция содержит (a) один или несколько полинуклеотидов, описанных в данном документе; (b) вторую последовательность, кодирующую спейсерный элемент, который образует петлю структуры в виде шпильки; и (c) третью последовательность, содержащую обратно комплементарную последовательность первой последовательности, расположенную в той же ориентации, что и первая последовательность, где вторая последовательность расположена между первой последовательностью и третьей последовательностью, и третья последовательность функционально связана с первой последовательностью и третьей последовательностью.
Раскрытые последовательности можно использовать для конструирования различных полинуклеотидов, которые не образуют структуры в виде шпильки. Например, двухнитевая РНК может быть образована посредством (1) транскрибирования первой цепи ДНК путем функционального связывания с первым промотором и (2) транскрибирования обратно комплементарной последовательности фрагмента ДНК первой цепи путем функционального связывания со вторым промотором. Каждую цепь полинуклеотида можно транскрибировать из одного вектора экспрессии, или из разных векторов экспрессии. Двойную спираль РНК, обладающую активностью в отношении РНК-интерференции, можно ферментативно превратить в малые интерферирующие РНК для модулирования уровней РНК.
Таким образом, различные варианты реализации направлены на векторы экспрессии, содержащие один или несколько полинуклеотидов или конструкций интерферирующей РНК, описанных в данном документе, кодирующих полинуклеотиды интерферирующей РНК, способные к самоотжигу, в которых конструкция содержит (a) один или несколько полинуклеотидов, описанных в данном документе; и (b) вторую последовательность, содержащую комплементарную (например, обратно комплементарную) последовательность первой последовательности, расположенную в той же ориентации, что и первая последовательность.
Предусмотрены различные композиции и способы для модулирования уровней эндогенной экспрессии одного или нескольких полипептидов, описанных в данном документе (или любой их комбинации, описанной в данном документе) путем стимулирования косупрессии экспрессии гена. Явление косупрессии возникает как результат введения множества копий трансгена в растительную клетку-хозяина. Интеграция множества копий трансгена может приводить к модулированию экспрессии трансгена и нацеленного эндогенного гена. Степень косупрессии зависит от степени идентичности последовательности между трансгеном и нацеленным эндогенным геном. Сайленсинг как эндогенного гена, так и трансгена может происходить из-за обширного метилирования подавляемых локусов (то есть, эндогенного промотора и эндогенного представляющего интерес гена), что может нарушать транскрипцию. В качестве альтернативы, в некоторых случаях косупрессия эндогенного гена и трансгена может происходить посредством пост-транскрипционного сайленсинга гена, при котором транскрипты могут быть получены, но повышенные уровни разрушения препятствуют накоплению транскриптов. Механизм косупрессии посредством пост-транскрипционного сайленсинга генов, как полагают, напоминает РНК-интерференцию в том, что РНК, по-видимому, важна как инициатор и мишень в этих процессах, и может быть опосредован, по меньшей мере частично, тем же молекулярным механизмом, возможно, через РНК-направляемое разрушение мРНК.
Косупрессии нуклеиновых кислотможно достичь путем интеграции множества копий нуклеиновой кислоты или ее фрагментов в качестве трансгенов в геном представляющего интерес растения. Растение-хозяин можно трансформировать вектором экспрессии, содержащим промотор, функционально связанный с нуклеиновой кислотой или ее фрагментами. Различные варианты реализации направлены на векторы экспрессии для стимулирования косупрессии эндогенных генов, содержащих промотор, функционально связанный с полинуклеотидом.
Различные варианты реализации направлены на способы модулирования уровня экспрессии одного или нескольких полинуклеотидов, описанных в данном документе (или любой комбинации полинуклеотидов, как описано в данном документе), путем интеграции множества копий полинуклеотида или полинуклеотидов в геном растения, включая: трансформацию клетки растения-хозяина вектором экспрессии, который содержит промотор, функционально связанный с полинуклеотидом.
Предложены различные композиции и способы модулирования уровня экспрессии эндогенного гена путем модулирования трансляции мРНК. Клетку растения-хозяина можно трансформировать вектором экспрессии, содержащим: промотор, функционально связанный с полинуклеотидом, расположенным в антисмысловой ориентации по отношению к промотору, для обеспечения возможности экспрессии полинуклеотидов РНК, содержащих последовательность, комплементарную части мРНК.
Различные векторы экспрессии для модулирования трансляции мРНК могут содержать: промотор, функционально связанный с полинуклеотидом, в котором последовательность расположена в антисмысловой ориентации по отношению к промотору. Длина полинуклеотидов антисмысловой РНК может варьировать и составлять от приблизительно 15-20 нуклеотидов, приблизительно 20-30 нуклеотидов, приблизительно 30-50 нуклеотидов, приблизительно 50-75 нуклеотидов, приблизительно 75-100 нуклеотидов, приблизительно 100-150 нуклеотидов, приблизительно 150-200 нуклеотидов и приблизительно 200-300 нуклеотидов.
Также на гены можно целенаправленно воздействовать для инактивации путем введения транспозонов (например, IS-элементов) в геномы представляющих интерес растений. Данные мобильные генетические элементы можно ввести с помощью полового перекрестного опыления, и мутантов со вставками можно проверить в отношении потери активности белка. Разрушенный ген родительского растения можно ввести в другие растения путем скрещивания родительского растения с растением, не подвергнутым индуцированному транспозоном мутагенезу, например, путем полового перекрестного опыления. Можно использовать любые стандартные методики селекции, известные специалистам в данной области техники. В одном варианте реализации один или несколько генов можно инактивировать путем вставки одного или нескольких транспозонов. Мутации могут привести к гомозиготному разрушению одного или нескольких генов, к гетерозиготному разрушению одного или нескольких генов, или к комбинации гомозиготных и гетерозиготных разрушений, если разрушен более чем один ген. Подходящие мобильные элементы включают в себя ретротранспозоны, ретропозоны и SINE-подобные элементы. Такие способы известны специалистам в данной области техники.
В качестве альтернативы, на гены можно целенаправленно воздействовать для инактивации путем введения рибозимов, полученных из ряда малых кольцевых РНК, которые способны к саморасщеплению и репликации в растениях. Данные РНК могут реплицироваться либо самостоятельно (РНК вироида), либо с участием вируса-помощника (сателлитные РНК). Примеры подходящих РНК включают полученные из вироида солнечной пятнистости авокадо и сателлитные РНК, полученные из вируса кольцевой пятнистости табака, вируса временной полосатости люцерны, вируса бархатной пятнистости табака, вируса пятнистости Solanum nodiflorum и вируса пятнистости клевера подземного. Различные специфичные к целевой РНК рибозимы известны специалистам в данной области техники.
Как обсуждалось в данном документе, экспрессию одного или нескольких полипептидов можно модулировать нетрансгенными средствами, как например, путем создания одной или нескольких мутаций в одном или нескольких генах, как обсуждалось в данном документе. Способы, с помощью которых вводят случайную мутацию в генную последовательность, могут включать химический мутагенез, мутагенез с помощью ЭМС и радиационный мутагенез. Способы, с помощью которых одну или несколько целевых мутаций вводят в клетку, включают без ограничения технологию редактирования генома, в частности, мутагенез, опосредованный нуклеазой с мотивом «цинкового пальца», а также целенаправленные индуцированные локальные повреждения в геноме (TILLING), гомологичную рекомбинацию, олигонуклеотид-направленный мутагенез и мутагенез, опосредованный мегануклеазой. В одном варианте реализации используют TILLING. Он представляет собой технологию мутагенеза, которую можно использовать для создания и/или идентификации полинуклеотидов, кодирующих полипептиды с модифицированной экспрессией и/или активностью. TILLING также обеспечивает возможность отбора растений, несущих такие мутации. В TILLING объединены мутагенез высокой плотности со способами высокопроизводительного скрининга. Способы для проведения TILLING хорошо известны из уровня техники (см. McCallum et al., (2000) Nat Biotechnol 18: 455-457 и Stemple (2004) Nat Rev Genet 5(2): 145-50).
Некоторые неограничивающие примеры мутаций представляют собой делеции, вставки и миссенс-мутации по меньшей мере одного нуклеотида, однонуклеотидные полиморфизмы и простую повторяющуюся последовательность. После внесения мутации можно провести скрининг для идентификации мутаций, которые создают ранние стоп-кодоны или другим образом нефункциональные гены. После внесения мутации можно провести скрининг для идентификации мутаций, которые создают функциональные гены, способные экспрессироваться на повышенных уровнях. Скрининг мутантов можно выполнить путем секвенирования, или с использованием одного или нескольких зондов или праймеров, специфичных для данного гена или белка. Можно также создать в полинуклеотидах конкретные мутации, которые приводят в результате к модулированию экспрессии гена, модулированию стабильности мРНК или модулированию стабильности белка. Такие растения названы в данном документе как «неприродные» или «мутантные» растения. Как правило, мутантные или неприродные растения будут включать в себя по меньшей мере часть чужеродной или синтетической или созданной человеком нуклеиновой кислоты (например, ДНК или РНК), которая не присутствовала в растении до проведения с ним манипуляций. Чужеродной нуклеиновой кислотой может быть один нуклеотид, два или более нуклеотида, два или более смежных нуклеотидов или два или более несмежных нуклеотидов например, по меньшей мере 10, 20, 30, 40, 50,100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400 или 1500 или более смежных или несмежных нуклеотидов.
Мутантные или неприродные растения или клетки растения могут содержать любую комбинацию одной или нескольких мутаций в одном или нескольких генах, что приводит к модулированию уровней белка. Например, мутантные или неприродные растения или клетки растения могут содержать одну мутацию в одном гене; несколько мутаций в одном гене; одну мутацию в двух или более, или трех или более, или четырех или более генах; или несколько мутаций в двух или более, или трех или более, или четырех или более генах. Примеры таких мутаций описаны в данном документе. В качестве дополнительного примера мутантные или неприродные растения или клетки растения могут содержать одну или несколько мутаций в конкретной части гена(ов), например, в участке гена, который кодирует активный сайт белка или его часть. В качестве дополнительного примера мутантные или неприродные растения или клетки растения могут содержать одну или несколько мутаций в участке вне одного или нескольких генов, например, в участке, расположенном выше по цепи или ниже по цепи относительно гена, который он регулирует, при условии, что они модулируют активность или экспрессию гена(ов). Элементы, расположенные выше по цепи, могут включать в себя промоторы, энхансеры или транскрипционные факторы. Некоторые элементы, такие как энхансеры, могут располагаться выше по цепи или ниже по цепи относительно гена, который они регулируют. Элемент(ы) не обязательно расположен рядом с геном, который он регулирует, так как было обнаружено, что некоторые элементы расположены на расстоянии в несколько тысяч пар оснований выше по цепи или ниже по цепи относительно гена, который они регулируют. Мутантные или неприродные растения или клетки растения могут содержать одну или несколько мутаций, расположенных в пределах первых 100 нуклеотидов гена(в), в пределах первых 200 нуклеотидов гена(ов), в пределах первых 300 нуклеотидов гена(ов), в пределах первых 400 нуклеотидов гена(ов), в пределах первых 500 нуклеотидов гена(ов), в пределах первых 600 нуклеотидов гена(ов), в пределах первых 700 нуклеотидов гена(ов), в пределах первых 800 нуклеотидов гена(ов), в пределах первых 900 нуклеотидов гена(ов), в пределах первых 1000 нуклеотидов гена(ов), в пределах первых 1100 нуклеотидов гена(ов), в пределах первых 1200 нуклеотидов гена(ов), в пределах первых 1300 нуклеотидов гена(ов), в первых 1400 нуклеотидов гена(ов) или в пределах первых 1500 нуклеотидов гена(ов). Мутантные или неприродные растения или клетки растения могут содержать одну или несколько мутаций, расположенных в первом, втором, третьем, четвертом, пятом, шестом, седьмом, восьмом, девятом, десятом, одиннадцатом, двенадцатом, тринадцатом, четырнадцатом или пятнадцатом наборе из 100 нуклеотидов гена(ов) или их комбинации. Раскрыты мутантные или неприродные растения или клетки растения (например, мутантные, неприродные или трансгенные растения или клетки растения и т. п., как описано в данном документе), содержащие варианты мутантного полипептида.
В одном варианте реализации семена растений подвергают мутагенезу и затем выращивают из них мутантные растения первого поколения. Затем обеспечивают возможность самоопыления растений первого поколения, и из семян растений первого поколения выращивают растения второго поколения, которые затем подвергают скринингу на наличие мутаций в их локусах. Хотя подвергнутый мутагенезу растительный материал можно подвергать скринингу на наличие мутаций, преимуществом скрининга растений второго поколения является то, что все соматические мутации соответствуют герминативным мутациям. Специалисту в данной области техники будет понятно, что ряд растительных материалов, в том числе, без ограничения, семена, пыльцу, растительную ткань или растительные клетки, можно подвергать мутагенезу для создания мутантных растений. Однако тип растительного материала, подвергаемого мутагенезу, может иметь значение в тех случаях, когда нуклеиновую кислоту растения подвергают скринингу на наличие мутаций. Например, если пыльцу подвергают мутагенезу до опыления не подвергаемого мутагенезу растения, то из семян, полученных в результате такого опыления, выращивают растения первого поколения. Каждая клетка растений первого поколения будет содержать мутации, созданные в пыльце; таким образом, эти растения первого поколения можно затем подергать скринингу на наличие мутаций вместо того, чтобы дожидаться получения второго поколения.
Для создания мутаций можно применять мутагены, которые создают главным образом точечные мутации и короткие делеции, вставки, трансверсии и/или транзиции, включая химические мутагены и облучение. Мутагены включают без ограничения этилметансульфонат, метилметансульфонат, N-этил-N-нитрозомочевину, триэтилмеламин, N-метил-N-нитрозомочевину, прокарбазин, хлорамбуцил, циклофосфамид, диэтилсульфат, акриламид-мономер, мелфалан, азотистый иприт, винкристин, диметилнитрозамин, N-метил-N'-нитронитрозогуанидин, нитрозогуанидин, 2-аминопурин, 7,12-диметилбенз(a)антрацен, этиленоксид, гексаметилфосфорамид, бисульфан, диэпоксиалканы (диэпоксиоктан, диэпоксибутан и т. п.), 2-метокси-6-хлор-9[3-(этил-2-хлорэтил)аминопропиламино]акридина дигидрохлорид и формальдегид.
Также предусматриваются спонтанные мутации в локусе, которые могут не быть непосредственно вызванными мутагеном, при условии, что они приводят к требуемому фенотипу. Подходящие мутагенные средства могут также включать в себя, например, ионизирующее излучение, например, рентгеновское излучение, гамма-излучение, излучение быстрых нейтронов и ультрафиолетовое излучение. Любой способ получения нуклеиновой кислоты растения, известный специалистам в данной области техники, можно использовать для получения нуклеиновой кислоты растения для скрининга в отношении мутаций.
Нуклеиновую кислоту, полученную из отдельных растений, клеток растения или растительного материала, можно необязательно объединить, чтобы ускорить скрининг в отношении мутаций в популяции растений, происходящих из подвергнутых мутагенезу растительных тканей, клеток или материала. Можно подвергать скринингу одно или несколько следующих поколений растений, клеток растения или растительного материала. Размер необязательно объединенной группы зависит от чувствительности используемого способа скрининга.
После необязательного объединения образцов нуклеиновых кислот, их можно подвергнуть методикам полинуклеотид-специфичной амплификации, таким как полимеразная цепная реакция. Любой один или несколько праймеров или зондов, специфичных в отношении гена или последовательностей, непосредственно примыкающих к гену, можно использовать для амплификации последовательностей в пределах необязательно объединенного образца нуклеиновых кислот. Примеры олигонуклеотидных праймеров изложены в SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15 и SEQ ID NO: 16. Предпочтительно, SEQ ID NO: 13 и SEQ ID NO: 14 используют в комбинации для детектирования NtAsn-1. Предпочтительно, SEQ ID NO: 15 и SEQ ID NO: 16 используют в комбинации для детектирования NtAsn-5. Предпочтительно, один или несколько праймеров или зондов конструируют для амплификации участков локуса, в которых с наибольшей вероятностью возникают полезные мутации. Наиболее предпочтительно, праймер конструируют для выявления мутаций в участках полинуклеотида. Кроме того, для праймера(ов) и зонда(ов) желательно избегать известных сайтов полиморфизмов для облегчения скрининга на точечные мутации. Для облегчения выявления продуктов амплификации один или несколько праймеров или зондов можно метить с применением любого общепринятого способа введения метки. Праймер(ы) или зонд(ы) можно сконструировать на основе последовательностей, описанных в данном документе, с помощью способов, которые хорошо известны из уровня техники.
Для облегчения детектирования продуктов амплификации праймер(ы) или зонд(ы) можно пометить с использованием любого общепринятого способа внесения метки. Их можно сконструировать на основе последовательностей, описанных в данном документе, с помощью способов, которые хорошо известны в данной области техники. Полиморфизмы можно идентифицировать с помощью средств, известных в данной области техники, и некоторых из описанных в литературе.
В дополнительном аспекте предложен способ получения мутантного растения. Способ включает обеспечение по меньшей мере одной клетки растения, содержащей ген, кодирующий функциональный полинуклеотид, описанный в данном документе (или любую их комбинацию, описанную в данном документе). Далее, эту по меньшей мере одну клетку растения обрабатывают в условиях, эффективных для модулирования активности полинуклеотида(ов), описанного в данном документе. По меньшей мере одну мутантную клетку растения затем размножают в мутантное растение, где мутантное растение имеет модулированный уровень описанного полипептида(полипептидов) (или любой их комбинации, описанной в данном документе) по сравнению с уровнем у контрольного растения. В одном варианте реализации данного способа получения мутантного растения стадия обработки включает воздействие по меньшей мере на одну клетку химическим мутагенным средством, описываемым выше, и в условиях, эффективных для получения по меньшей мере одной мутантной клетки растения. В другом варианте реализации данного способа стадия обработки включает воздействие по меньшей мере на одну клетку источником ионизирующего излучения в условиях, эффективных для получения по меньшей мере одной мутантной клетки растения. Термин «мутантное растение» включает мутантные растения, в которых генотип модифицирован по сравнению с контрольным растением, предпочтительно с помощью средств, отличных от генной инженерии и генетической модификации.
В некоторых вариантах реализации мутантное растение, мутантная клетка растения или мутантный растительный материал может содержать одну или несколько мутаций, которые встречаются в природе в другом растении, клетке растения или растительном материале и обеспечивают необходимый признак. Данную мутацию можно ввести (например, путем интрогрессии) в другое растение, клетку растения или растительный материал (например, растение, клетку растения или растительный материал с генетическим фоном, отличающимся от такового растения, из которого происходит мутация) для создания мутации, которая не встречается в природе в данном растении, и для обеспечения у указанного растения данного признака. Таким образом, в качестве примера, мутацию, которая встречается в природе в первом растении,можно ввести во второе растение, например, второе растение с другим генетическим фоном относительно первого растения. Специалист в данной области техники, таким образом, может осуществлять поиск и идентифицировать растение, несущее в естественных условиях в своем геноме один или несколько мутантных аллелей генов, описанных в данном документе, которые обеспечивают необходимый признак. Мутантный аллель(и), который встречается в природе, можно перенести во второе растение различными способами, включая селекцию, возвратное скрещивание и интрогрессию, с получением линий, разновидностей или гибридов, которые имеют одну или несколько мутаций в генах, описанных в данном документе. Растения, демонстрирующие необходимый признак, можно отобрать из пула мутантных растений. Предпочтительно отбор осуществляют с использованием данных о нуклеотидных последовательностях, описанных в данном документе. Следовательно, можно осуществлять скрининг в отношении генетического признака по сравнению с контролем. Такой скрининговый подход может включать применение общепринятых методик амплификации и/или гибридизации нуклеиновых кислот, как обсуждается в данном документе. Таким образом, следующий аспект относится к способу идентификации мутантного растения, причем указанный способ включает следующие стадии: (a) обеспечение образца, содержащего нуклеиновую кислоту из растения; и (b) определение нуклеотидной последовательности полинуклеотида, причем отличие в последовательности полинуклеотида по сравнению с полинуклеотидной последовательностью контрольного растения свидетельствует о том, что указанное растение является мутантным растением. В другом аспекте предложен способ идентификации мутантного растения, которое накапливает сниженные уровни аспарагина по сравнению с контрольным растением, причем указанный способ включает следующие стадии: (a) обеспечение образца из растения, подлежащего скринингу; (b) определение того, содержит ли указанный образец одну или несколько мутаций в одном или нескольких полинуклеотидах, описанных в данном документе; и (c) определение содержания по меньшей мере аспарагина в указанном растении.
В другом аспекте предложен способ получения мутантного растения, которое характеризуется сниженными уровнями аспарагина по сравнению с контрольным растением, причем указанный способ включает следующие стадии: (a) обеспечение образца из первого растения; (b) определение того, содержит ли указанный образец одну или несколько мутаций в одном или нескольких полинуклеотидах, описанных в данном документе, которые приводят к снижению уровней аспарагина; и (c) перенос одной или нескольких мутаций во второе растение. Мутацию(и) можно перенести во второе растение с помощью различных способов, которые известны в данной области техники, например, с помощью генной инженерии, манипуляции с генами, интрогрессии, селекции растений, возвратного скрещивания и т. п. В одном варианте реализации первое растение является встречающимся в природе растением. В одном варианте реализации второе растение имеет генетический фон, отличный от такового первого растения.
В другом аспекте предложен способ получения мутантного растения, которое характеризуется сниженными уровнями аспарагина по сравнению с контрольным растением, причем указанный способ включает следующие стадии: (a) обеспечение образца из первого растения; (b) определение того, содержит ли указанный образец одну или несколько мутаций в одном или нескольких полинуклеотидах, описанных в данном документе, которые приводят к снижению уровней аспарагина; и (c) интрогрессию одной или нескольких мутаций из первого растения во второе растение. В одном варианте реализации стадия интрогрессии включает селекцию растений, необязательно включающую возвратное скрещивание и т. п. В одном варианте реализации первое растение является встречающимся в природе растением. В одном варианте реализации второе растение имеет генетический фон, отличный от такового первого растения. В одном варианте реализации первое растение не представляет собой сорт или элитный сорт. В одном варианте реализации второе растение представляет собой сорт или элитный сорт.
Дополнительный аспект относится к мутантному растению (включая мутантное растение сорта или элитного сорта), полученному или получаемому с помощью способов, описанных в данном документе. В некоторых вариантах реализации «мутантное растение» может содержать одну или несколько мутаций, локализованных только в конкретном участке растения, например, в последовательности одного или нескольких полинуклеотидов, описанных в данном документе. Согласно данному варианту реализации остальная часть геномной последовательности мутантного растения будет такой же или практически такой же, как у растения перед осуществлением мутагенеза.
В некоторых вариантах реализации мутантные растения могут содержать одну или несколько мутаций, локализованных в более чем одном участке растения, например, в последовательности одного или нескольких полинуклеотидов, описанных в данном документе, и в одном или нескольких дополнительных участках генома. Согласно данному варианту реализации остальная часть геномной последовательности мутантного растения не будет такой же или не будет практически такой же, как у растения перед осуществлением мутагенеза. В некоторых вариантах реализации мутантные растения могут не содержать одну или несколько мутаций в одном или более, двух или более, трех или более, четырех или более или пяти или более экзонах полинуклеотида(ов), описанного в данном документе; или могут не содержать одну или несколько мутаций в одном или более, двух или более, трех или более, четырех или более или пяти или более интронах полинуклеотида(ов), описанного в данном документе; или могут не содержать одну или несколько мутаций в промоторе полинуклеотида(ов), описанного в данном документе; или могут не содержать одну или несколько мутаций в 3'-нетранслируемом участке полинуклеотида(ов), описанного в данном документе; или могут не содержать одну или несколько мутаций в 5'-нетранслируемом участке полинуклеотида(ов), описанного в данном документе; или могут не содержать одну или несколько мутаций в кодирующем участке полинуклеотида(ов), описанного в данном документе; или могут не содержать одну или несколько мутаций в некодирующем участке полинуклеотида(ов), описанного в данном документе; или любую комбинацию двух или более, трех или более, четырех или более, пяти или более, или шести или более из их частей.
В следующем аспекте предложен способ идентификации растения, клетки растения или растительного материала, содержащих мутацию в гене, кодирующем полинуклеотид, описанный в данном документе, причем указанный способ включает: (a) осуществление мутагенеза растения, клетки растения или растительного материала; (b) получение образца нуклеиновой кислоты из указанного растения, клетки растения или растительного материала или их потомков; и (c) определение нуклеотидной последовательности гена, кодирующего полинуклеотид, описанный в данном документе, или его вариантов или фрагментов, где отличие в указанной последовательности свидетельствует об одной или нескольких мутациях в ней.
Белки «цинковые пальцы» также можно применять для модулирования экспрессии или активности одного или нескольких полинуклеотидов, описанных в данном документе. В различных вариантах реализации последовательность геномной ДНК, содержащую часть или всю кодирующую последовательность полинуклеотида, модифицируют путем мутагенеза, опосредованного нуклеазой с мотивом «цинкового пальца». В последовательности геномной ДНК осуществляют поиск уникального сайта связывания белка «цинковый палец». В качестве альтернативы, в последовательности геномной ДНК осуществляют поиск двух уникальных сайтов для связывания белка «цинковый палец», причем оба сайта находятся на противоположных нитях и недалеко друг от друга, например, на расстоянии 1, 2, 3, 4, 5, 6 или более пар оснований друг от друга. Соответственно, предложены белки «цинковые пальцы», которые связываются с полинуклеотидами.
Белок «цинковый палец» можно сконструировать для распознавания выбранного целевого сайта в гене. Белок «цинковый палец» может содержать любую комбинацию мотивов, полученных из природных ДНК-связывающих доменов «цинковых пальцев» и неприродных ДНК-связывающих доменов «цинковых пальцев», полученных за счет усечения или расширения или способа сайт-направленного мутагенеза в сочетании со способом отбора, таким как без ограничения отбор с помощью фагового дисплея, отбор с помощью бактериальной дигибридной системы или отбор с помощью бактериальной одногибридной системы. Термин «неприродный ДНК-связывающий домен цинковый палец» относится к ДНК-связывающему домену «цинковому пальцу», который связывает последовательность из трех пар оснований в целевой нуклеиновой кислоте и который не происходит из клетки или организма, содержащего нуклеиновую кислоту, которая подлежит модификации. Способы конструирования белка «цинковый палец», который связывает специфические нуклеотидные последовательности, которые являются уникальными для целевого гена, известны из уровня техники.
Нуклеазу с мотивом цинкового пальца можно сконструировать путем создания слияния первого полинуклеотида, кодирующего белок «цинковый палец», который связывается с полинуклеотидом, и второго полинуклеотида, кодирующего неспецифическую эндонуклеазу, такую как без ограничения эндонуклеаза типа IIS. Белок слияния между белком «цинковый палец» и нуклеазой может содержать спейсер, состоящий из двух пар оснований или, в качестве альтернативы, спейсер, состоящий из трех, четырех, пяти, шести, семи или более пар оснований. В различных вариантах реализации нуклеаза цинкового пальца вводит двухцепочечный разрыв в регуляторную область, кодирующую область или некодирующую область последовательности геномной ДНК полинуклеотида и приводит к снижению уровня экспрессии полинуклеотида или снижению активности белка, кодируемого им. Расщепление с помощью нуклеаз с мотивом цинкового пальца часто приводит в результате к делеции ДНК в сайте расщепления с последующей репарацией ДНК путем соединения негомологичных концов.
В других вариантах реализации белок «цинковый палец» можно выбирать для связывания с регуляторной последовательностью полинуклеотида. Более конкретно, регуляторная последовательность может содержать сайт инициации транскрипции, стартовый кодон, участок экзона, границу раздела экзон-интрон, терминатор или стоп-кодон. Соответственно, в настоящем изобретении предложено мутантное, неприродное или трансгенное растение или клетки растения, полученные с помощью мутагенеза, опосредованного нуклеазой с мотивом «цинкового пальца», вблизи от или в пределах одного или нескольких полинуклеотидов, описанных в данном документе, и способы создания такого растения или клетки растения с помощью мутагенеза, опосредованного нуклеазой с мотивом «цинкового пальца». Способы доставки белка «цинковый палец» и нуклеазы с мотивом «цинкового пальца» в растение подобны описанным ниже для доставки мегануклеазы.
В другом аспекте описаны способы получения мутантных, не встречающихся в природе или трансгенных или другим образом генетически модифицированных растений с использованием мегануклеаз, таких как I-CreI. Встречающиеся в природе мегануклеазы, а также рекомбинантные мегануклеазы можно использовать для того, чтобы специфически вызывать двухнитевой разрыв в одном сайте или в относительно небольшом числе сайтов в геномной ДНК растения с разрушением одного или нескольких полинуклеотидов, описанных в данном документе. Мегануклеаза может быть сконструированной мегануклеазой с измененными свойствами распознавания ДНК. Белки мегануклеаз можно доставлять в клетки растения с помощью ряда различных механизмов, известных в данной области техники.
Настоящее изобретение также охватывает применение мегануклеаз для инактивации полинуклеотида или полинуклеотидов, описанных в данном документе (или любой комбинации полинуклеотидов, как описано в данном документе), в клетке растения или в растении. В частности, в настоящем изобретении предложен способ инактивации полинуклеотида в растении с помощью мегануклеазы, причем указанный способ включает: (a) обеспечение клетки растения, содержащей полинуклеотид, описанный в данном документе; (b) введение мегануклеазы или конструкции, кодирующей мегануклеазу, в указанную клетку растения и (c) предоставление возможности мегануклеазе по существу инактивировать полинуклеотид или полинуклеотиды.
Мегануклеазы можно использовать для расщепления сайтов распознавания мегануклеазы в кодирующих участках полинуклеотида. Такое расщепление часто приводит в результате к делеции ДНК в сайте распознавания мегануклеазы с последующей репарацией мутагенной ДНК путем соединения негомологичных концов. Такие мутации в кодирующей последовательности гена являются, как правило, достаточными для инактивации гена. Этот способ модификации клетки растения включает, во-первых, доставку кассеты экспрессии мегануклеазы в клетку растения с помощью подходящего способа трансформации. Для достижения максимальной эффективности желательно связать кассету экспрессии мегануклеазы с селектируемым маркером и отобрать успешно трансформированные клетки в присутствии селективного средства. Этот подход приведет к интеграции кассеты экспрессии мегануклеазы в геном, что, однако, может быть нежелательным, если растение, скорее всего, потребует официального разрешения. В таких случаях кассету экспрессии мегануклеазы (и связанный селектируемый маркерный ген) можно выделить из последующих поколений растения с использованием традиционных методик селекции. В качестве альтернативы, клетки растения можно изначально трансформировать кассетой экспрессии мегануклеазы без селектируемого маркера и можно вырастить на средах без селективного средства. В таких условиях часть обработанных клеток приобретет кассету экспрессии мегануклеазы и будет экспрессировать сконструированную мегануклеазу временно без интеграции кассеты экспрессии мегануклеазы в геном. Поскольку это не обеспечивает эффективность трансформации, то для этой последней процедуры трансформации требуется скрининг большего количества обработанных клеток для получения требуемой модификации генома. Описанный выше подход можно также применить для модификации растительной клетки при использовании белка «цинковый палец» или нуклеазы с мотивом цинкового пальца.
После доставки кассеты экспрессии мегануклеазы клетки растения выращивают, изначально, в условиях, которые являются типичными для конкретной процедуры трансформации, которую использовали. Это может означать, что трансформированные клетки растут на средах при температуре ниже 26 C, зачастую в темноте. Такие стандартные условия можно использовать в течение периода времени, предпочтительно 1-4 дней, чтобы дать растительной клетке восстановиться после процесса трансформации. В любой момент после этого начального периода восстановления температуру роста можно повысить для стимулирования активности сконструированной мегануклеазы в отношении расщепления и мутирования сайта распознавания мегануклеазы.
Для определенных применений может быть желательным точно удалить полинуклеотид из генома растения. Такие применения возможны с использованием пары сконструированных мегануклеаз, каждая из которых расщепляет сайт распознавания мегануклеазы по обе стороны от предполагаемой делеции. Также можно использовать эффекторные нуклеазы TAL (TALEN), которые способны распознавать и связываться с геном и вносить двухнитевой разрыв в геном. Таким образом, в другом аспекте предлагаются способы получения мутантных, не встречающихся в природе или трансгенных или другим образом генетически модифицированных растений, описанных в данном документе, с помощью эффекторных нуклеаз TAL.
Гены, которые были идентифицированы как вовлеченные в синтез аспарагина в ходе обработки в разновидности Берли Nicotinia tabacum, могут являться применимыми для других растений и других разновидностей растений. Вследствие этого настоящее изобретение является, вероятно, воспроизводимым в других растениях и примененимым для селекции с линиями вариантов.
Растения, подходящие для применения в настоящем изобретении, включают, без ограничения, однодольные и двудольные растения и системы клеток растений, в том числе виды из одного из следующих семейств: Acanthaceae, Alliaceae, Alstroemeriaceae, Amaryllidaceae, Apocynaceae, Arecaceae, Asteraceae, Berberidaceae, Bixaceae, Brassicaceae, Bromeliaceae, Cannabaceae, Caryophyllaceae, Cephalotaxaceae, Chenopodiaceae, Colchicaceae, Cucurbitaceae, Dioscoreaceae, Ephedraceae, Erythroxylaceae, Euphorbiaceae, Fabaceae, Lamiaceae, Linaceae, Lycopodiaceae, Malvaceae, Melanthiaceae, Musaceae, Myrtaceae, Nyssaceae, Papaveraceae, Pinaceae, Plantaginaceae, Poaceae, Rosaceae, Rubiaceae, Salicaceae, Sapindaceae, Solanaceae, Taxaceae, Theaceae или Vitaceae.
Подходящие виды могут включать представителей рода Abelmoschus, Abies, Acer, Agrostis, Allium, Alstroemeria, Ananas, Andrographis, Andropogon, Artemisia, Arundo, Atropa, Berberis, Beta, Bixa, Brassica, Calendula, Camellia, Camptotheca, Cannabis, Capsicum, Carthamus, Catharanthus, Cephalotaxus, Chrysanthemum, Cinchona, Citrullus, Coffea, Colchicum, Coleus, Cucumis, Cucurbita, Cynodon, Datura, Dianthus, Digitalis, Dioscorea, Elaeis, Ephedra, Erianthus, Erythroxylum, Eucalyptus, Festuca, Fragaria, Galanthus, Glycine, Gossypium, Helianthus, Hevea, Hordeum, Hyoscyamus, Jatropha, Lactuca, Linum, Lolium, Lupinus, Lycopersicon, Lycopodium, Manihot, Medicago, Mentha, Miscanthus, Musa, Nicotiana, Oryza, Panicum, Papaver, Parthenium, Pennisetum, Petunia, Phalaris, Phleum, Pinus, Poa, Poinsettia, Populus, Rauwolfia, Ricinus, Rosa, Saccharum, Salix, Sanguinaria, Scopolia, Secale, Solanum, Sorghum, Spartina, Spinacea, Tanacetum, Taxus, Theobroma, Triticosecale, Triticum, Uniola, Veratrum, Vinca, Vitis и Zea.
Подходящие виды могут включать Panicum spp., Sorghum spp., Miscanthus spp., Saccharum spp., Erianthus spp., Populus spp., Andropogon gerardii (бородач Жерара), Pennisetum purpureum (слоновая трава), Phalaris arundinacea (двукисточник тростниковидный), Cynodon dactylon (свинорой пальчатый), Festuca arundinacea (овсяница тростниковая), Spartina pectinata (спартина гребешковая), Medicago sativa (люцерна), Arundo donax (арундо тростниковый), Secale cereale (рожь), Salix spp. (ива), Eucalyptus spp. (эвкалипт), Triticosecale (тритикале), бамбук, Helianthus annuus (подсолнечник), Carthamus tinctorius (сафлор красильный), Jatropha curcas (ятрофа), Ricinus communis (клещевина), Elaeis guineensis (масличная пальма), Linum usitatissimum (лен), Brassica juncea, Beta vulgaris (сахарная свекла), Manihot esculenta (маниок), Lycopersicon esculentum (томат), Lactuca sativa (латук), Musyclise alca (банан), Solanum tuberosum (картофель), Brassica oleracea (брокколи, цветная капуста, брюссельская капуста), Camellia sinensis (чай), Fragaria ananassa (земляника), Theobroma cacao (какао), Coffea ycliseca (кофе), Vitis vinifera (виноград), Ananas comosus (ананас), Capsicum annum (острый и сладкий перец), Allium cepa (лук), Cucumis melo (дыня), Cucumis sativus (огурец), Cucurbita maxima (тыква гигантская), Cucurbita moschata (тыква мускатная), Spinacea oleracea (шпинат), Citrullus lanatus (арбуз), Abelmoschus esculentus (бамия), Solanum melongena (баклажан), Rosa spp. (роза), Dianthus caryophyllus (гвоздика), Petunia spp. (петуния), Poinsettia pulcherrima (пуансеттия), Lupinus albus (люпин), Uniola paniculata (овес), полевица (Agrostis spp.), Populus tremuloides (тополь осинообразный), Pinus spp. (сосна), Abies spp. (пихта), Acer spp. (клен), Hordeum vulgare (ячмень), Poa pratensis (мятлик), Lolium spp. (плевел) и Phleum pratense (тимофеевка), Panicum virgatum (просо), SorghuXXXycliseXXXor (сорго, суданская трава), Miscanthus giganteus (мискантус), Saccharum sp. (сахарный тростник), Populus balsamifera (тополь), Zea mays (кукуруза), Glycine max (соя), Brassica napus (канола), Triticum aestivum (пшеница), Gossypium hirsutum (хлопок), Oryza sativa (рис), Helianthus annuus (подсолнечник), Medicago sativa (люцерна), Beta vulgaris (сахарная свекла) или Pennisetum glaucum (просо жемчужное).
Различные варианты реализации направлены на мутантные, неприродные или трансгенные растения или клетки растения, модифицированные для модулирования уровней экспрессии гена, в результате чего получают растение или клетку растения, например, растение табака или клетку растения табака, в которых уровень экспрессии полипептида модулирован в тканях, представляющих интерес, по сравнению с контролем. Раскрытые композиции и способы можно применять для любого вида из рода Nicotiana, в том числе N. rustica и N. tabacum (например, LA B21, LN KY171, TI 1406, Basma, Galpao, Perique, Beinhart 1000-1 и Petico). Другие виды включают N. acaulis, N. acuminata, N. africana, N. alata, N. ameghinoi, N. amplexicaulis, N. arentsii, N. attenuata, N. azambujae, N. benavidesii, N. benthamiana, N. bigelovii, N. bonariensis, N. cavicola, N. clevelandii, N. cordifolia, N. corymbosa, N. debneyi, N. excelsior, N. forgetiana, N. fragrans, N. glauca, N. glutinosa, N. goodspeedii, N. gossei, N. hybrid, N. ingulba, N. kawakamii, N. knightiana, N. langsdorffii, N. linearis, N. longiflora, N. maritima, N. megalosiphon, N. miersii, N. noctiflora, N. nudicaulis, N. obtusifolia, N. occidentalis, N. occidentalis subsp. hesperis, N. otophora, N. paniculata, N. pauciflora, N. petunioides, N. plumbaginifolia, N. quadrivalvis, N. raimondii, N. repanda, N. rosulata, N. rosulata subsp. ingulba, N. rotundifolia, N. setchellii, N. simulans, N. solanifolia, N. spegazzinii, N. stocktonii, N. suaveolens, N. sylvestris, N. thyrsiflora, N. tomentosa, N. tomentosiformis, N. trigonophylla, N. umbratica, N. undulata, N. velutina, N. wigandioides и N. x sanderae.
Применение сортов табака и элитных сортов табака также предусмотрено в данном документе. Трансгенное, неприродное или мутантное растение, следовательно, может являться сортом табака или элитным сортом табака, который содержит один или несколько трансгенов или одну или несколько генетических мутаций или их комбинацию. Генетическая мутация(и) (например, один или несколько полиморфизмов) может представлять собой мутации, которые не существуют в природе в отдельной разновидности табака или сорте табака (например, элитном сорте табака), или может представлять собой генетическую мутацию(и), которая существует в природе при условии, что мутация не существует в природе в отдельной разновидности табака или сорте табака (например, в элитном сорте табака).
Особенно применимые разновидности Nicotiana tabacum включают табак типа Берли, табак темного типа, табак паровой обработки и табак типа Ориентал. Неограничивающими примерами разновидностей или сортов являются: BD 64, CC 101, CC 200, CC 27, CC 301, CC 400, CC 500, CC 600, CC 700, CC 800, CC 900, Coker 176, Coker 319, Coker 371 Gold, Coker 48, CD 263, DF911, DT 538 LC Galpao tobacco, GL 26H, GL 350, GL 600, GL 737, GL 939, GL 973, HB 04P, HB 04P LC, HB3307PLC, гибрид 403LC, гибрид 404LC, гибрид 501 LC, K 149, K 326, K 346, K 358, K394, K 399, K 730, KDH 959, KT 200, KT204LC, KY10, KY14, KY 160, KY 17, KY 171, KY 907, KY907LC, KY14xL8 LC, Little Crittenden, McNair 373, McNair 944, msKY 14xL8, Narrow Leaf Madole, Narrow Leaf Madole LC, NBH 98, N-126, N-777LC, N-7371LC, NC 100, NC 102, NC 2000, NC 291, NC 297, NC 299, NC 3, NC 4, NC 5, NC 6, NC7, NC 606, NC 71, NC 72, NC 810, NC BH 129, NC 2002, Neal Smith Madole, OXFORD 207, PD 7302 LC, PD 7309 LC, PD 7312 LC, табак «Перик», PVH03, PVH09, PVH19, PVH50, PVH51, R 610, R 630, R 7-11, R 7-12, RG 17, RG 81, RG H51, RGH 4, RGH 51, RS 1410, Speight 168, Speight 172, Speight 179, Speight 210, Speight 220, Speight 225, Speight 227, Speight 234, Speight G-28, Speight G-70, Speight H-6, Speight H20, Speight NF3, TI 1406, TI 1269, TN 86, TN86LC, TN 90, TN 97, TN97LC, TN D94, TN D950, TR (Tom Rosson) Madole, VA 309, VA359, AA 37-1, B13P, Xanthi (Mitchell-Mor), Bel-W3, 79-615, Samsun Holmes NN, KTRDC номер 2 гибрид 49, Берли 21, KY8959, KY9, MD 609, PG01, PG04, PO1, PO2, PO3, RG11, RG 8, VA509, AS44, Бенкет A1, Basma Drama B84/31, Basma I Zichna ZP4/B, Basma Xanthi BX 2A, Batek, Besuki Jember, C104, Coker 347, Criollo Misionero, Delcrest, Djebel 81, DVH 405, Galpão Comum, HB04P, Hicks Broadleaf, Kabakulak Elassona, Kutsage E1, LA BU 21, NC 2326, NC 297, PVH 2110, Red Russian, Samsun, Saplak, Simmaba, Talgar 28, Wislica, Yayaldag, Prilep HC-72, Prilep P23, Prilep PB 156/1, Prilep P12-2/1, Yaka JK-48, Yaka JB 125/3, TI-1068, KDH-960, TI-1070, TW136, Basma, TKF 4028, L8, TKF 2002, GR141, Basma xanthi, GR149, GR153, Petit Havana. Также предусмотрены подразновидности вышеуказанного с низким уровнем превращения, даже если они специально не указаны в данном документе.
В одном варианте реализации используют типовой сорт Nicotiana tabacum типа Берли.
Другие растения, подходящие для применения в настоящем изобретении, включают, без ограничения, чай (Camellia sinensis).
Варианты реализации также направлены на композиции и способы получения мутантных растений, не встречающихся в природе растений, гибридных растений и трансгенных растений, которые были модифицированы для модулирования экспрессии или активности полинуклеотида(ов), описанного в данном документе (или любой их комбинации, как описано в данном документе). Преимущественно, полученные мутантные растения, неприродные растения, гибридные растения или трансгенные растения по общему внешнему виду могут быть подобными или практически такими же, как контрольные растения. Различные фенотипические свойства, такие как степень зрелости, количество листьев на растении, высота стебля, угол отхождения листьев, размер листьев (ширина и длина), длина междоузлия и соотношение листовая пластина-главная жилка, можно оценивать путем полевых наблюдений.
Один аспект относится к семени мутантного растения, неприродного растения, гибридного растения или трансгенного растения, описанного в данном документе. Предпочтительно, семя представляет собой семя табака. Следующий аспект относится к пыльце или семяпочке мутантного растения, неприродного растения, гибридного растения или трансгенного растения, описанного в данном документе. Кроме того, предложено мутантное растение, неприродное растение, гибридное растение или трансгенное растение, описанное в данном документе, которое дополнительно содержит нуклеиновую кислоту, придающую мужскую стерильность.
Также предложена тканевая культура регенерируемых клеток мутантного растения, неприродного растения, гибридного растения или трансгенного растения или его части, как описано в данном документе, при этом из культуры регенерируют растения, способные экспрессировать все морфологические и физиологические свойства родителя. Регенерируемые клетки включают без ограничения клетки из листьев, пыльцы, зародышей, семядолей, гипокотилей, корней, кончиков корней, пыльников, цветков и их части, семяпочек, побегов, стеблей, черешков, сердцевины и семенных коробочек или каллюса или протопластов, полученных из них.
Еще один дополнительный аспект относится к обработанному растительному материалу, такому как обработанный лист или обработанный табак, полученному или получаемому из мутантного, неприродного или трансгенного растения или клетки, причем экспрессия одного или нескольких полинуклеотидов, описанных в данном документе, или активность белка, кодируемого указанными полинуклеотидами, снижена, что приводит к снижению уровней аспарагина в указанном растении.
Предпочтительно, внешний вид указанного растения (например, листа) является практически таким же, как у контрольного растения. Предпочтительно, растение представляет собой растение табака.
Варианты реализации также направлены на композиции и способы получения мутантных, не встречающихся в природе или трансгенных растений или клеток растения, которые были модифицированы для модулирования экспрессии или активности одного или нескольких полинуклеотидов или полипептидов, описанных в данном документе, что может приводить к получению растений или компонентов растения (например, листьев, таких как обработанные или высушенные листья), или клеток растений со сниженными уровнями аспарагина.
Мутантные, неприродные или трансгенные растения, полученные в соответствии со способами, описанными в данном документе, по внешнему виду могут быть подобными контрольным растениям или практически такими же. В одном варианте реализации масса листьев у мутантного, неприродного или трансгенного растения является практически такой же, как у контрольного растения. В одном варианте реализации количество листьев у мутантного, неприродного или трансгенного растения является практически таким же, как у контрольного растения. В одном варианте реализации масса листьев и количество листьев у мутантного, неприродного или трансгенного растения являются практически такими же, как у контрольного растения. В одном варианте реализации высота стебля у мутантных, не встречающихся в природе или трансгенных растений является практически такой же, как у контрольных растений через, например, один, два или три или более месяцев после пересадки в поле или через 10, 20, 30 или 36 или более дней после обрезания верхушек. Например, высота стебля у мутантных, не встречающихся в природе или трансгенных растений не меньше, чем высота стебля у контрольных растений. В другом варианте реализации содержание хлорофилла у мутантных, не встречающихся в природе или трансгенных растений является практически таким же, как у контрольных растений. В другом варианте реализации высота стебля у мутантных, не встречающихся в природе или трансгенных растений является практически такой же, как у контрольных растений, и содержание хлорофилла у мутантных, не встречающихся в природе или трансгенных растений является практически таким же, как у контрольных растений. В других вариантах реализации размер или форма или количество или окраска листьев у мутантных, не встречающихся в природе или трансгенных растений является практически такой же, как у контрольных растений. Предпочтительно, растение представляет собой растение табака.
Согласно другому аспекту предложен способ модулирования (например, снижения) количества аспарагина по меньшей мере в части растения (например, в листьях, таких как обработанные или высушенные листья, или в табаке), причем указанный способ включает следующие стадии: (i) модулирование (например, снижение) экспрессии или активности одного или нескольких полипептидов, описанных в данном документе, причем, предпочтительно, полипептид или полипептиды кодируются соответствующей полинуклеотидной последовательностью, описанной в данном документе; (ii) измерение содержания аспарагина по меньшей мере в части (например, в листьях, таких как обработанные листья) мутантного, неприродного или трансгенного растения, полученного на стадии (i); и (iii) идентификацию мутантного, неприродного или трансгенного растения, в котором содержание аспарагина было снижено по сравнению с контрольным растением. Предпочтительно, внешний вид указанного мутантного, неприродного или трансгенного растения является практически таким же, как у контрольного растения. Предпочтительно, растение представляет собой растение табака.
В другом аспекте предложен способ снижения количества аспарагина по меньшей мере в части обработанного или высушенного растительного материала, такого как обработанный или высушенный лист, причем указанный способ включает следующие стадии: (i) снижение экспрессии или активности одного или нескольких полипептидов, описанных в данном документе, причем, предпочтительно, полипептид или полипептиды кодируются соответствующей полинуклеотидной последовательностью, описанной в данном документе; (ii) сбор растительного материала, такого как один или несколько листьев, и обработка или сушка в течение определенного периода времени; (iii) измерение содержания аспарагина по меньшей мере в части обработанного или высушенного растительного материала, полученного на стадии (ii); и (iv) идентификацию обработанного или высушенного растительного материала, в котором содержание аспарагина было снижено по сравнению с контрольным растением.
Снижение уровня экспрессии по сравнению с контролем может составлять от приблизительно 5% до приблизительно 100%, или снижение может составлять по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100%, которое включает снижение транскрипционной активности или уровня экспрессии полинуклеотида или уровня экспрессии полипептида или их комбинации.
Снижение активности по сравнению с контролем может составлять от приблизительно 5% до приблизительно 100%, или снижение может составлять по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% или 100%.
Полинуклеотиды и рекомбинантные конструкции, описанные в данном документе, можно использовать для модулирования экспрессии ферментов, описанных в данном документе, в видах растений, представляющих интерес, предпочтительно, в табаке.
Множество способов на основе полинуклеотидов можно применять для повышения уровня экспрессии генов, например, в растениях и клетках растений. В качестве примера, можно получать конструкцию, вектор или вектор экспрессии, совместимые с подлежащим трансформированию растением, которые содержат представляющий интерес ген вместе с расположенным выше по цепи промотором, способным осуществлять сверхэкспрессию гена в растении и клетке растения. Иллюстративные промоторы описаны в данном документе. После трансформации и при выращивании в подходящих условиях промотор может управлять экспрессией для модулирования (например, снижения) уровней данного фермента в растении или в его конкретной ткани. В одном иллюстративном варианте реализации создают вектор, несущий один или несколько полинуклеотидов, описанных в данном документе (или любую их комбинацию, как описано в данном документе), для сверхэкспрессии гена в растении или клетке растения. Вектор несет подходящий промотор, такой как промотор 35S вируса мозаики цветной капусты CaMV, выше по цепи относительно трансгена, который управляет его конститутивной экспрессией во всех тканях растения. Вектор также несет ген устойчивости к антибиотику для обеспечения возможности отбора трансформированных каллюсов и клеточных линий.
Различные варианты реализации направлены на способы снижения уровня экспрессии одного или нескольких полинуклеотидов, описанных в данном документе, путем интеграции множества копий полинуклеотида в геном растения, причем указанные способы включают: трансформацию клетки растения-хозяина вектором экспрессии, который содержит промотор, функционально связанный с одним или несколькими полинуклеотидами, описанными в данном документе. Полипептид, кодируемый рекомбинантным полинуклеотидом, может быть нативным полипептидом или может быть гетерологичным по отношению к клетке.
Растение, несущее мутантный аллель одного или нескольких полинуклеотидов, описанных в данном документе (или любой комбинации полинуклеотидов, как описано в данном документе), можно применять в программе селекции растений для создания применимых линий, разновидностей и гибридов. В частности, мутантный аллель интрогрессируют в коммерчески важные разновидности, описанные выше. Таким образом, предложены способы селекции растений, которые предусматривают скрещивание мутантного растения, неприродного растения или трансгенного растения, как описано в данном документе, с растением, характеризующимся иными генетическими особенностями. Способ может дополнительно предусматривать скрещивание растения-потомка с другим растением и, необязательно, повторное скрещивание до тех пор, пока не будет получен потомок с необходимыми генетическими признаками или генетическим фоном. Одной целью, для которой служат такие способы селекции, является введение необходимого генетического признака в другие разновидности, селекционные линии, гибриды или сорта, особенно те, которые представляют коммерческий интерес. Другой целью является облегчение пакетирования генетических модификаций различных генов в отдельной разновидности, линиях, гибридах или сортах растений. Предусмотрены внутривидовые, а также межвидовые скрещивания. Растения-потомки, которые возникают в результате таких скрещиваний, также называемые селекционными линиями, являются примерами не встречающихся в природе растений согласно настоящему изобретению.
В одном варианте реализации предложен способ получения неприродного растения, причем указанный способ включает: (a) скрещивание мутантного или трансгенного растения со вторым растением с получением семени-потомка табака; (b) выращивание семени-потомка в условиях роста растений с получением неприродного растения. Способ может дополнительно включать: (c) скрещивание предыдущего поколения неприродного растения с самим собою или с другим растением с получением семени-потомка; (d) выращивание семени-потомка со стадии (c) в условиях роста растений с получением дополнительных не встречающихся в природе растений; и (e) повторение стадий скрещивания и выращивания (c) и (d) много раз с получением следующих поколений не встречающихся в природе растений. Способ может необязательно предусматривать перед стадией (a) стадию обеспечения родительского растения, которое характеризуется отличительными генетическими особенностями и которое не идентично мутантному или трансгенному растению. В некоторых вариантах реализации в зависимости от программы селекции стадии скрещивания и выращивания повторяют от 0 до 2 раз, от 0 до 3 раз, от 0 до 4 раз, 0 до 5 раз, от 0 до 6 раз, от 0 до 7 раз, от 0 до 8 раз, от 0 до 9 раз или от 0 до 10 раз для получения поколений не встречающихся в природе растений. Возвратное скрещивание является примером такого способа, при котором потомка скрещивают с одним из его родителей или с другим растением, генетически подобным его родителю, чтобы получить растение-потомка в следующем поколении, которое характеризуется генетическими особенностями, более близкими к одному из родителей. Методики селекции растений, в частности, селекции растений, хорошо известны и могут быть использованы в способах согласно настоящему изобретению. В настоящем изобретении дополнительно предложены неприродные растения, полученные с помощью данных способов. Определенные варианты реализации исключают стадию селекции растения.
В некоторых вариантах реализации способов, описанных в данном документе, линии, полученные в результате селекции и скрининга вариантных генов, оценивают в поле с использованием стандартных полевых процедур. В эксперимент включают контрольные генотипы, в том числе исходного не подвергнутого мутагенезу родителя, и места посадки растений располагают в поле согласно рандомизированному полноблочному плану или согласно другому соответствующему планированию поля. Для табака используют стандартные агротехнические приемы, например, табак собирают, взвешивают и отбирают образцы для химического и другого общепринятого тестирования до и во время обработки или сушки. Статистические анализы данных выполняют для подтверждения сходства выбранных линий с родительской линией. Цитогенетические анализы выбранных растений необязательно выполняют для подтверждения взаимосвязей набора хромосом и конъюгации хромосом.
ДНК-фингерпринтинг, однонуклеотидный полиморфизм, микросателлитные маркеры или подобные технологии можно использовать в программе селекции с отбором с помощью маркера (marker-assisted selection, MAS) для переноса или разведения мутантных аллелей гена в других растениях, как описано в данном документе. Например, селекционер может создать сегрегирующие популяции за счет гибридизации генотипа, содержащего мутантный аллель, с генотипом, необходимым с точки зрения агрономии. Растения в поколении F2 или поколении возвратного скрещивания можно подвергнуть скринингу с использованием маркера, полученного из геномной последовательности или ее фрагмента, с использованием одной из методик, перечисленных в данном документе. Растения, идентифицированные как обладающие мутантным аллелем, можно подвергнуть возвратному скрещиванию или самоопылению для получения второй популяции, подлежащей скринингу. В зависимости от предполагаемого характера наследования или используемой технологии MAS может быть необходимым самоопыление отобранных растений перед каждым циклом возвратного скрещивания для облегчения детектирования необходимых отдельных растений. Возвратное скрещивание или другую процедуру селекции можно повторять до тех пор, пока не восстановится необходимый фенотип рекуррентного родителя.
В программе селекции успешные скрещивания дают растения F1, которые являются фертильными. Отобранные растения F1 можно скрещивать с одним из родителей, и растения первого поколения возвратного скрещивания самоопыляют с получением популяции, которую снова подвергают скринингу в отношении экспрессии вариантного гена (например, нулевой версии гена). Процесс возвратного скрещивания, самоопыления и скрининга повторяют, например, по меньшей мере 4 раза до тех пор, пока при последнем скрининге не получат растение, которое является фертильным и в достаточной степени схожим с рекуррентным родителем. Это растение, при необходимости, самоопыляют и затем потомство снова подвергают скринингу, чтобы подтвердить, что растение демонстрирует экспрессию вариантного гена. В некоторых вариантах реализации популяцию растений в поколении F2 подвергают скринингу в отношении экспрессии вариантного гена, например, растение, которое не экспрессирует полипептид из-за отсутствия гена, идентифицируют согласно стандартным способам, например, с помощью методики ПЦР с использованием праймеров, созданных на основе информации о нуклеотидной последовательности полинуклеотида(в), описанного в данном документе (или любой их комбинации, как описано в данном документе).
Гибридные разновидности можно получить путем предотвращения самоопыления женских родительских растений (то есть, родительских семян) первой разновидности, позволяя пыльце с мужских родительских растений второй разновидности оплодотворить женские родительские растения и обеспечивая образование гибридных семян F1 на женских растениях. Самоопыление женских растений можно предотвратить путем кастрации цветков на ранней стадии развития цветка. В качестве альтернативы, образование пыльцы на женских родительских растениях можно предотвратить с применением какой-либо формы мужской стерильности. Например, мужскую стерильность можно получить с помощью цитоплазматической мужской стерильности (CMS) или трансгенной мужской стерильности, где трансген подавляет микроспорогенез и/или образование пыльцы, или вызывает самонесовместимость. Женские родительские растения, характеризующиеся CMS, являются особенно применимы. В вариантах реализации, в которых женские родительские растения характеризуются CMS, пыльцу собирают с мужских фертильных растений и наносят вручную на рыльца женских родительских растений с CMS, и собирают полученные семена F1.
Разновидности и линии, описанные в данном документе, можно использовать для образования простых гибридов F1. В таких вариантах реализации растения родительских разновидностей можно выращивать в виде практически однородно соединенных популяций для облегчения естественного перекрестного опыления мужскими родительскими растениями женских родительских растений. Семя F1, образованное на женских родительских растениях, селективно собирают с помощью обычных средств. Можно также выращивать две разновидности родительского растения в общей массе и собирать смесь гибридных F1 семян, образовавшихся на женском родителе, и семян, образовавшихся на мужском родителе в результате самоопыления. В качестве альтернативы, можно осуществить трехлинейное скрещивание, где простой гибрид F1 используют в качестве женского родителя и скрещивают с другим мужским родителем. В качестве другой альтернативы, можно создать гибриды двойного скрещивания, где потомство F1 двух разных простых гибридов скрещивают само с собой.
Популяцию мутантных, не встречающихся в природе или трансгенных растений можно подвергнуть скринингу или отбору в отношении тех представителей популяции, которые имеют необходимый признак или фенотип. Например, популяцию из потомков линии с одной трансформацией можно подвергнуть скринингу в отношении тех растений, которые имеют необходимый уровень экспрессии или активности полипептида(ов), кодируемого с ее помощью. Для выявления уровней экспрессии или активности можно применять физические и биохимические способы. Они включают анализ по Саузерну или ПЦР-амплификацию для детектирования полинуклеотида; нозерн-блоттинг, анализ с защитой от РНКазы S1, анализ методом удлинения праймера или ПЦР-амплификацию в режиме реального времени для детектирования РНК-транскриптов; ферментные анализы для детектирования ферментативной или рибозимной активности полипептидов и полинуклеотидов; и гель-электрофорез белков, вестерн-блоттинг, иммунопреципитацию и иммуноферментные анализы для детектирования полипептидов. Другие методики, такие как гибридизация in situ, ферментативное окрашивание и иммуноокрашивание, и ферментативный анализ также можно использовать для детектирования присутствия, или экспрессии, или активности полипептидов или полинуклеотидов.
В данном документе описаны мутантные, неприродные или трансгенные клетки растения и растения, содержащие один или несколько рекомбинантных полинуклеотидов, одну или несколько полинуклеотидных конструкций, одну или несколько двухнитевых РНК, один или несколько конъюгатов или один или несколько векторов/векторов экспрессии.
Без ограничения, растения, описанные в данном документе, можно модифицировать для других целей либо до того, либо после того, как экспрессия или активность была модулирована согласно настоящему изобретению. Одна или несколько следующих генетических модификаций могут присутствовать в мутантных, не встречающихся в природе или трансгенных растениях. В одном варианте реализации один или несколько генов, которые вовлечены в превращение промежуточных продуктов азотного обмена, модифицируют с получением растений (например, листьев), у которых в обработанном состоянии присутствуют более низкие уровни по меньшей мере одного специфичного для табака нитрозамина, чем у контрольных растений. Неограничивающие примеры генов, которые можно модифицировать, включают, как описано в данном документе, гены, кодирующие аспарагинсинтетазу, такие как CYP82E4, CYP82E5 и CYP82E10, которые принимают участие в превращении никотина в норникотин и описаны в публикациях WO2006091194, WO2008070274, WO2009064771 и PCT/US2011/021088, и как подробно описано в данном документе. В другом варианте реализации один или несколько генов, которые вовлечены в поглощение тяжелых металлов или транспорт тяжелых металлов, модифицируют с получением растений или частей растений (таких как листья), характеризующихся более низким содержанием тяжелых металлов, чем у контрольных растений или их частей без модификации(й). Неограничивающие примеры включают гены семейства белков, ассоциированных с множественной лекарственной устойчивостью, семейства посредников диффузии катионов (CDF), семейства Zrt-, Irt-подобных белков (ZIP), семейства катионообменников (CAX), семейства транспортеров меди (COPT), семейства АТФаз тяжелых металлов Р-типа (например, HMA, как описано в WO2009074325), семейства гомологов белков макрофагов, ассоциированных с естественной устойчивостью (NRAMP), и семейства транспортеров с АТФ-связывающей кассетой (ABC) (например, MRP, описанных в WO2012/028309, которые участвуют в транспорте тяжелых металлов, таких как кадмий). Термин тяжелый металл в этом документе включает переходные металлы. Примеры других модификаций включают выносливость в отношении гербицидов, например, глифосат является действующим веществом многих гербицидов широкого спектра действия. Устойчивые к глифосату трансгенные растения были разработаны путем переноса гена aroA (глифосат EPSP-синтетаза из Salmonella typhimurium и E.coli). Устойчивые к сульфонилмочевине растения были получены путем трансформации мутантного гена ALS (ацетолактатсинтетаза) из Arabidopsis. Белок OB фотосистемы II из мутантного Amaranthus hybridus был перенесен в растения с получением атразин-устойчивых трансгенных растений; а бромоксинил-устойчивые трансгенные растения были получены путем встраивания гена bxn от бактерии Klebsiella pneumoniae. Другие примеры модификаций приводят к получению растений, которые устойчивы к насекомым. Токсин Bacillus thuringiensis (Bt) может обеспечить эффективный путьзадержки появления Bt-устойчивых вредителей, как недавно показанона брокколи, где пирамидальные cry1Ac и cry1C гены Btконтролировали капустную моль, устойчивую к любому одному белку,и существенно задерживалиразвитие устойчивых насекомых. Другая иллюстративная модификация приводит к получению растений, которые устойчивы к заболеваниям, вызываемым патогенами (например, вирусами, бактериями, грибами). Были разработаны растения, экспрессирующие ген Xa21 (устойчивость к бактериальному некрозу), с растениями, экспрессирующими одновременно ген слияния Bt и ген хитиназы (устойчивость к желтой огневке-травянке и выносливость в отношении ризоктониоза). Другая иллюстративная модификация приводит к измененной репродуктивной способности, такой как мужская стерильность. Другая иллюстративная модификация приводит к получению растений, которые выносливы в отношении абиотического стресса (например, засухи, изменения температуры, засоленности), и выносливые трансгенные растения были получены путем переноса фермента глицеролфосфат-ацилтрансферазы из Arabidopsis; генов, кодирующих маннитдегидрогеназу и сорбитдегидрогеназу, которые вовлечены в синтез маннита и сорбита, улучшающих устойчивость к засухе. Другие иллюстративные модификации могут приводить в результате к получению растений с улучшенным накоплением белков и масел, растений с повышенной эффективностью фотосинтеза, растений с длительным сроком хранения, растений с повышенным содержанием углеводов и растений, устойчивых к грибам; растений, кодирующих фермент, участвующий в биосинтезе алкалоидов. Также рассматриваются трансгенные растения, в которых экспрессия S-аденозил-L-метионин (SAM) и/или цистатионин-гамма-синтазы (CGS) была модулирована.
Один или несколько таких признаков можно интрогрессировать в мутантные, неприродные или трансгенные растения, принадлежащие к другому сорту, или можно непосредственно трансформировать в него. Интрогрессия признака или признаков в мутантные, неприродные или трансгенные растения может быть достигнута любым способом селекции растений, известным в данной области техники, например, селекцией на основе родословной, возвратным скрещиванием, селекцией двойных гаплоидов и т. п. (см. публикации Wernsman, E. A, and Rufty, R. C. 1987. Chapter Seventeen. Tobacco. Pages 669-698 в: Cultivar Development. Crop Species. W. H. Fehr (ed.), MacMillan Publishing Co, Inc., New York, N.Y 761 pp.). Методики на основе молекулярной биологии, описанные выше, в частности, ПДРФ (полиморфизм длин рестрикционных фрагментов) и микросателлитные маркеры, можно использовать при таких возвратных скрещиваниях для детектирования потомства с самой высокой степенью генетической идентичности с рекуррентным родителем. Данный подход позволяет ускорить получение разновидностей, которые характеризуются генетической идентичностью по меньшей мере 90%, предпочтительно, по меньшей мере 95%, более предпочтительно, по меньшей мере 99% с рекуррентным родителем, еще более предпочтительно, генетически идентичных с рекуррентным родителем и которые дополнительно содержат признак или признаки, интрогрессированные от донорского родителя. Такое определение генетической идентичности может основываться на молекулярных маркерах, известных из уровня техники.
Последнее поколение, полученное путем возвратного скрещивания, можно подвергать самоопылению с получением потомков чистой линии в отношении переносимой нуклеиновой кислоты(кислот). Полученные растения обычно характеризуются по существу всеми морфологическими и физиологическими свойствами мутантных, не встречающихся в природе или трансгенных растений в дополнение к перенесенному признаку или признакам (например, одному или нескольким признакам, определяемым одним геном). Точный протокол возвратного скрещивания будет зависеть от изменяемого признака для определения соответствующего протокола испытаний. Хотя способы возвратного скрещивания упрощаются, если переносимый признак представляет собой доминантный аллель, рецессивный аллель также можно переносить. В данном случае может быть необходимо применять исследование потомства для определения успешности переноса необходимого признака.
В различных вариантах реализации предложены мутантные растения, неприродные растения или трансгенные растения, а также биомасса, в которой уровень экспрессии полинуклеотида (или любой комбинации полинуклеотидов, как описано в данном документе) снижен для снижения содержания аспарагина в указанных растениях.
Части таких растений, в частности растений табака, и более конкретно листовую пластинку и среднюю жилку растений табака, можно включать в различные продукты потребления или использовать в их изготовлении, включая без ограничения материалы, образующие аэрозоль, устройства, образующие аэрозоль, курительные изделия, изделия для курения, бездымные продукты и табачные продукты. Примеры материалов, образующих аэрозоль, включают, без ограничения, табачные композиции, виды табака, табачный экстракт, резаный табак, резаный наполнитель, обработанный или высушенный табак, взорванный табак, гомогенизированный табак, восстановленный табак и трубочный табак. Курительные изделия и изделия для курения представляют собой, как правило, устройства, образующие аэрозоль. Примеры курительных изделий или изделий для курения включают без ограничения сигареты, сигариллы и сигары. Примеры бездымных продуктов включают жевательный табак и нюхательный табак. В определенных устройствах, образующих аэрозоль, вместо сгорания (или сжигания) табачную композицию или другой материал, образующий аэрозоль, нагревают, например, с помощью одного или нескольких электрических нагревательных элементов или источников тепла на основе угля с получением аэрозоля. Как правило, в таких нагреваемых курительных изделиях аэрозоль генерируется в результате передачи тепла от источника тепла на физически отделенный субстрат или материал, образующий аэрозоль, который может быть расположен внутри, вокруг или ниже по потоку относительно источника тепла. Во время курения летучие соединения высвобождаются из субстрата, образующего аэрозоль, путем передачи тепла от источника тепла и захватываются воздухом, втягиваемым через курительное изделие. Когда происходит охлаждение выделенных соединений, они конденсируются с образованием аэрозоля, вдыхаемого пользователем. Такие устройства включают, например, генерирующие аэрозоль устройства с электрическим нагревом, в которых аэрозоль генерируется в результате передачи тепла от генерирующего аэрозоль устройства на образующий аэрозоль субстрат нагретого курительного изделия. Предпочтительно, в течение нагревания субстрата, образующего аэрозоль, сгорания или сжигания табака не происходит. Подходящее изделие, образующее аэрозоль, описано в публикации WO2013/098405 и содержит субстрат, образующий аэрозоль, для генерирования ингалируемого аэрозоля при нагревании внутренним нагревательным элементом устройства, генерирующего аэрозоль. Данное изделие может содержать нагреваемое электрическим способом генерирующее аэрозоль устройство, содержащее внутренний нагревательный элемент. Данное изделие также может содержать в линейном последовательном расположении, субстрат, образующий аэрозоль, опорный элемент, размещенный непосредственно ниже по потоку от субстрата, образующего аэрозоль, элемент, охлаждающий аэрозоль, размещенный ниже по потоку от опорного элемента, и наружную обертку, окружающую субстрат, образующий аэрозоль, опорный элемент и элемент, охлаждающий аэрозоль. Опорный элемент может граничить с субстратом, образующим аэрозоль. В субстрат, образующий аэрозоль, может проникать нагревательный элемент устройства, генерирующего аэрозоль.
В настоящем документе термин «сгорание» означает окислительно-восстановительную химическую реакцию, в которой молекулы, вступающие в реакцию, а именно, топливо и окислитель, перемешиваются и перегруппируются, чтобы стать молекулами продукта с одновременным высвобождением тепла. Сгорание можно положительно определить на основании присутствия соответствующих количеств оксидов азота в газообразных продуктах, которые не образовались в результате разложения нитратов, присутствующих в исходном субстрате, вступающем в реакцию, и однозначного доказательства одновременного общего экзотермического процесса. Изменение соответствующих количеств оксидов азота в газообразных продуктах для сгорания можно определить посредством сравнения общих количеств оксидов азота, образованных в условиях, представляющих интерес (например, в воздухе), и оксидов азота, образованных в тех же условиях, но при отсутствии кислорода (например, в атмосфере чистого азота или гелия).В другом типе нагреваемого устройства, образующего аэрозоль, аэрозоль получают путем передачи тепла от горючего топливного элемента или источника тепла к физически отделенному материалу, образующему аэрозоль, который может быть расположен внутри, вокруг или ниже по потоку относительно источника тепла. Бездымные табачные продукты и различные табаксодержащие материалы, образующие аэрозоль, могут содержать табак в любом виде, в том числе в виде высушенных частиц, обрезков, гранул, порошков или суспензии, нанесенных на другие ингредиенты, смешанных с другими ингредиентами, окруженных другими ингредиентами или иным образом объединенных с другими ингредиентами в любом формате, таком как хлопья, пленки, таблетки, пены или шарики. Используемый в данном документе термин «дым» используют для описания типа аэрозоля, который получен с помощью курительных изделий, таких как горючие сигареты, или при сжигании материала, образующего аэрозоль.
В некоторых вариантах реализации нагревание без сгорания или сжигания растительного материала является предпочтительным.
В одном варианте реализации также предложен обработанный растительный материал из мутантных, трансгенных и не встречающихся в природе растений табака, описанных в данном документе. Способы обработки зеленых табачных листьев известны средним специалистам в данной области техники и включают, без ограничения, воздушную обработку, огневую обработку, паровую обработку и обработку на солнце. Способ обработки зеленых табачных листьев зависит от типа собранного табака. Например, Берли и определенные темные разновидности обычно обрабатывают воздушной обработкой, а трубочный табак, жевательный табак и нюхательный табак обычно обрабатывают огневой обработкой.
В другом варианте реализации также предложен высушенный растительный материал из мутантных, трансгенных и не встречающихся в природе растений, описанных в данном документе. Способы сушки листьев известны средним специалистам в данной области техники и включают, без ограничения, воздушную сушку и сушку на солнце. Точный способ сушки листьев зависит от типа собранного растения. Предпочтительно, растительный материал высушивают после сбора. Таким образом, в данном документе предусмотрено применение высушенного материала и послеуборочного высушенного материала. Процесс сушки может активировать один или несколько генов, связанных со старением. Экспрессию активности генов и белков, описанных в данном документе, можно контролировать в ходе обработки или сушки. Аналогично, уровни аспарагина, никотина, глутамина, аспарагиновой кислоты и глутаминовой кислоты можно контролировать в ходе обработки или сушки. В качестве примера, измерения можно проводить через 0, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 часов обработки или сушки. В качестве примера, измерения можно проводить через 0 дней, 1 день, 2 дня, 3 дня, 4 дня, 5 дней или 10 дней или более.
В другом варианте реализации описаны табачные продукты, в том числе табаксодержащие материалы, образующие аэрозоль, содержащие растительный материал, такой как листья, предпочтительно, обработанные или высушенные листья, из мутантных растений табака, трансгенных растений табака или не встречающихся в природе растений табака, описанных в данном документе. Табачные продукты, описанные в данном документе, могут представлять собой смешанный табачный продукт, который может дополнительно содержать немодифицированный табак.
Количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте, описанном в данном документе, после обработки или сушки (например, 3 дней обработки или сушки) приблизительно на 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% и 100% ниже - например, приблизительно на 200% или 300% ниже, по сравнению с контрольным растением. Предпочтительно, количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте, описанном в данном документе, после обработки или сушки (например, 3 дней обработки или сушки) по меньшей мере приблизительно на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% и 100% ниже - например, приблизительно на 200% или 300% ниже, по сравнению с контролем, более предпочтительно, по меньшей мере приблизительно на 80% ниже или по меньшей мере приблизительно на 90% ниже по сравнению с контролем.
Количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте, описанном в данном документе, после обработки или сушки (например, 3 дней обработки или сушки) составляет приблизительно 40 мг/г сухой массы листьев или менее, более предпочтительно, приблизительно 35 мг/г сухой массы листьев или менее, более предпочтительно, приблизительно 30 мг/г сухой массы листьев или менее, более предпочтительно, приблизительно 25 мг/г сухой массы листьев или менее, более предпочтительно, приблизительно 20 мг/г сухой массы листьев или менее, более предпочтительно, приблизительно 15 мг/г сухой массы листьев или менее, более предпочтительно, приблизительно 12 мг/г сухой массы, более предпочтительно, приблизительно 10 мг/г сухой массы сухой массы листьев или менее, или, более предпочтительно, приблизительно 5 мг/г сухой массы листьев или менее. Предпочтительно, отбирают листья нижней-средней части стебля растения.
Количество акриламида в аэрозоле, полученном из растения, части растения, растительного материала, растительного продукта или табачного продукта после обработки или сушки и полученном посредством нагревания, по меньшей мере приблизительно на 70% ниже по сравнению с контролем. В качестве примера, количество акриламида в аэрозоле, полученном из табачного продукта после обработки или сушки и полученном посредством нагревания, составляет приблизительно 1,5 мкг на сигарету.
Количество акриламида в аэрозоле, полученном из растения, части растения, растительного материала, растительного продукта или табачного продукта после обработки или сушки и полученном посредством сгорания, по меньшей мере приблизительно на 55% ниже по сравнению с контролем. В качестве примера, количество акриламида в аэрозоле, полученном из табачного продукта после обработки или сушки и полученном посредством сгорания, составляет приблизительно 2,5 мкг на сигарету.
Предпочтительно, количество акриламида в аэрозоле, полученном из растения, части растения, растительного материала, растительного продукта или табачного продукта после обработки или сушки и полученном посредством сгорания или нагревания, по меньшей мере приблизительно на от 55% до приблизительно 70% ниже по сравнению с контролем. В качестве примера, количество акриламида в аэрозоле, полученном из табачного продукта после обработки или сушки и полученном посредством сгорания или нагревания, составляет приблизительно от 1,5 мкг на сигарету до 2,5 мкг на сигарету.
Количество никотина в растении, части растения, растительном материале, растительном продукте, табачном продукте или аэрозоле может являться по существу таким же, как количество никотина из растений, частей растений, растительных материалов, изделий для курения, бездымных продуктов и аэрозолей из контрольного растения. Например, количество никотина может находиться в пределах приблизительно 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% или менее от количества никотина, которое присутствует в контрольном растении. Предпочтительно, количество никотина может находиться в пределах приблизительно 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% или менее от количества никотина, которое присутствует в контрольном растении. Количество никотина, как правило, составляет от приблизительно 15 до 20 мг/г сухой массы биомассы листьев.
В одном варианте реализации количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте после обработки (например, обработанный формованный лист) по меньшей мере приблизительно на 20% ниже (например, по меньшей мере приблизительно на 22% ниже), и количество акриламида в аэрозоле, полученном из растения, части растения, растительного материала, растительного продукта или табачного продукта после обработки (например, обработанный формованный лист), по меньшей мере приблизительно на 25% ниже (например, по меньшей мере приблизительно на 24% или ниже), чем в контроле. В другом варианте реализации количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте после обработки (например, обработанный формованный лист) составляет приблизительно 12 мг/г сухой массы или менее, и количество акриламида в аэрозоле, полученном из растения, части растения, растительного материала, растительного продукта или табачного продукта после обработки (например, обработанный формованный лист), составляет по меньшей мере приблизительно 2,5 мкг/сигарету или менее. Предпочтительно, уровень никотина является по существу таким же, как и уровень никотина в контроле.
В другом варианте реализации количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте после обработки (например, обработанный формованный лист) по меньшей мере приблизительно на 70% ниже (например, по меньшей мере приблизительно на 66% ниже), и количество акриламида в аэрозоле, полученном из растения, части растения, растительного материала, растительного продукта или табачного продукта после обработки по меньшей мере приблизительно на 45% ниже (например, по меньшей мере приблизительно на 44% ниже), чем в контроле. В другом варианте реализации количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте после обработки (например, обработанный формованный лист) составляет приблизительно 12 мг/г сухой массы или менее, и количество акриламида в аэрозоле, полученном из растения, части растения, растительного материала, растительного продукта или табачного продукта после обработки, составляет приблизительно 3 мкг/сигарету или менее. Предпочтительно, уровень никотина является по существу таким же, как и уровень никотина в контроле.
В другом варианте реализации количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте после обработки (например, обработанный формованный лист) по меньшей мере приблизительно на 90% ниже (например, по меньшей мере приблизительно на 88% или ниже), и количество акриламида в аэрозоле, полученном из растения, части растения, растительного материала, растительного продукта или табачного продукта после обработки, по меньшей мере приблизительно на 70% ниже, чем в контроле. В другом варианте реализации количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте после обработки (например, обработанный формованный лист) составляет приблизительно 5 мг/г сухой массы или менее, и количество акриламида в аэрозоле, полученном из растения, части растения, растительного материала, растительного продукта или табачного продукта после обработки, составляет приблизительно 1,5 мкг/сигарету или менее. Предпочтительно, уровень никотина является по существу таким же, как и уровень никотина в контроле.
В другом варианте реализации модуляция экспрессии NtASN1 посредством РНКи может: (i) снижать количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте после обработки или сушки (например, обработанный формованный лист) на от 24% до приблизительно 89% (например, от приблизительно 35 мг/г до приблизительно 5 мг/г); (ii) сохранять уровень никотина приблизительно 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% или менее от количества никотина, которое присутствует в контрольном растении (например, на уровне приблизительно 15 и 20 мг/г сухой массы биомассы листьев); и (iii) снижать уровень акриламида в аэрозоле по меньшей мере приблизительно на 70% до 1,5 3 мкг/сигарету или менее. Количество биомассы листа, как правило, составляет приблизительно от 300 до 400 г.
В другом варианте реализации модуляция экспрессии NtASN5 посредством РНКи может: (i) снижать количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте после обработки или сушки (например, обработанный формованный лист) на от приблизительно 35% до приблизительно 66% (например, от приблизительно 31 мг/г до приблизительно 18 мг/г); (ii) сохранять уровень никотина приблизительно 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% или менее от количества никотина, которое присутствует в контрольном растении (например, на уровне приблизительно 15 и 20 мг/г сухой массы биомассы листьев); и (iii) снижать уровень акриламида в аэрозоле по меньшей мере приблизительно на 44% до приблизительно 3 мкг/сигарету или менее. Количество биомассы листа, как правило, составляет приблизительно от 300 до 400 г.
В другом варианте реализации модуляция экспрессии NtASN1 посредством мутагенеза может: (i) снижать количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте после обработки или сушки (например, обработанный формованный лист) на от приблизительно 17% до приблизительно 83% (например, до от приблизительно 19,4 мг/г до приблизительно 4,7 мг/г); и (iii) снижать уровень акриламида в аэрозоле по меньшей мере приблизительно на 24% до приблизительно 2,5 мкг/сигарету или менее.
В другом варианте реализации модуляция экспрессии NtASN5 посредством мутагенеза может снижать количество аспарагина в растении, части растения, растительном материале, растительном продукте или табачном продукте после обработки или сушки (например, обработанный формованный лист) на от приблизительно 44% (например, до приблизительно 14,5 мг/г). Мутантные, неприродные или трансгенные растения могут найти другие варианты применения, например, в сельском хозяйстве. Например, мутантные, неприродные или трансгенные растения, описанные в данном документе, можно использовать для изготовления корма для животных и продуктов питания для человека.
В настоящем изобретении также предложены способы получения семян, причем указанные способы включают культивирование мутантного растения, неприродного растения или трансгенного растения, описанных в данном документе, и сбор семян культивируемых растений. Семена растений, описанных в данном документе, можно кондиционировать и упаковать в упаковочный материал с помощью средств, известных в данной области техники, с получением промышленного изделия. Упаковочный материал, такой как бумага или ткань, хорошо известен в данной области техники. Упаковка с семенами может иметь этикетку, например, бирку или этикетку, прикрепленную к упаковочному материалу, этикетку, напечатанную на упаковке, которая описывает происхождение содержащихся в ней семян.
Композиции, способы и наборы для генотипирования растений для детектирования, отбора или селекции могут включать в себя средства для детектирования присутствия полинуклеотида (или любой их комбинации, как описано в данном документе) в образце полинуклеотида. Соответственно, описана композиция, содержащая один или несколько праймеров для специфической амплификации по меньшей мере части одного или нескольких полинуклеотидов и, необязательно, один или несколько зондов и, необязательно, один или несколько реагентов для проведения амплификации или детектирования.
Соответственно, раскрыты специфичные в отношении гена олигонуклеотидные праймеры или зонды, содержащие приблизительно 10 или более смежных полинуклеотидов, соответствующих полинуклеотиду или полинуклеотидам, описанным в данном документе. Указанные праймеры или зонды могут содержать или состоять из приблизительно 15, 20, 25, 30, 40, 45 или 50 или более смежных полинуклеотидов, которые гибридизируются (например, специфично гибридизируются) с полинуклеотидом или полинуклеотидами, описанными в данном документе. В некоторых вариантах реализации праймеры или зонды могут содержать или состоять из приблизительно 10-50 смежных нуклеотидов, приблизительно 10-40 смежных нуклеотидов, приблизительно 10-30 смежных нуклеотидов или приблизительно 15-30 смежных нуклеотидов, которые можно использовать в зависимых от последовательности способах идентификации гена (например, Саузерн-гибридизация) или выделения (например, гибридизация in situ бактериальных колоний или бляшек бактериофага) или детектирования гена (например, в виде одного или нескольких праймеров амплификации при амплификации и детектировании нуклеиновой кислоты). Один или несколько специфических праймеров или зондов можно разработать и использовать для амплификации и детектирования части или всего полинуклеотида(ов). В качестве конкретного примера, два праймера можно использовать в протоколе полимеразной цепной реакции для амплификации фрагмента нуклеиновой кислоты, кодирующего нуклеиновую кислоту, такую как ДНК или РНК. Полимеразную цепную реакцию можно также проводить с использованием одного праймера, который получен из последовательности нуклеиновой кислоты, и второго праймера, который гибридизируется с последовательностью выше по цепи или ниже по цепи относительно последовательности нуклеиновой кислоты, такой как промоторная последовательность, 3'-конец мРНК-предшественника или последовательность, происходящая из вектора. Примеры термических и изотермических методик, применимых для амплификации полинуклеотидов in vitro, хорошо известны из уровня техники. Образец может представлять собой растение, клетку растения или растительный материал или табачный продукт или его можно получить из них, при этом табачный продукт изготовлен или получен из растения, клетки растения или растительного материала, как описано в данном документе.
В следующем аспекте также предложен способ детектирования полинуклеотида(ов), описанного в данном документе (или любой их комбинации, описанной в данном документе) в образце, предусматривающий стадию: (a) обеспечения образца, содержащего или предположительно содержащего полинуклеотид; (b) приведения указанного образца в контакт с одним или несколькими праймерами или одним или несколькими зондами для специфического выявления по меньшей мере части полинуклеотида(ов) и (c) детектирования присутствия продукта амплификации, где присутствие продукта амплификации свидетельствует о присутствии полинуклеотида(ов) в образце. В следующем аспекте также предложено применение одного или нескольких праймеров или зондов для специфического детектирования по меньшей мере части полинуклеотида(ов). Также предложены наборы для детектирования по меньшей мере части полинуклеотида(ов), содержащие один или несколько праймеров или зондов для специфического детектирования по меньшей мере части полинуклеотида(ов). Набор может содержать реагенты для амплификации полинуклеотида, например для ПЦР, или реагенты для технологии детектирования с использованием гибридизационного зонда, такой как Саузерн-блоттинг, нозерн-блоттинг, гибридизация in situ или микрочип. Набор может содержать реагенты для технологии детектирования связывания с антителами, такой как вестерн-блоттинг, ELISA, SELDI-масс-спектрометрия или тест-полоски. Набор может содержать реагенты для секвенирования ДНК. Набор может содержать реактивы и инструкции по определению по меньшей мере содержания аспарагина и/или содержания акриламида в аэрозоле в растительном материале, обработанном или высушенном растительном материале или обработанных или высушенных листьях.
В некоторых вариантах реализации набор может содержать инструкции для одного или нескольких описанных способов. Описанные наборы могут быть применимы для определения генетической идентичности, филогенетических исследований, генотипирования, гаплотипирования, анализа родословной или селекции растений, в частности, с количественной оценкой кодоминантных признаков.
В настоящем изобретении также предложен способ генотипирования растения, клетки растения или растительного материала, содержащих полинуклеотид, описанный в данном документе. Генотипирование обеспечивает средства различения гомологов пары хромосом и может быть использовано для различения сегрегантов в популяции растений. Способы с использованием молекулярных маркеров можно применять для филогенетических исследований, характеризующих генетические связи между разновидностями сельскохозяйственных культур, выявления гибридов или соматических гибридов, локализации хромосомных сегментов, влияющих на моногенные признаки, клонирования на основе генетических карт и изучения наследования количественных признаков. В определенном способе генотипирования может использоваться любое количество аналитических методик с молекулярным маркером, включая полиморфизмы длины амлифицированного фрагмента (ПДАФ). ПДАФ являются продуктом аллельных различий между фрагментами амплификации, вызванных изменчивостью нуклеотидной последовательности. Таким образом, в настоящем изобретении дополнительно предложены средства для отслеживания сегрегации одного или нескольких генов или нуклеиновых кислот, а также хромосомных последовательностей, генетически связанных с данными генами или нуклеиновыми кислотами, с применением таких методик, как анализ ПДАФ.
В одном варианте реализации также предложен обработанный или высушенный растительный материал из мутантных, трансгенных и не встречающихся в природе растений, описанных в данном документе. Например, способы обработки или сушки табачных листьев известны специалистам в данной области техники и включают, без ограничения, воздушную обработку, огневую обработку, паровую обработку и обработку на солнце. Способ обработки зеленых табачных листьев зависит от типа собранного табака, как описано в данном документе.
В другом варианте реализации описаны табачные продукты, в том числе табачные продукты, содержащие растительный материал, такой как листья, предпочтительно, обработанный растительный материал, такой как обработанные или высушенные листья, из мутантных, трансгенных и не встречающихся в природе растений, описанных в данном документе, или которые получены согласно способам, описанным в данном документе. Табачные продукты, описанные в данном документе, могут дополнительно содержать немодифицированный табак.
В другом варианте реализации описаны табачные продукты, содержащие растительный материал, предпочтительно, листья, такие как обработанные или высушенные листья, из мутантных, трансгенных и не встречающихся в природе растений, описанных в данном документе. Например, растительный материал можно добавить внутрь или снаружи табачного продукта, вследствие чего при сгорании высвобождается необходимый аромат. Табачный продукт в соответствии с этим вариантом реализации может даже представлять собой немодифицированный табак или модифицированный табак. Табачное продукт в соответствии с этим вариантом реализации может быть получен из мутантного, трансгенного или неприродного растения, которое содержит модификации в одном или нескольких генах, отличающихся от генов, раскрытых в данном документе.
Настоящее изобретение дополнительно описано в Примерах ниже, которые представлены для более подробного описания настоящего изобретения. Эти примеры, в которых изложен предпочтительный режим, предполагаемый в настоящее время для реализации настоящего изобретения, предназначены для иллюстрации, а не для ограничения настоящего изобретения.
ПРИМЕРЫ
Пример 1 - Синтез аспарагина в обработанных листьях табака
В ходе обработки листа табака и, в особенности, листа сортов Берли, активно образуется аспарагин, который составляет приблизительно 50% от суммарного количества аминокислот, присутствующих в обработанных листьях. На фигуре1 показано повышение уровня аспарагина и суммарных свободных аминокислот, которое, главным образом, является следствием повышения уровня аспарагина, в обработанных листьях трех сортов табака Берли, параллельно выращенных и высушенных воздушной обработкой на одном и том же поле и в амбаре. Неожиданно было установлено, что аспарагин является единственной аминокислотой, которая накапливается в заметных количествах в ходе обработки. Принимая во внимание тот факт, что аспарагин синтезируется в течение ранней фазы обработки, реакция в таком случае зависит от активностей аспарагинсинтетазы, которая использует в качестве субстрата глутамин и аспартат. Однако на фигуре1 Asp и Gln демонстрируют исключительно незначительное повышение по сравнению с аспарагином; это свидетельствует, что пулы Gln и Asp, вероятно, варьируют в течение длительного процесса воздушной обработки (>2 месяцев). Какие-либо изменения пула глутамата не наблюдались (данные не показаны).
Фигура 2 свидетельствует, что уровень Gln увеличивался в течение первых 4 дней обработки (96 ч), а затем снижался в начале накопления аспарагина. Напротив, пул аспартата остается, главным образом, постоянным, несмотря на то, что могут наблюдаться некоторые флуктуации. Не опираясь на какую-либо конкретную теорию, считают, что наблюдаемый пул глутамина может выступать в качестве субстрата для аспарагинсинтетазы или аспарагинсинтетаз для образования аспарагина в течение первых 4-5 дней обработки (см. фигуру 3).
Как упоминалось выше, накопление аспарагина, наблюдаемое в ходе обработки, может являться следствием специфичной активности индуцированной старением аспарагинсинтетазы. После нагревания или сгорания аспарагин и восстанавливающие сахара обладают потенциалом образовывать в аэрозоле акриламид посредством реакции Майяра. Существует хорошая корреляция между аспарагином, накопленным в обработанном листе табака или смесях табака, и акриламидом, измерянным в аэрозоле (см. фигуру4). Не опираясь на какую-либо конкретную теорию, считают, что эти данные свидетельствуют, что повышение уровня акриламида в аэрозоле строго зависит от присутствия аспарагина. Кроме того, присутствие аспарагина в тканях листа также зависит от использованного способа обработки, как показано на фигуре 5.
Пример 2 - Гены аспарагинсинтетазы в табаке
6 генов полноразмерной аспарагинсинтетазы были идентифицированы в табаке, которые называют NtASN1-S, NtASN1-T, NtASN3-S, NtASN3-T, NtASN5-S и NtASN5-T. Интересно отметить, что Arabidopsis содержит 3 гена, родственных аспарагинсинтетазе: AtASN1, AtASN2 и AtASN3, а томат - только 2 полноразмерных гена (Solyc06g007180 и Solyc04g055200). Филогенетический анализ с применением предполагаемых белковых последовательностей, полученных из геномных последовательностей, выделенных из генома табака (Sierro et al., 2014), свидетельствует, что в процессе эволюции две группы аспарагинсинтетазы разделились (см. фигуру 6). В действительности, одна группа, содержащая Solyc04g055200, AtASN2 и 3, является близкой к NtASN3-S и NtASN3-T. Продукт гена AtASN2, принадлежащего к данной группе, расположен во флоэме и, по-видимому, является существенным для ассимиляции, распределения и ремобилизации азота в зеленых тканях (Gaufichon et al., 2013 Plant Cell Environ. Feb;36(2):328-42.). Данная группа относится к подклассу Двудольных класса II. Вторая группа, содержащая Solyc06g007180, AtASN1, NtASN1-S, NtASN1-T, NtASN5-S и NtASN5-T, относится к подклассу Двудольных класса I (Gaufichon et al., 2010 Ann Bot. 2010 Jun;105(7):1141-57.). Сверхэкспрессия AtASN1 способствует повышению содержания аспарагина в цветах и развитию стручков и повышению содержания белка в семенах, что свидетельствует об усиленном транспорте аспарагина из источника в потребляющие ткани. Это свидетельствует, что группа AtASN1 является более приверженной перемещению источника азота в семена для обеспечения надлежащего прорастания семян и высокой устойчивости проростков к азот-лимитирующему субстрату (Lam et al., 2003 Plant Physiol. 2003 Jun;132(2):926-35).
Процент идентичности между продуктами гена позволил выяснить, что NtASN3-S и NtASN3-T являются идентичными и весьма близки к Solyc04g055200 и AtASN3 (идентичность 85%); это свидетельствует, что данный кластер генов являлся относительно хорошо сохранившимся в течение эволюции, вероятно, для поддержания ассимиляции азота в вегетативных тканях (см. выше). Интересно отметить, что каждая из выведенных копий белка NtASN1-S/NtASN1-T и NtASN5-S/NtASN5-TN. sylvestris и N. tomemtosiformis характеризуется высоким процентом идентичности, близким к 98%. Это свидетельствует, что функция гена копий ASN1 и ASN5 является, вероятно, консервативной среди каждого предка табака и, вероятно, данные гены произошли от одного единого общего предка, как в томате и арабидопсисе. Более того, NtASN5-S обладает 95% идентичностью с Solyc06g007180, что свидетельствует о подобной функции.
С учетом того, что уровень аспарагина быстро увеличивается в течение процесса воздушной обработки табака Берли, образование аспарагина в течение ранней фазы обработки (см. фигуру1 и фигуру2) может не являться прямым следствием протеолитического расщепления белков или полипептидов, но следствием синтеза de novo в ходе обработки. В таком случае и как упоминается выше, синтез аспарагина может возникать исключительно за счет активной аспарагинсинтетазы или аспарагинсинтетаз.
Для определения экспрессии генов аспарагинсинтетазы в ходе обработки анализировали данные, собранные в результате микроматричных анализов с применением генных чипов Tobarray Affymetrix (Martin et al.BMC Genomics 2012, 13:674). РНК листа выделяли в зависимости от времени в течение первых 96 часов обработки. Были идентифицированы специфичные зонды (идентичность 100%), совпадающие с шестью кодирующими последовательностями ASN (NtPMIa1g47375e1_st для ASN1-T; NtPMIa1g31395e1_st для ASN1-S; NtPMIa1g25255e1_st для ASN5-T; NtPMIa1g57337e1_st для ASN5-S; NtPMIa1g152348e1_st для ASN3-T и NtPMIa1g121582e1_st для ASN3-S). Для получения РНК листья отбирали в средней части стебля Стелла (CH-Берли) непосредственно в поле (-4 ч); время 0 являлось моментом, когда листья вывешивали в амбаре для воздушной обработки. ASN3-S и ASN3-T исключительно слабо экспрессировались в листьях Берли в течение воздушной обработки. Напротив, ASN1-S, ASN1-T и ASN5-S экспрессировались на высоких уровнях в начале обработки (фаза старения, желтения), тогда как ASN5-T повышающе регулировался сразу после сбора, а затем постоянно снижался в течение первых 96 ч обработки (фигура 7).
Для проверки какой-либо перекрестной гибридизации, которая может возникнуть в микроматричных анализах, в особенности в случае генов, которые характеризуются высокой гомологией последовательности, анализировали экспрессию ASN1-S/T и ASN5-S/T с применением технологии секвенирования РНК. Данные представлены на фигуре8. В швейцарском Берли «Стелла» (фигура 8A) данные секвенирования РНК подтвердили, что ASN5-T являлся менее индуцированным в ходе обработки, чем три другие копии ASN1/5 (см. фигуру7), причем после двух первых дней воздушной обработки ASN1-T являлся наиболее интенсивно экспрессируемым геном. Такая активность гена ASN не возникает с подобной интенсивностью в швейцарском Брайт Вирджиния «ITB 683», который классическим способом обрабатывают в печи. В таком табаке через 48 ч обработки экспрессия ASN1-S/T и ASN5-S/T оставалась низкой. В обоих видах табака экспрессия ASN3S/T не индуцировалась обработкой (фигура 8C и D); таким образом, это подтверждает данные Affymetrix, представленные на фигуре7.
Для исследования устойчивости индукции гена ASN в течение ранней фазы обработки Берли из четырех различных образцов Берли (BU1-4: Сеплек, Бенкет A1, Стелла и TN90), выращенных одновременно на швейцарском поле (2013), выделяли РНК листа до и после 60 ч обработки в амбаре. Временную точку выделения РНК увеличивали до 60 ч, поскольку в Швейцарии TN90 требуется больше времени для начала желтения. Данные экспрессии (фигура 9A) демонстрируют, что во всех листьях Берли ASN1-T и ASN5-T являлись снова наиболее и наименее экспрессированными генами ASN в течение воздушной обработки (за исключением ASN1-T в BU2), соответственно, таким образом, это подтверждает данные, представленные на фигуре8. Транскрипты ASN1-S и ASN5-S постоянно и надежно экспрессируются в течение ранней фазы обработки в листьях различных образцов Берли. Фигура 9B подтверждает, что две копии ASN3, происходящие из N. tomentosiformis и N. sylvestris, не индуцировались в ходе обработки; это косвенно подтверждает, что данные две копии, которые являются более близкими к AtASN2 (см. фигуру6), вероятно, характеризуются более основной функцией в вегетативных тканях растения для поддержания физиологических пулов аспарагина.
Анализ соответствующих транскриптов в 8 тканях, отобранных от растений TN90 на стадии цветения и выращенных в поле, свидетельствует, что ASN5-T являлся более активным в цветке, чем в листьях; копии РНК ASN5-T в особенности обнаруживались в незрелом цветке, лепестке и чашелистике (см. фигуру10, левый чертеж). Помимо экспрессии в листьях, ASN1-T и ASN1-S также интенсивно экспрессировались в лепестке, возможно, для ассимиляции нитрата, что является функцией, которая уже была описана у арабидопсиса (Lam et al., 2003 Plant Physiol. 2003 Jun;132(2):926-35) для ортологического гена AtASN1 (см. фигуру6). ASN3-S/T характеризовался низкой основной экспрессией во всех из данных 8 тканей (см. фигуру10, правый чертеж).
Пример 3 - линииASN1 и ASN5 RNAi TN90e3
Сайленсинг копий ASN1-S/ASN1-T и ASN5-S/ASN5-T в растениях табака Берли исследовали для определения того, способствует ли данный подход снижению уровня аспарагина в обработанных листьях Берли. Специфичные фрагменты ДНК клонировали между сильным конститутивным промотором MMV (вируса мозаики мирабилис) или промотором E4, специфичным для старения (аспарагинсинтетазы). Фрагмент гена ASN фланкировали между MMV или E4 и 3' nos терминаторной последовательностью гена нопалин-синтазы Agrobacterium tumefaciens (Cheng et al., 2003 In Vitro Cellular & Developmental Biology-Plant, 39 595 604). Диаграмма конструкций, использованных в данном исследовании, представлена на фигуре 22. Линию TN90e4e5e10 (Zyvert) табака Берли трансформировали по отдельности каждой из пяти конструкций с применением стандартных протоколов трансформации, опосредованной Agrobacterium. TN90e4e5e10 (Zyvert) представляет собой селекцию из мутагенизированной посредством этилметансульфоната (ЭМС) популяции Берли, которая содержит нокаутные мутации в CYP82E4, CYP82E5v2 и CYP82E10, главных генах аспарагинсинтетазы табака (Lewis et al., 2010), для предотвращения образования норникотина. Применение такой фоновой линии может позволить избежать потенциальных трудностей при интерпретации будущих данных ТСНА (табако-специфического нитрозамина).
Для обеспечения отбора растений, которые экспрессируют соответствующие трансгены на высоком уровне, по меньшей мере 10 отдельных растений T0 для каждой конструкции анализировали методом кПЦР. Семена собирали от трех линий T0 каждой конструкции, которые демонстрировали наивысшие уровни событий сайленсинга ASN. Для изучения того, осуществляет ли потомство T1, которое унаследовало фрагмент или фрагменты гена РНКи, эффективный сайленсинг гена, выделяли РНК из 4 растений выбранных независимых событий трансформации для получения семян T2, доступных для последующих полевых испытаний (2016). Контрольные линии TN90e4e5e10 (Zyvert) использовали в качестве фона для сравнения эффективности трансформации.
Три независимых события трансформации выбирали в T0 растениях ASN1-RNAi и ASN5-RNAi на основании уровня транскриптов ASN1 и ASN5. Растения ASN1-RNAi получали для сайленсинга копий как ASN1-T, так и ASN1-S под контролем промотора MMV или промотора E4, и растения ASN5-RNAi получали для сайленсинга копий как ASN5-T, так и ASN5-S под контролем промотора MMV или промотора E4. С применением канамицина выбирали пятнадцать растений для каждого события трансформации и выращивали в теплице. После созревания от каждого растения отбирали четыре листа в среднем-нижнем положении и подвергали воздушной обработке в течение 7 недель.
Концентрации аминокислот и никотина определяли методом ВЭЖХ-МС (высокоэффективной жидкостной хроматографии - масс-спектрометрии): Аликвоты (~30 мг) измельченного табака экстрагировали смесью этанол-вода (1:1; 6 мл) в течение 45 минут при температуре 50°C. Отцентрифугированные экстракты разводили в 10 раз. Разделение методом жидкостной хроматографии проводили на амидной колонке (Waters Acquity BEH amide, 2,1×150 мм, 1,7 мкм) при температуре 45°C при элюировании градиентом 2 мМ формиата аммония в воде+0,25% муравьиная кислота (элюент A) и ацетонитрил+0,1% муравьиная кислота (элюент B), применяя градиент элюента (0 мин. -10% A, 0,5 мин. -10% A, 4 мин. -60% A, 4,5 мин. -60% A, 4,6 мин. -10% A, 6,8 мин. -10% A; поток 0,5 мл/мин.). Для масс-спектрометрического детектирования аминокислот использовали инструмент Q Exactive (Thermo Scientific) в режиме электрораспыления положительно заряженных ионов, позволяющем получить полное сканирование масс-спектра. Концентрации никотина в экстрактах рассчитывали на основании площади его пика на кривой фотодиодного детектора в ультрафиолетовой и видимой области спектра при 260 нм.
Аспарагин измеряли в обработанной листовой пластинке. Через 3 дня обработки в течение фазы пожелтения фрагмент пластинки экстрагировали, РНК выделяли из каждого растения и проводили кПЦР. На фигуре11A показано, что фрагмент гена РНКи ASN1 под контролем обоих промоторов MMV и E4 оказывает влияние на уровень аспарагина в образцах обработанного листа, количество аспарагина варьирует в различных событиях трансформации. E284-4 продемонстрировал снижение уровня аспарагина на 24%, тогда как E284-6 продемонстрировал снижение уровня аспарагина на 89% по сравнению с фоновыми растениями TN90e4e5e10 (Zyvert), выращенными параллельно. Содержание аспарагина коррелировало с уровнем РНК ASN1, измерянным через 3 дня обработки, для обоих промоторов MMV или E4 (фигура 11B); это подтверждает, что аспарагин синтезируется в ходе обработки, и что копии ASN1-T и/или ASN1-S вовлечены в образование аспарагина в течение ранней фазы обработки. Как конститутивный промотор, такой как MMV, так и промотор, специфичный для старения, такой как E4, являются активными для сайленсинга ASN1. Эффективность промотора E4 свидетельствует, что гены ASN1 являются, главным образом, активными в течение старения (пожелтения), что способствует образованию аспарагина в обработанном листе, вероятно, для перераспределения азота из источника - листьев - в семена. Через три дня обработки параллельно с анализами транскрипции также измеряли аспарагин, уже показав подобные профили снижения уровня аспарагина для 6 исследуемых линий, несмотря на то, что всего лишь приблизительно одна треть данной аминокислоты была синтезирована после данного периода обработки (16,4 мг/г в контроле TN90e4e5e10 (Zyvert), данные не показаны), данный пик аспарагина является максимальным через 9-10 дней воздушной обработки после сбора листьев (см. фигуру2).
Аспарагинсинтетаза переносит аминогруппу с глутамина (Gln), первой аминокислоты, которая фиксирует аммоний в форме азота, ассимилированного из запасов нитрата в листьях, на аспартат (Asp). Данная активность с применением АТФ приводит к образованию аспарагина (аспарагина) и глутамата (Glu, см. также фигуру3). Следовательно, можно высказать предположение, что сайленсинг ASN1 приведет к повышению уровня Gln и Asp и снижению Glu.
Вследствие этого также анализировали количество Gln, Asp, Glu и никотина в тех же образцах. В образцах, в которых наблюдалось значительное снижение уровня аспарагина, наблюдалось сопутствующее повышение уровня Gln, т.е. E281-6 и E282-1, и наоборот: в образцах, в которых наблюдался низкий уровень аспарагина, наблюдалось меньшее количество Gln, т.е. E282-7 (фигура 12A). Как для Gln, так и для аспарагина баланс аминокислот, который является следствием сайленсинга генов ASN1, является модифицированным обратным способом в обработанных листьях. Подобный эффект, но менее заметный, чем для Gln, также наблюдался для Asp и Glu. Как Asp, так и Glu демонстрировали повышение уровня (в ~1,5-3 раза) во всех трансформированных линиях, демонстрируя подобные Gln профили (за исключением Asp в E282-7). При изучении активности аспарагинсинтетазы (см. фигуру3) повышение уровня Glu не ожидали.
Сайленсинг ASN1 не оказывал влияния на содержание никотина ни в конструкциях MMV, ни в конструкциях E4; это подтверждает, что активность ASN1-S и/или ASN1-T не связана с путем синтеза никотина и ограничена послеуборочным созреванием листа в течение ранней программы старения.
Подобный эксперимент проводили для сайленсинга ASN5-S и ASN5-T с применением трансгенных линий РНКи. Данные демонстрируют, что уровень аспарагина эффективно снижался, в особенности, в двух линиях, когда фрагмент РНКи ASN5 экспрессировался под контролем конститутивного промотора MMV, и не снижался или снижался исключительно незначительно под контролем промотора E4 (фигура 13A). Интересно отметить, что данные лишь частично коррелировали с уровнем мРНК ASN5, который оценивали методом кПЦР через 3 дня обработки (фигура 13B). Во-первых, в событиях трансформации под контролем промотора E4 наблюдались большие количества мРНК ASN5, чем под контролем MMV; это подтверждает, что промотор E4 является менее эффективным, чем промотор MMV, для сайленсинга ASN5; возможно, это свидетельствует, что программа синтеза белка ASN5 возникает быстрее и раньше, чем образование белка ASN1 после сбора листьев (фигура 7). В определенных линиях процент аспарагина не коррелировал с уровнем мРНК, например, сравните E303-14 с E303-16 или E305-5 с E305-15. Данное наблюдение может быть обусловлено тем фактом, что копия ASN5-T вовлечена в активности, не представленные или исключительно незначительно представленные в обработанном листе (фигура 9), но представленные в цветковых тканях (фигура 10). Через три дня обработки также измеряли аспарагин, и были продемонстрированы подобные профили снижения уровня аспарагина для 6 линий (данные не показаны), как и для трансгенных линий ASN1 (см. текст выше).
Количество Gln, Asp, Glu и никотина также измеряли в линиях ASN5-RNAi (фигура 14). В линиях ASN1-RNAi в образцах, в которых наблюдалось значительное снижение уровня аспарагина, наблюдалось сопутствующее повышение уровня Gln, т.е. E303-1 и E303-16, и напротив: в образцах, в которых наблюдалось высокое содержание аспарагина, наблюдалось более низкое количество Gln, т.е. E303-14 и E-305-16. E305-5 демонстрировал снижение уровня аспарагина менее чем на 10% и повышение уровня Gln, Asp и Glu по сравнению с контролем и сестринскими линиями E-305-15 и E305-16; это свидетельствует, что в определенном случае промотор E4 может частично сайленсировать копии ASN5. Линия E303-1 демонстрировала максимальное снижение уровня мРНК (фигура 13B), значительное снижение уровня аспарагина (66%, фигура 13A) и соответствующее повышение уровня Gln (14-кратное), Asp (3-кратное) и Glu (3-кратное) по сравнению с контролем (фигура 14A, B и C). Таким образом, в случае растений ASN1-RNAi сайленсинг ASN5 приводил к заметному снижению уровня аспарагина и повышению уровня Gln, Asp и Glu. Уровень никотина не менялся в растениях ASN5-RNAi (фигура 14D), что наблюдали для линий ASN1-RNAi; это свидетельствует, что реакция отделена от пути никотина.
Листья в средней части стебля взвешивали в каждом событии трансформации и контрольных линиях. Данные не продемонстрировали заметных изменений биомассы листьев, когда растения ASN1- и ASN5-RNAi выращивали в контролируемых условиях, таких как теплица (фигура 15).
В заключении, семейства генов ASN1 и ASN5 сайленсированы в соответствующих линиях MMV RNAi, таким образом, это свидетельствует о том, что копии генов ASN1 и ASN5 вовлечены в синтез аспарагина в ходе обработки Берли.
Проводили анализы методом перекрестной кПЦР для определения того, сайленсированы ли оба семейства транскриптов ASN1 и ASN5 в соответствующих растениях ASN1-RNAi и ASN5-RNAi под контролем промотора MMV. Данные таблицы 2 демонстрируют, что в растениях ASN5-RNAi оба транскрипта ASN1 и ASN5 сайленсированы, но это менее очевидно в растениях ASN1-RNAi, в особенности, для линий ASN1-1 и ASN1-2. Данное наблюдение, вероятно, обусловлено тем фактом, что как гены ASN5, так и ASN1 (S и T) являются весьма подобными и принадлежат к одному кластеру генов (фигура 6). Однако это может также свидетельствовать о том, что для изменения синтеза аспарагина в обработанных листьях достаточно инактивировать только кластер гена ASN1.
Анализ экспрессии CYP82E4 в ходе обработки Берли свидетельствует, что промотор E4 становится полностью активным лишь через 48 ч обработки; таким образом, это свидетельствует, что все трансгенные конструкции или фрагменты генов под контролем промотора E4 экспрессируются позже в ранней фазе обработки (>48 ч). Вследствие этого на основании данных, представленных на фигуре11 и фигуре13, было высказано предположение, что ASN1-T и -S активируются позже в течение первой фазы процесса обработки, поскольку сайленсинг ASN1 может в равной степени запускаться промоторами MMV и E4. Промотор E4 слабо контролирует сайленсинг ASN5, таким образом, это свидетельствует, что гены ASN1 контролируются последовательностью событий, связанной с индуцированной программой старения листьев.
Пример 4 - Линии варианта ASN (мутанта ЭМС)
Были идентифицированы ЭМС-мутанты ASN для двух копий ASN1-S и -T и двух копий ASN5-S и -T. Проводили скрининг специфичных мутаций на геномной ДНК в предсказанных последовательностях экзонов. Три стоп-мутации были идентифицированы в копиях ASN1-S (ASN1-S_W156*), ASN1-T (ASN1-T_W156*) и ASN5-S (ASN5-S_Q66*), но не в копии ASN5-T. Поскольку в ASN5-T не были идентифицированы стоп-мутации, была выбрана мутация, отличная от стоп-мутации, демонстрирующая показатель SIFT, составляющий 0,0004, в качестве потенциального кандидата, который может заменить стоп-кодон (ASN5-T_G59D).
Семена потомков от родителей M2, несущие мутации (варианты) или являющиеся немутированными в выбранных генах (аут-сегреганты ДТ), генотипировали, идентифицировали и высаживали в поле. 10 растений каждого выбранного варианта выращивали сбоку от 10 растений их соответствующих аут-сегрегантов ДТ на отдельных участках, чтобы предотвратить любое влияние вариабельности поля (т.е. негомогенногоудобрения нитратами и различий почвы). Четыре листа из средней-нижней части стебля отбирали в период сбора урожая и подвешивали на веревках в течение 2 месяцев в амбаре для воздушной обработки. Листья разрезали, и пластинку измельчали с получением мелкоизмельченного порошка. В каждом пуле листьев определяли аспарагин (мг/г). Несмотря на то, что в связи с вариабельностью от участка к участку наблюдались некоторые изменения, наблюдалась однозначная тенденция к снижению уровня аспарагина в ASN1-S_W156*, ASN1-T_W156* и ASN5-S_Q66* по сравнению с их соответствующими сегрегантами ДТ, но не для ASN5-T_G59D (см. таблицу 3).
Процент снижения уровня аспарагина показан на фигуре17. Наибольшее снижение наблюдалось для ASN1-T_W156* (83%), затем для ASN5-S_Q66*(44%) и, наконец, для ASN1-S_W156* (17%). В линии ASN5-T_G59D наблюдалось незначительное повышение уровня аспарагина. Содержание аспарагина в вариантах ASN коррелировало с уровнем активации генов в ходе обработки. В действительности, ASN1-T больше экспрессировался в ходе обработки, чем ASN5-S и ASN1-S через 48 ч и 60 ч обработки (см. фигуру8 и фигуру9). Однако данное наблюдение все же необходимо принимать с осторожностью, поскольку данные для ASN1-T_W156* были получены только с одного участка на поле, и как растения ASN1-T_W156*, так и растения-аут-сегреганты ДТ для данной линии демонстрировали значительное снижение биомассы в данном полевом эксперименте (см. фигуру18).
Снижение биомассы в ASN1-T_W156* и его соответствующих аут-сегрегантах ДТ ASN1-T_WT*, вероятно, является следствием другой мутации или мутаций, все еще присутствующих в обоих вариантах линий. В действительности, трансгенные эксперименты с линиями ASN1/5-RNAi, как правило, свидетельствуют о том, что ASN1-T и любые другие копии ASN1 и ASN5 не оказывают влияния на высоту растения и фенотип биомассы (см. фигуру15). Скрещивания с фоновой сортом AA37 и последующее самоопыление растений позволило авторам настоящего изобретения сделать заключение об отсутствии эффекта стоп-мутации ASN1-T в отношении размера растения. Фигура 18 также демонстрирует, что стоп-мутации в ASN1-S и ASN5-S не оказывали влияния на биомассу растений. Это также справедливо для точковой мутации G59D в ASN5-T, которая, вероятно, согласно данным экспрессии (фигура 7 и фигура 8), предпочтительно, вовлечена в других тканях, таких как цветок (см. фигуру10).
Второй полевой тест проводили с подобными стоп-мутантами для ASN1-T (6 участков, включая 10 аут-сегрегантов растений и 10 растений стоп-мутантов), ASN1-S (6 участков, включая 10 аут-сегрегантов растений и 10 растений стоп-мутантов) и ASN5-S (9 участков, включая 10 аут-сегрегантов растений и 10 растений стоп-мутантов). После обработки листьев (6 листьев середней части стебля) растения-стоп-мутанты ASN1-T демонстрировали снижение уровня аспарагина в листовой пластинке на 19% по сравнению с аут-сегрегантами, растения-стоп-мутанты ASN1-S демонстрировали снижение уровня аспарагина в листовой пластинке на 23% по сравнению с аут-сегрегантами и, наконец, растения-стоп-мутанты ASN5-S демонстрировали снижение уровня аспарагина в листовой пластинке приблизительно на 10% по сравнению с аут-сегрегантами. Данные результаты подтверждают предшествующие результаты, представленные в таблице 3, которые свидетельствуют, что выбранные стоп-мутанты являются эффективными для снижения уровня аспарагина в обработанных листьях. По сравнению с данными для аспарагина из линий ASN-RNAi комбинация трех стоп-мутантов ASN1-T_W156*, ASN1-S_W156* и ASN5-S_Q66* должна обеспечить подобное снижение уровня аспарагина в пластинке обработанных листьев.
На основании этих данных авторы настоящего изобретения заключили, что ASN1-S, ASN1-T и ASN5-S вовлечены в образование аспарагина в течение воздушной обработки листьев табака Берли.
Пример 5 - Влияние снижения уровня аспарагина на трансгенных линиях ASN на образование акриламида
Порошок пластинки листа трансгенных линий MMV-ASN1-RNAi (линия E281-6 и ее соответствующий контроль, см. фигуру11) и MMV-ASN5-RNAi (линия E303-1 и ее соответствующий контроль, см. фигуру13) перемешивали с 50% порошком табака Вирджиния с получением суспендированного и формованного материала листа в соответствии со спецификацией производителя. Получали изделия, образующие аэрозоль, и аэрозоль, генерированный посредством нагревания указанных изделий, подвергали анализу (в режиме согласно Министерству здравоохранения Канады) для измерения акриламида. Данные представлены на фигуре19. Наблюдалось снижение уровня аспарагина на 88% в линиях ASN1-RNAi, которое преобразовывалось в снижение уровня акриламида 70% по сравнению с соответствующим контролем табака. Аналогичные данные были получены со вторыми трансгенными линиями, где наблюдалось снижение уровня аспарагина на 66% в листьях линий ASN5-RNAi и снижение уровня акриламида на 40% в аэрозоле по сравнению с контролем табака.
Акриламид представляет собой биологический маркер (Naufal et al., 2011) для курильщиков. Образование акриламида также анализировали в дыме горючих сигарет (фигура 20). Горючие сигареты получали из обработанных листьев средней части стебля растений ASN1-RNAi с применением следующей процедуры. Табачный лист выдерживали при температуре 22°C и при влажности окружающей среды 60%. Резаный наполнитель получали из листовой пластинки после удаления центральной жилки. Сигареты изготавливали вручную, заполняли с помощью электрической инжекторной машины для сигарет «Powermatic 2» с использованием стандартных сигаретных трубок. Масса резаного наполнителя, введенного в сигарету, варьировала (приблизительно 800 мг) для получения сопротивления затяжке, соответствующего от 120 до 140 мм водяного столба. Затем прочность (твердость) каждой сигареты контролировали вручную. После определения параметров для ряда измельченных наполнителей данные параметры поддерживали для серий каждой изготовленной сигаретной палочки (200 на образец).
В контролируемых растениях содержание аспарагина в резаном наполнителе табака (фигура 20) являлось сравнимым с содержанием аспарагина, измерянным в смешанном порошке для формованного листа (см. фигуру19), и являлось приблизительно на 10% ниже по сравнению с количеством, измерянным на фигуре11. Такое различие обусловлено положением листа на стебле во время сбора: после обработки Берли аспарагин присутствует в большем количестве в листьях нижней части стебля, чем в листьях верхней части стебля. Однако снижение уровня аспарагина в ASN-RNAi по сравнению с растениями дикого типа являлось аналогичным данным, представленным на фигуре11 и фигуре19. Интересно отметить, что снижение уровня акриламида являлось менее эффективным в дыме горючих сигарет (57%), чем в изделии, генерирующем аэрозоль (70%); возможно, это свидетельствует, что реакция Майяра, приводящая к образованию аспарагина, является более эффективной в нагретых, чем в горючих или соженных табачных продуктах.
Снижение уровня акриламида также является эффективным в аэрозоле из изделий, образующих аэрозоль, полученных из пластинки стоп-мутанта ASN1-S (фигура 21). Однако в данном случае данное снижение ограничено в связи с ограниченным влиянием единственного ASN1-S, который способствует синтезу аспарагина в обработанном листе, по сравнению с, например, ASN1-T (на основании данных, представленных на фигуре17). Эти данные свидетельствуют, что как в трансгенных линиях ASN-RNAi, так и в мутантных линиях ASN снижение уровня аспарагина в обработанной пластинке сопровождается последовательным снижением уровня акриламида в дыме горючих сигарет и аэрозоле, образованном в результате нагревания. Другими словами, нокаут копий гена ASN1 и ASN5 приводит к снижению уровня аспарагина в матрице табака и снижению образования акриламида в дыме горючих сигарет и аэрозоле, образованном в результате нагревания.
ПЕРЕЧЕНЬ ИСТОЧНИКОВ
Canales J, Rueda-López M, Craven-Bartle B, Avila C, Cánovas FM. (2012). Novel insights into regulation of asparagine synthetase in conifers. Front Plant Sci. 3:100
Lewis RS, Bowen SW, Keogh MR and Dewey RE. (2010) Three nicotine demethylase genes mediate nornicotine biosynthesis in Nicotiana tabacum L.: functional characterization of the CYP82E10 gene. Phytochemistry 71: 17-18.
Moldoveanu SC (2005) Analysis of Protein Amino Acids in Tobacco Using Microwave Digestion of Plant Material. Contributions to Tobacco Research 21: 451-465
Naufal ZS, Marano KM, Kathman SJ, Wilson CL.(2011) Differential exposure biomarker levels among cigarette smokers and smokeless tobacco consumers in the National Health and Nutrition Examination Survey 1999-2008. Biomarkers 16:222-235
Onoa H, Chudab Y, Ohnishi-Kameyamaa M, H. Yadaa, M. Ishizakac, H. Kobayashia & M. Yoshidaa. 2003. Analysis of acrylamide by LC-MS/MS and GC-MS in processed Japanese foods. Food Additives & Contaminants, 20: 215-220.
Papousek et al.,(2014) Determination of Acrylamide and Acrolein in Smoke from Tobacco and E-Cigarettes, Chromatographia 77: 1145-1151
Sierro N, Battey JN, Ouadi S, Bakaher N, Bovet L, Willig A, Goepfert S, Peitsch MC and Ivanov NV. (2014) The tobacco genome sequence and its comparison with those of tomato and potato. Nat Commun. 5:3833.
Следующий аспект относится к мутантному, не встречающемуся в природе или трансгенному растению или части указанного растения, которое характеризуется сниженной экспрессией или активностью аспарагинсинтетазы, причем указанная аспарагинсинтетаза содержит, состоит из или состоит по существу из: (i) полинуклеотидной последовательности, содержащей, состоящей из или состоящей по существу из последовательности, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7; или (ii) полипептида, кодируемого полинуклеотидом, описанным в (i); или (iii) полипептида, который характеризуется идентичностью последовательности по меньшей мере 78% с SEQ ID NO:2, или SEQ ID NO:4, или SEQ ID NO:6, или SEQ ID NO:8; причем экспрессия или активность аспарагинсинтетазы, изложенной в (i), (ii) или (iii), снижена по сравнению с контрольным растением.
Следующий аспект относится к способу получения растительного материала со сниженными уровнями аспарагина и сниженными уровнями акриламида в аэрозоле, полученном из указанного растительного материала, по сравнению с растительным материалом из контрольного растения, причем указанный способ включает следующие стадии: (a) обеспечение растения или части указанного растения, содержащего полинуклеотид, содержащий, состоящий из или состоящий по существу из последовательности, кодирующей аспарагинсинтетазу, которая характеризуется идентичностью последовательности по меньшей мере 72% с SEQ ID NO:1, или SEQ ID NO:3, или SEQ ID NO:5, или SEQ ID NO:7; (b) снижение экспрессии полинуклеотида или активности белка, кодируемого указанным полинуклеотидом, в растении или части указанного растения ; (c) сбор растительного материала из растения или части указанного растения; (d) обработку или сушку растительного материала; (e) необязательно, измерение уровней аспарагина в растении или части указанного растения и/или измерение уровней акриламида в аэрозоле, полученном из растения или части указанного растения; и (f) получение обработанного или высушенного растительного материала со сниженными уровнями аспарагина и сниженными уровнями акриламида в аэрозоле, полученном из указанного растительного материала, причем, предпочтительно, уровень никотина является по существу таким же, как и уровень никотина в контрольном растении; причем, предпочтительно, образование глутамина, аспарагиновой кислоты и глутаминовой кислоты повышено по сравнению с обработанным или высушенным листом контрольного растения.
Любая публикация, цитированная или описанная в данном документе, предоставляет соответствующую информацию, раскрытую до даты подачи настоящей заявки. Заявления, сделанные в данном документе, не должны быть истолкованы как допускающие, что авторы настоящего изобретения не имеют право датировать более ранним числом такие раскрытия. Все публикации, упомянутые в вышеприведенном описании, включены в данный документ с помощью ссылки. Различные модификации и варианты настоящего изобретения будут очевидны специалистам в данной области техники без выхода за рамки объема и сущности настоящего изобретения. Несмотря на то, что настоящее изобретение было описано на основании конкретных предпочтительных вариантов реализации, следует понимать, что заявленное изобретение не должно слишком ограничиваться такими конкретными вариантами реализации. В действительности различные модификации описанных способов реализации изобретения, которые очевидны специалистам в клеточной, молекулярной биологии и биологии растений или в смежных областях, предназначены находиться в пределах объема прилагаемой формулы изобретения.
ТАБЛИЦА 1
Процент идентичных остатков в областях выравнивания без пропусков между продуктами гена аспарагинсинтетазы (ASN) табака, томата и арабидопсиса.
ТАБЛИЦА 2
Анализ методом кПЦР (относительные единицы экспрессии генов) для определения степени сайленсинга транскриптов ASN1 и ASN5 в трансгенных линиях ASN1-RNAi (ASN1-1, ASN1-2 и ASN1-3) и ASN5-RNAi (ASN5-1, ASN5-2 и ASN5-3) с применением специфичных праймеров ASN1p и ASN5p.
ТАБЛИЦА 3
Анализ аспарагина (мг/г) в листьях после воздушной обработки, отобранных в нижней/средней части стебля в четырех вариантах ASN: ASN1-S_W156* и его соответствующие аут-сегреганты ДТ ASN1-S_WT*, ASN1-T_W156* и его соответствующие аут-сегреганты ДТ ASN1-T_WT*, ASN5-S_Q66* и его соответствующие аут-сегреганты ДТ ASN5-S_WT*, ASN5-T_G59D и его соответствующие аут-сегреганты ДТ ASN5-T_WT*. Каждый вариант и аут-сегрегант ДТ высаживали рядом друг с другом на отдельных участках для предотвращения каких-либо нежелательных эффектов поля (т.е. различий нитратов и почвы).
ПОСЛЕДОВАТЕЛЬНОСТИ
SEQ ID NO:1 - NtASN1-S, последовательность геномной ДНК
atgtgcgggatcttggctgttttgggttgttctgatgattctcaggccaaaagggttcgtgttctcgagctctctcgcaggtaaattatatttcccttcttttctgtattgttaatgttattgtgttgcgttattatattgattctttttatgagaaagtagtttcctgaacgtttgcattttagaataaaagaagactaaagcttaagatagattatatatttagatattcttgacgtggacctatatcttagattcacaatataattttctcggaccaactgttttagcatgacttgtaaatgatgttgatatgaaaagctgtgatttaattagtaggagtaattcatttccatgttacaattacttatctttctcctccttgtgttcttgccactgctagaggcagatctataattcctacttagagtattgaacacattgtacttttgaaattgtaagtgtgcagtttaatatttgttaaaattttgaaggtttttttaattacatgtatatcgtgacgttatatagcctctctacccgcataaggtaggggtaatgtctgcgtacaaataggctgatttcactctgtgttcagttgaatcaatatatgcggcatccaccactgtttgcaaatgcatgggaatggaaaagtcatgcctcatcactgtcacacatgatttttatttatttatttattatatatgattctttaaatctattagtgggtactgaatcttgctaactgatcgattgatgaaacaggttgaagcatcgtggaccagattggagtgggctgtatcaacatggggactgttacttggcacatcagcgtctagctattgttgatcctgcttccggtgatcaacctctgtttaacgaagataagacgattgttgttacggtgggtgcttttaatttcatgtttactctcatttcttgctcagtttactgatgaaaacgaaactaatgctgataactatcaaccaatcaaccacatacaagtaggggtcgactatatgattcattatatcctttcctctattcaaatagttgatgattaatttcatgccaaaaatttcaggtaaatggagagatctacaatcacgagcaacttcgtaagcaaatgcctaatcataagttccggactggcagtgactgtgatgtcattgcacacctagtgagtactcatcttccaacttaggaaacatgtatgtatatgtaaaattattccctgattatattatgtacattaatcagtatgaagaacatggagaagattttgtggacatgctggatgggatcttcgcttttgtgttattggatactcgagataacagctttcttgttgctcgtgatgccattggaattacttccctttatattggttggggacttgatggtaagattccatatgattttctcacttagattttgttattgcaaatgaatatgttgaatccttatatgtactaattgttgtaattctgttccttctcagggtctgtatggatatcatctgagcttaagggcttgaatgatgactgcgaacattttgaagttttcccaccagggcacttgtactctagcaagaatggcggctttaggaggtggtacaatcctccttggttctctgaggccattccttccactcgttatgatcccttagttctcaggcgtgcctttgaaaatgtgagttaatattgttgttgttgaaatggatccaacatgttggttttaaggctatatatcattttgcaggctgttatcaaaaggttgatgactgatgtcccctttggtgttctcctctccgggggactcgattcatccttggttgcttcgattactgctcgctacttggctggtacaaaggctgccaagcagtggggagcacagcttcattccttctgtgttggccttgaggtcagataatcttgagacttgtgtgattacaacattaataacgacaacacctcattcacaagctagtaaggtcagctatatgaaatctgataccgatttgcttgctttcaccgaacagggctcaccggatctcaaggctgcaagagaagttgctgactacttgggaaccgttcaccacgagtttcacttcaccgttcaggttttgatggagcaactctattttcacttgatctatcatcctcttcgtttctcctttgtttacctgatggcaaatgacatgtgttgtatttgcaggatggaattgatgcaattgaagatgttatttaccatattgagacatacgatgtaacgacaatcagagcaagcactcctatgttccttatgtcgcgtaagattaagtcactaggagtgaagatggtcatatctggggaaggatctgatgaagtgtttggtggctacttgtactttcacaaggctcccaacaaggaagagttccacaaggaaacatgtcgcaaggtataagaaagtgcttccagttcctagtttaaataggacaaggagttgagaaaatgttgattttttccctatgcagattaaagcgcttcaccaatatgactgcttaagagcaaataagtcaacatctgcatggggtttagaagctagagtccctttcctagataaggagttcatcaatgttgccatgagtattgatccagagtggaagttggtaagtagttgacccccaattgttgatcgaataaacaatcaactgtccctaaaactatgtatttattcatttcagattaaaccagagcaaaggaggattgaaaagtgggctctaaggagggcctttgatgatgaggagcatccttatctcccaaaggtgtactaaatcgccattttcaaactgatttgcaaatgattttgaatgttatattattaatcaaactaacaatttactcattacattccttgcagcacatcctgtataggcaaaaagaacaattcagtgatggcgtgggctatagttggatagatggactcaaagcacatgctgaacaacatgtgcgtttcaatgactaatcaacgactttctactgctcttatttatcttccaatgcttgtttgttttaaagctgaccgaggaaacaaattttttcctttaataggtgaccaataggatgatgtttaatgcttcacatatattccctcataacacacccattacaaaggaagcatactactataggatgattttcgagcgctttttcccacaggtaatttgttcagataatagtgagaatttatgataaatggactaatttacgtttgttgtcgattttgcaaccttccaacagtgtgagcaagctcacataatcggaattttttgttcaaataataatgcattggaaatcataatgctaaattgagatttacttttaaccatgttcctctaccatttttggttctgcagaattcagctgggctaaccgttcctggaggagcaagtgtggcgtgtagcacagctaaagctgtagagtgggatgcttcttggtcaaagaaccttgatccttcaggcagggctgctattggtgtacataactcggcttatgagaatcatgtacctgctatggctaatgggaatttgaccaaaaaaatcattggtcgtgtgccttctatggtagaagttggtgctgctcccgagctcacaataaagagttag
SEQ ID NO:2 - NtASN1-S, предсказанная аминокислотная последовательность
MCGILAVLGCSDDSQAKRVRVLELSRRLKHRGPDWSGLYQHGDCYLAHQRLAIVDPASGDQPLFNEDKTIVVTVNGEIYNHEQLRKQMPNHKFRTGSDCDVIAHLYEEHGEDFVDMLDGIFAFVLLDTRDNSFLVARDAIGITSLYIGWGLDGSVWISSELKGLNDDCEHFEVFPPGHLYSSKNGGFRRWYNPPWFSEAIPSTRYDPLVLRRAFENAVIKRLMTDVPFGVLLSGGLDSSLVASITARYLAGTKAAKQWGAQLHSFCVGLEGSPDLKAAREVADYLGTVHHEFHFTVQDGIDAIEDVIYHIETYDVTTIRASTPMFLMSRKIKSLGVKMVISGEGSDEVFGGYLYFHKAPNKEEFHKETCRKIKALHQYDCLRANKSTSAWGLEARVPFLDKEFINVAMSIDPEWKLIKPEQRRIEKWALRRAFDDEEHPYLPKHILYRQKEQFSDGVGYSWIDGLKAHAEQHVTNRMMFNASHIFPHNTPITKEAYYYRMIFERFFPQNSAGLTVPGGASVACSTAKAVEWDASWSKNLDPSGRAAIGVHNSAYENHVPAMANGNLTKKIIGRVPSMVEVGAAPELTIKS
SEQ ID NO:3 - NtASN1-T, последовательность геномной ДНК
atgtgcgggatcttggctgttttgggttgttctgatgattctcaggccaaaagggttcgtgttctcgagctctctcgcaggtaaattatattcccttcttttctctattgttaatgttattgtgttgcatgcgttattaattctttttatgagaaagtagtttcctgagcgtttggagctgctactgattgcttgcattttagaataaaacttaagatggattatatatttagatattcttgacgtggacctatatcttagattcacaattttctcggaccaactgttttagcatgacttgtatatgatgttgatatgaaaagctgtggtttagtaattcatttccatgttacaattactaatctttctcctatctgtgttcttgccactgctagaggcagatctataattcgtaattagagtattgaacacattgtacttttcaaaattataagtgcgcaatttaatatttgttaaaattttgaaggcttttttaattacatgtatatcgtgtctcgtatagcctctctacccatgtaaggtctgcgtacacattacccgccccatgccctcccgtgaaattttactggggtttgctgttgttgttggtatcgtgtgtcgcatatactttgttctgttgaatcaatatatgcgtcatctgccactgtttgcaaatgcatggaatggaaaagtcatgcctcatcactgtcacacaggatttttatttatttatttatttattatatatgattctttttcttgatgtgaaatctattagtgggtactgaatctggctaactgactaatgaaacaggttgaagcatcgtggaccagattggagtgggctgtatcaacatggggactgttacttggcacatcagcgtctagctattgttgatcctgcttccggtgatcaacctctgtttaacgaagataagacgattgttgttacggtgggtgcttttaatttcatgtttactctcatttcttgctcagtttactgatgaacatgaaactaatgactatcaatcagccaatcaaccacgcttaattccatacaagtaggggtcgactatatgattcattatatcctttcctctattccggttcctttttattcaaatagttgatgattaatttcatgccaaaaatttcaggtaaatggagagatctacaatcacgagcaacttcgcaagcaaatgcctgatcataagttccggactggaagtgactgtgatgtcattgcacacctagtgagtaatctagattactcatcttccaacttaggaaacatgtatgtattcccctgattatttcatcatctttatgtacattaatcagtatgaagaacatggagaagattttgtggacatgctggatgggatcttcgcttttgtgttactggatactcgagataacagctttcttgttgctcgtgatgccattggaattacttccctttatattggttggggacttgatggtaagattccatattattttctcacttagattttgttattgcaaatgaatatgttgaatctttatatgtagtaactgttgtaatttggttccttctcagggtctgtatggatatcatctgagcttaagggcttgaatgatgactgcgaacattttgaagttttcccaccaggacacttgtactctagcaagaatggcggctttaggaggtggtacaatcctctttggttctctgaggctattccttccactccttatgatcccttagttctcaggcgcgcctttgaaaatgtgagttattattgttgttgttgaaatggatccaacatgttggttctaaggctggatatcattgtgcaggctgttatcaaaaggttgatgactgatgtcccttttggtgttctgctctccgggggactcgattcatccttggttgcttcgattactgcccgctacttggctggcacaaaggctgccaagcagtggggagcacagcttcattccttctgtgttggccttgaggtcagataatcttgagatttctgtgattacaacattaataacgaccacacctcattcacaagctagtaaggtcagctatatgaatctgataccgatttgcttgctttcaaacgaacagggatcaccggatctcaaggctgcaagagaagttgctgactacttgggaaccgttcaccacgagtttcacttcaccgttcaggtattgatggagcaactctattttgacttgatctatcatcctctgcgttcccttttgtttacctgatggcaaatgacatgtgtcgtatttgcaggatggaattgatgcaattgaagatgttatttaccatattgagacatacgatgtaacgacaatcagagcaagcactcctatgttccttatgtcgcgtaagattaagtcactaggagtgaagatggttatatctggggaaggctctgatgaagtgtttggtggctacttgtactttcacaaggctcccaacaaggaagagttccacaaggaaacatgtcgcaaggtataaaaaagtgtttccagttccttgtttaaaaccttatttaaataggataagcaacaacaacaacaacaacccagtataatcccactagtggggtctgaggagggtattgtgtatgcagaccttacccctaccctggggtagagaggctgtttcagatagaaactcgactccctccctccaaaaaccttatttaaataggacaaggagttgagaaaatgttgatctttttcctccatctctatgcagattaaagcacttcaccaatatgactgtttaagagcaaataagtcaacatctgcatggggcttagaagctagagtgcctttcctagataaggagttcatcaatgttgccatgagtattgatccagagtggaagttggtaagtagttgaaccccaattgttgatcgaataaacgatcaactgtccctaaaaccttgtattctatccatttcagattaaaccagagcaaaggaggattgagaagtgggctctaaggagggcctttgatgatgaggagcatccctatctcccaaaggtggtactaaatcgcccttttcaaactgatttgcaagtgatttcgaatgttatattgttaaccaaactaacaatttnaaggctctgatgaagtgtttggtggctacttgtactttcacaaggctcccaacaaagaagagttccacaaggaaacatgtcgcaaggtataaaaaagtgtttccagttccttgtttaaaaccttatttaaataggataagcaacaacaacaacaacaacccagtataatcccactagtggggtctgaggagggtattgtgtatgcagaccttacccctaccctggggtagagaggctgtttcagatagaaactcgactccctccctccaaaaaccttatttaaataggacaaggagttgagaaaatgttgatctttttcctccatctctatgcagattaaagcacttcaccaatatgactgtttaagagcaaataagtcaacatctgcatggggcttagaagctagagtgcctttcctagataaggagttcatcaatgttgccatgagtattgatccagagtggaagttggtaagtagttgaaccccaattgttgatcgaataaacgatcaactgtccctaaaaccttgtattctatccatttcagattaaaccagagcaaaggaggattgagaagtgggctctaaggagggcctttgatgatgaggagcatccctatctcccaaaggtggtactaaatcgcccttttcaaactgatttgcaagtgatttcgaatgttatattgttaaccaaactaacaatttactcattacatttcctgcagcacatcctatacaggcagaaagaacaattcagtgatggcgtaggctatagttggatagatggactcaaagcacatgctgaacaacatgtgcgtttcaatgactaatcaacaactttctaatgttcttatttatcttccaatgtttgtttgttttaaagctgacttggaatttttttgtttcctttaataggtgaccaataggatgatgcttaatgcttcacatatattccctcataacacaccgattacaaaggaagcatactattataggatgattttcgagcgctttttcccacaggtaatttgttcatgtattgtgaggattcatgataaattagctaacttatgctggttttcaaccttgcaaccttccgataagtcacattgttggagtttgttgttcaaatgcgagatttacttttaaccatgttcctctggtatttttggttctgcagaattcagctgggctaaccgttcctggaggagcgagtgtggcgtgtagcacagctaaagctgtagagtgggatgcttcttggtcaaagaaccttgatccttcaggaagggctgctattggtgtacataactcagcttatgagaatcatgaacctgctatggctaatgggaatttggccacaaaaatcattggccgtgcgccgtctatggtagaagttggtgctgctcatgagctcacaataaggagttag
SEQ ID NO:4 - NtASN1-T, предсказанная аминокислотная последовательность
MCGILAVLGCSDDSQAKRVRVLELSRRLKHRGPDWSGLYQHGDCYLAHQRLAIVDPASGDQPLFNEDKTIVVTVNGEIYNHEQLRKQMPDHKFRTGSDCDVIAHLYEEHGEDFVDMLDGIFAFVLLDTRDNSFLVARDAIGITSLYIGWGLDGSVWISSELKGLNDDCEHFEVFPPGHLYSSKNGGFRRWYNPLWFSEAIPSTPYDPLVLRRAFENAVIKRLMTDVPFGVLLSGGLDSSLVASITARYLAGTKAAKQWGAQLHSFCVGLEGSPDLKAAREVADYLGTVHHEFHFTVQDGIDAIEDVIYHIETYDVTTIRASTPMFLMSRKIKSLGVKMVISGEGSDEVFGGYLYFHKAPNKEEFHKETCRKIKALHQYDCLRANKSTSAWGLEARVPFLDKEFINVAMSIDPEWKLIKPEQRRIEKWALRRAFDDEEHPYLPKHILYRQKEQFSDGVGYSWIDGLKAHAEQHVTNRMMLNASHIFPHNTPITKEAYYYRMIFERFFPQNSAGLTVPGGASVACSTAKAVEWDASWSKNLDPSGRAAIGVHNSAYENHEPAMANGNLATKIIGRAPSMVEVGAAHELTIRS
SEQ ID NO:5 - NtASN5-S, последовательность геномной ДНК
atgtgtggaatcttggctttgttgggttgttcagatgattctcaggccaaaagggttcgagttcttgagctctctcgcaggcaatttcccctttcttttctcttttattttattactccatcatataattattctactttcaccttttttgttttttaatttcgatttgtttgctttttgctttcttcatttaatgttcttgcattgctctagctcctatacgaataataggagctaaagtaatagagtgaaggattattttggagcaacggtaaagttatttttgtatagttcacaggttggagccgtaaaagcagttactagtgtttgcattagggtaagttgtctatcaggaaagatatttgtagcaattgaccaccctttggcaaaaaattatgcgtaaaatattactacttgttattgattcaaaaaatatgactttgaacatccttgtatagcctagtggcaaagggtgttcaaaatttttgaacacctttattgaattttctggtttcgtcgttgctgtctacaacacactctcgaagtgcggccttccctcaaaccctacgtgaatatggaacgcctagtgtgccgggatgccctttaggctgagtaatagagtactatgttgatatacctaaaatacggaactggcacgagagataaattcagaatttatagcttaattatttaaaatttatcatttgatttatttcaaccagagttcattctagtagagtttatgttgcatatttgtctgatttgcaccatttgttttaatgttcttgcattactctaatttatttatgaataataagctaaactaattgagtttatgttgatgtaactgacctgagttggcagtgatgtttggctacttgaatttaaaacaagattaagacttgtgagttggcagtaattgtttggatacttgaatttagaacaagaataaaacaagattaattaaggatacattattcctaacccggaccctaattctatagattgattattaaatttttcttggaccaaccgtcaccgtgacctgttcatgatggtgatggcaaaagccataatacacttgatttctgttttcaatttttaaggttcttattattatacttattatatactttgaaaattatgggtcaaaattttaatatttgttattccttccgtatgtttgactcaacttggagttcatgaaaaaaaaaaaaaatttttttaaacttgtggtcttaaacaagtcatcgatatatgtgtggttgtgaatcgcctcgttaggataaagtagaaattttaaagttaaactatttcgaaataaaaaagtatgatagtgtttttgggacatactaaaaaggaaataacacataaaatggaacaaagaagtatattttagtgatttttcacatatatatcagtattttgtatcaaaaatgctggttcagttgaactcattgaactgggtgctccatccacctccgagtactgcaacatgtgaccggtggcagatgcaacccttacgtaccagggtcatctgaacctagtataaatatatttgtgaaaaatccctaaaatttcaataattagtagatttaaaaccttaattttaaaactacaatgagttcagttttaaaatctttatatgtcaaacctatcaagattaaatcctggatccaccttttgcatgtgacagtacaatagccttttgtaattgaaacaaatatttgaattttggtaactatttgatcacataggttgaagcatcgtggaccagattggagtgggatatatcaacatggtgatttttacttagcacatcaacgtttagcaattatcgatcctgcttctggtgatcagcctctgtttaatcaagataagacgattgttgttacagtgagtgcctctaatttacctcttttttttttctttttttttttacgatcgcgttgtttagattatttatggatgaaaatttgtatatttgattattgattgtgtgctaaattttgcaggtcaatggagagatttacaatcatgagaaacttcgtaatcttatgcctaatcacaagttcagaactggaagtgattgtgatgttattgcacatcttgtgagttagtttacttttctcattgaccaacttaggccagtgaattatattaggaatttcgggtgtttggatatggtcaaattagcttataagcacttattatcaattttaacatttttatccatacacgtaactgttcattcataaagtcattttagcacttgatgctcatcaacctatgttgttgggactcttcgaaaatgtcagcccgatagcatttcggagagtccgagcaacatagcaatcaactacttttaatcagttaaacgaacatgctcatagtgataacctctggaaaatatgttatagtaaattagaaaaatttatacgatgtctcactatcggtatgtattgatcagtatgaagaatatggagaaaattttgtggacatgttggatggggtgttctcttttgtattgttggatacgcgcgataacagctttcttgctgctcgtgatgcaattggaattactcccctatatattggttggggacttgatggtaagattttctatacaattttccagttagaaattaaaactgaagctattcctatttactattgaagaatcttgatacatgttatttggtatttcgtcgtgttgttatgtctcaggctctgtgtggatttcatctgagctaaaaggattaaatggtgactgtgaacattttgaagttttccctcccggtcacttgtactcgagcaagaatggcgggtttaggagatggtacaatcctcaatggttctctgaggctattccatcaaatccttacgaccccttagttttgagacgtgccttcgaaaatgtgaggcttgttaatgaatctagtgtacaatcttggatgtttttctctcgactttggttttagcactcaaaaagttgtctgtacaaacgatcttggctataaggagtggtggttaattttgcaggctgttatcaaacgattgatgaccgatgtcccctttggtgttctgctctccgggggacttgattcgtctttggttgcttctgtcactgctcgctacttggctggaacaaaagctgctaagcaatggggagcacagcttcattccttctgtgttggtctcgaggtcagactatcaatcctgagcggagacatatttggggcgggttctgtgggttcaactgaacccattgcttttggttcgaaccatatatacaaatccaagaaaatataagatgtgtacatataatatgacactcattctgagcctactgacgctagatactggactcgcctttggtcttgagactttataatagaaaatagatattcaataccaatttctgtttttcttttctgactgacagggctcaccagatctcaaggctgcaagagaagttgctgactatttgggaaccgttcaccacgagttcaccttcacagttcaggtttgtaaatcgtgttgctactccgatttattcatgtgagtctcttctgttttcctcctagaaagcaacaagtgttcatgttttgcaggatggaattgatgctattgaagatgttatttaccatatcgagacatacgatgtaacaacgatcagagcaagcactcctatgttccttatgtcgcgtaagattaaatcactgggagttaagatggtcatatcaggggaaggctcagatgaactgtttggcggctatttgtacttccacaaggctccgaacaaggaagaattccatgtggagacatgtcacaaggtaataaaacacgtcggctcccttggcaagtctgattctccttgtcattgtttgtctcgggttggaataaattgctataattcgagaagaaatgttaatcctgctctttcactcgcatgcagataaaagcgcttcaccaatacgactgtttgagagcaaataaggcaa
catcagcatggggcttagaagctagagtaccatttctggataaagagttcatcaacgttgctatgagtatcgatcctgaatggaagatggtaacaaaactgagctccaattgtcaatgagatgagaagcataactgaatatgcacgttctctaaaacttgtttttcatccgtttcagattaaacacgatcatggtaggatcgagaagtgggttcttaggaaggcttttgatgatgaggagcaaccctatctcccaaaggtccacaaagtcatgtttttcctactgatttgccgattagtatggatcaaaactaacatgttttgttcttttatgtactctgcagcatattctgtaccggcagaaagaacaattcagtgatggcgtaggctatagttggatcgatggactcaaagcacatgctgaacaacatgtgagcttctaaataaatccagaactattacatcgttatcgtccaattttcgtttgttttgctgtcagctgatcttggaaacaattcttttctctcaggtgactgataggatgatgcttaatgctgcacatatcttccctcacaacactccaactacaaaggaagcatactattacaggatgattttcgagaggttcttcccacaggtacttatagcgcgagactagaatcatatatctcttaaatggaaacatgaaaaataaaggaggtgttttttctccttggaaaaagtatttactactgattaaagcaccttcattcacccatcttgctacttcgcgtactttactaagttgtggttctaaaaaagaagatttagcttggtatattccatcacgaggaagccacaaaaagattctacttataagaagttggatacttttaatggtgtgagaccttttgggaaaaaccatgcgggcttggcccaaagccgacaatatcacaccatgttaagagtatctttgcttcgttttagccaaacagtacaataagtcaatttttaaaccatgttgttactattgttggatttgcagaattcagcaaggctaactgttcctggaggaccgagtatagcttgcagcacagctaaagctattgagtgggatgcttcgtggtcgaacaaccttgatccttccggtagggctgcaatcggtgtacataactcggcttatgacgatcatctccccgatgttggtaatgggaatttggacacaacgatcatcgataatgtgccgaggatggtaggagtgggtgctgctgcagagctcacaataaggagctag
SEQ ID NO:6 - NtASN5-s, предсказанная аминокислотная последовательность
MCGILALLGCSDDSQAKRVRVLELSRRLKHRGPDWSGIYQHGDFYLAHQRLAIIDPASGDQPLFNQDKTIVVTVNGEIYNHEKLRNLMPNHKFRTGSDCDVIAHLYEEYGENFVDMLDGVFSFVLLDTRDNSFLAARDAIGITPLYIGWGLDGSVWISSELKGLNGDCEHFEVFPPGHLYSSKNGGFRRWYNPQWFSEAIPSNPYDPLVLRRAFENAVIKRLMTDVPFGVLLSGGLDSSLVASVTARYLAGTKAAKQWGAQLHSFCVGLEGSPDLKAAREVADYLGTVHHEFTFTVQDGIDAIEDVIYHIETYDVTTIRASTPMFLMSRKIKSLGVKMVISGEGSDELFGGYLYFHKAPNKEEFHVETCHKIKALHQYDCLRANKATSAWGLEARVPFLDKEFINVAMSIDPEWKMIKHDHGRIEKWVLRKAFDDEEQPYLPKHILYRQKEQFSDGVGYSWIDGLKAHAEQHVTDRMMLNAAHIFPHNTPTTKEAYYYRMIFERFFPQNSARLTVPGGPSIACSTAKAIEWDASWSNNLDPSGRAAIGVHNSAYDDHLPDVGNGNLDTTIIDNVPRMVGVGAAAELTIRS
SEQ ID NO:7 - NtASN5-T, последовательность геномной ДНК
atgacgtagtttagcttgttcgtccacggcataaaaacgcctataaattcggtggtcattttggttcatttacagaatcccacgatcatcttttgtgtattatcattttgccttctttacgtgttcttctctcccaaaatcctgcaaatctttaactatcttcccatatttctctccctctttctctttctttttgttatctactttactacttctattagtagagctttatccaattaaagtcatctatattggatatagtttttactactactttatcttgtttgggaaagaaaagaataaaaaattagagtgtctatattaagtataaaaaggtgaaaaataaaaaagtagtgatcaatcatgtgtgggatcttggctttgttgggttgtccagatgattctcaagccaaaagggttcgagttcttgagctctctcgcaggcaattcccctttcttttctcttttattttattactccatcatattattcaactataatttcaccttttttgtttttcaattaatatttgtctgcttttgctttcttcatttaatgttcttgcattgctctagttcctttatgaataataggctaaagtaatagagtgaaggagtagagtcggagcaacggtgaagctgtctttacatgatctatagctcacgggttcaccatagaagcagccaatggcgaagctagaaaattcaataagggtgttctttcctttgccagtgggctatgcaagggtgttcaaagtctatttttaatcaataacaagtaatattttatcttatagtgaaagcaaccactaatacttgcatttaggtaattagctgtctatcaagggaagttatgtgtggtaagtagtggcggagccagaattttcgttaaggggtgtcaaaatatataaaagtaaacataccaggaaattaaggggagtcaatacatagtatatatacatataatttatttttttacctagctatacagtatattttttccgcgaagtgacaccccttcctataaggtggctccgccactggtggtaagggtggtcaattgaacacccttcgccgaaaaattatactctggatggtgtaaaatattactacttgttactgattcaataaataaattttgaaaacccttgcatagtctagtagcaaagagtgttcaaaattttgaacacatttattgaattctctggtttcgccactgctgtctacatcacatcccttaaggtgcgatcttttttcgaatcatgtgtgaatacgggatgccttgtgttccaggctgcccctttaggctaaagtaatagagtacgtcttaattcttatgttgatcatatacctgaaactcggaactggcacgagagatataaattcagaatttatagcaaatttatcattatgttgcatttgttttaatgttcttgcattactctaatttttttatgaataataagctaaactaattgagtttatgttgatgtaactgacctgagtttggctacttgaaagttaaaacaagattaagacttgtgagttggcagcgattgtttggatacttgaacacgtttaaaacaagattaagacattatgatggatacattattcctaacctgggtgggacctaattctatggattcattattaaaatttcttggacaaaccatcaccatgatctgttcatgatgtttatggcaaaagccataatacactttatttctgtatcacaactactttatacacatgttatagagttttaaaattattttaatatttgttattccttgagtctcctgatatttgactccgcatgaaattcaagaaaaaaaaaagaagatttttaaaatttatagtcgaaaacaagccatcgatatatgtgtggttgttaataacctcgttaggcgtaaagtagaaattttaaagttaaactatttctaaataagaaaatatgatattgtttttgcatagactaataaggaaagtatgacaagtaaaataaaacaaagaagtatattttaatgattttttccgtgtcaaaaatgtcagtttagttgaacttgaactcattgaaactggtgaccagtggtggatgcaaaccttaggtacggaggtcatccgaacctagtatagttatatttgtgaaaaatcactaaaatttcaataattactagatatgaaaccttaattttaaaactacaatgagtgcagttttaaaatctttaaatgtcaaaccgatcaagattaaattatggatccacctttaaatgtcaaaccgacagtacaatagccttttgcaattgaaacaaatataaacattacatttattaaactagttttaattttggttactatttgattacataggttgaagcatcgtggaccagattggagtgggatatatcaacatggtgatttttacttagcacatcaacgtttagcaattatcgatcctacttctggtgatcagcctctgtttaatcaagataagactattgttgttacagtgagtgcctctaatttacccttttttttgtggacgaatgcgttgtttagattactttaattatggctgaaaatttgtatatttgattattgattgtgtgccaaatgttgcaggtcaatggagaaatttacaatcatgagaaacttcgtaatcttatgcctaatcacaagttcagaaccggaagtgattgtgatgttattgcacatcttgtgagttagtttactttactcatcaaccaacttaggctagtgaattcaaaggcggagctagaattttgagtatatgagttctggaatttaggacaagataagttactgggttcggatagattatttagacatattaagtagatttcttaacacaaatacacggcccgagccgaagctattgggttctgccgaacccgtagctagacttgtagctccgccattgagtgaattatataaggaatttcgggtgtttgggtaaacttataagctggtcaaattagcttataagcacttattatcaatttcaacatttctatccaaacacgtaactattcattcgattttagcacttgatgcttatcgactacttttaatcagttaaacgaacaggctcatagtgacaaactctagaaaatatggtattgtaaattagtagaagaatttatacgatgtctcactatcgatatgtattgatcagtatgaagaatatggagaaaattttgtggacatgttggatggggtgttctcttttgtattgttggatacgcgcgataacagctttcttgctgctcgtgatgcgattggaattactcccctctatattggttggggacttgatggtaagattttctatacaattttccaagtagaaattaaaactgaagccattccaatttgctattgtagaatcttgatatatgttatttggtatttcaggctctgtgtggatttcatctgagttaaagggcttaaatgatgattgtgaacattttgaagttttccctcccgggcacttgtactcgagcaagaatggcgggtttaggagatggtacaatcctcaatggttctctgaggctgttccatcaaatccttatgaccccttagttctgaggcgtgccttcgaaaatgtgagaaatgaatctaggaggtttccctctcgactttggacttagcactctaaaaatttgtctgtacaaacgatcttggctataaggagtggttgttaatttcgcaggctgttattaaacggttgatgaccgatgtaccctttggtgttctgctctccgggggacttgattcgtctttggttgcttctgtcactgctcgctacttggctggaacaaaagctgctaagcaatggggagcgcagcttcattccttctgtgttggtctcgaggtcagtcaatcaatcttgagtagaaaaaatttagggtcggttctgtgggtttaacttaactcattgcttttggctcgaaccatatatgcaaatccaaaaagatttaagatgtatacatataatataacactcattctgagcctactgacgctagatactggactgtgcctttggtcttgagacttttgtaatagaaaattgatattcaataccaatttctgttcttttgtgactaacagggctcaccagatctcaaggctgcaagagaagttgctgactatttgggaaccgttcaccacgagttcaccttcacagttcaggtttgtaaatcgcgatgctacttcaatttattcatgtgaggctcttctgttttcctactagaaagcaacatgtgtgcatgttttggcaggacggaattgatgctattgaagatgttatttaccatatcgagacgtatgatgtaacaacgatcagagcaagcacccctatgttccttatgtcgcgtaagattaaatcacttggagtgaagatggtcatatcaggggaaggctcagatgaactgtttggtggctacttgtacttccacaaggctcccaacaaggaagaattccacacggagacatgtcacaaggtaataaaacacatcggcaagtctgattctccttgtcattgtttgtctcgggttggaataaattgctataattcgagatatggaatgagttaagaaatgttaatcatgtttttccattcgcatgcagataaaagcgcttcaccaatacgactgtttgagagcaaataaggcaacatcagcatggggcttagaagctagagtaccatttctggataaagagttcatcaatattgctatgagtatcgatcctgaatggaagatggtaagaaaactgagctccaattgtcaatgagacgagaagcataactgaatatgcgcgttctctaaaacctgtttttcatccgtttcagattaaacacgatcaaggtaggatcgagaagtgggttcttaggaaggcttttgatgatgaggagcacccctatctcccaaaggtccacaaactcatatttttcctactgatttgccaataattatggatcaaaactaacatgtttgttctgttatgtgctctgcagcatattttgtaccggcagaaagaacaattcagcgatggtgtaggctatagttggatcgatgggctcaaagcacatgctgaacaacatgtgagcttccttaataaatccagaactaatgcagcgttatcgtccaatttttgtttgttttactatcaactaattttagtcatgttagtatgtttaccttaggtactatgtttttaaggagtttttagtcgtgtcaagaatttattcgctggtaaaaaatagattgaacttccagtattttctacttttattatgcatatgattggactgagatgaggttaaatggaacaacatggttgaggattaatataaccaattccaacttggttggaattgaggcgtcgtagttgttgttagatctcagaaacaattcttttctctcaggtgactgataggatgatgcttaatgcttcacatatcttccctcacaacactccaactacaaaggaagcatactattacaggatgatttttgagaggttctttccacaggtaatattagtgagagattaaaatctatagattttgtttatggctgaggtttaaatcatagatttcttaagctcaaaaactaaaaccatgtccctgctactgttggatttgcagaattcagcaaggctaactgttcctggaggaccgagtatagcttgcagcacggctaaagctattgagtgggacgcttcttggtcgaacaaccttgatccttccggtagggctgctatcggtgtacataactcagcttatgacgatcatctacccgatgttagtaatgggaatttggacacaacgatcatcgataatgtgccaaggatggtaggagtgggtgcttctgcagagctcacaataaggagctag
SEQ ID NO:8 - NtASN5-T, предсказанная аминокислотная последовательность
MTLKHRGPDWSGIYQHGDFYLAHQRLAIIDPTSGDQPLFNQDKTIVVTVNGEIYNHEKLRNLMPNHKFRTGSDCDVIAHLYEEYGENFVDMLDGVFSFVLLDTRDNSFLAARDAIGITPLYIGWGLDGSVWISSELKGLNDDCEHFEVFPPGHLYSSKNGGFRRWYNPQWFSEAVPSNPYDPLVLRRAFENAVIKRLMTDVPFGVLLSGGLDSSLVASVTARYLAGTKAAKQWGAQLHSFCVGLEGSPDLKAAREVADYLGTVHHEFTFTVQDGIDAIEDVIYHIETYDVTTIRASTPMFLMSRKIKSLGVKMVISGEGSDELFGGYLYFHKAPNKEEFHTETCHKIKALHQYDCLRANKATSAWGLEARVPFLDKEFINIAMSIDPEWKMIKHDQGRIEKWVLRKAFDDEEHPYLPKHILYRQKEQFSDGVGYSWIDGLKAHAEQHVTDRMMLNASHIFPHNTPTTKEAYYYRMIFERFFPQNSARLTVPGGPSIACSTAKAIEWDASWSNNLDPSGRAAIGVHNSAYDDHLPDVSNGNLDTTIIDNVPRMVGVGASAELTIRS
SEQ ID NO:9 - NtASN3-S, последовательность геномной ДНК
atgtgtggaatactagctgtttttggttgcattgataactctcaggccaagcgttcccgaatcatcgaactttccagaaggtttcttctttttattgctttccgattttctacagcattcttattcttctggggtgtcaatggtatgcatgtgttttggatttgctattcgtgaacttgttcgctttctcgtggtcttcgatttatttttgtcaatttataatcagcgttttttgtatggatttctcttttgaatctctcaaatatatgacatttgatttcctcgagaaaatttggcatttttatggttcttatgatatgtggttaacccctcgctctgcatcgcttcagcgctttaaaacacgaagtgatggctattagtaatactgctagagtggttattgaattacattaaataacgggttgctagtagatcaaattcagaatgacagtgaaatatctctggaaacttgaatccccgttcgattcgctggtaaaacctggctaaaaaatagctttatctcaaaggagagtatgtcactgtgaaattggtggtatgtgttgtttaaggcatgatatgctgctcttaggtgatgtcaagatgataataccatccttttcacactagactcagctttgtagcacttacatctcagttaatcgatggaggaaaatctagtctgctttggggcatattataccaaatgtcttccttttgttttctagtgttgtttctcaagaattgaaaaaccaatctacaatatagtgatataattaactagttaactcttcctgtgttttctgaaaatttccaataattgagaagtgtttactggtcaattagtaaagggagaacggaatatttttttcctctaaagaacttctctttttccatacagaatgactttctatacaaggcctaagtctaatttagtgtctgcgattaagcatcttctgggaacttcacacttgaaagaagtagaactaggagggggcaggtctagaaagataattgcttcagtcatttctcataaaaaacattgcttcagtcattatgtgcattcatatagagttggtcaagctgctaagctgagctaatagtatgatagctgtgactctcgacaaagtaggttgcctcttagggctttatcctttggctccgtaaagaatttctttgggagggctccagagtgggtaggattgttctcatggtaagctgtaacaccaacgaaacaagcctttgtagctgtggaagtccgagggggtggtggtggtggtggtggtgtgagagttagagtggtggaggagaggggaggagtgaagagaagtttatattttaaagaatagggccttgagaacttttagtagtgagcatgcattattattattgttattgttattattattattattattattattattattatattattattattattatattattgttatgtaattacttagctaatgaggtttttccaactgtttccaaattaggaaatgtgttgaaagcatagttaagtatataaaagcgtgttctttctaatatccttagcttttagagcagataatcacaaaatttatcatgataatatagcagacaatttttcctagttcaaatctcatgccacccttcataaaaaatttccacgtgcttggctggcatgttaggggacgtcttggcataattaaacaaatagaattgtccctttctaacagcttaagcttttagatgaggcggtcaacaattcaacaatttgcatttgtttgataagaagatattaaaagaaccttaaaaggtgtcagtatgtgaacaaactcaaggggtagactaactagaagtcaatttaaattttctgcttgtctgtgttttacttcttttgcactatagaaattgcatttccagcattccgaaagctgcttcctttgcatatagacttcacatttccagcgtttctgataagcgttcttgcttattcctgtccttctacttggttgattctcctctcacttagttcactatcttcttgcacgcattttcaaattgttttatcaattctcttgctttcttttcatctgaccccacaaaatacctcccaaaagcctttctgcttttccgagtctgaaattatctgcttattactgcctttcaagtattctgatgcgttgactgtttttttcccttgtagtaatgcatggttcgctgtccaatctctattttgttttttgtgaaccactattctttgttgcttctgtttcgtcatcctcagttccccatcttctcagtttgattttcctcatttaaaaagcgtgtgccccttttcttgtgctaccctctaaaaaatcagttgctgtcatctcgtttgatctgtcatgatcccagttgatgccacatgtttagttctttacctctggacaacttttacttcctgagaaccacgatttcttttcctgaagaaattcttagaaattggttatgtaaaggaaattcccagcaatcaccatttggcctctacctggcttgcttttcaacttaccgacacgtgttcatccccttttaacagatgctgaccattatcttctgcttgttggggcatacttcttacccatggtgttaaccctgatctttcgtatcacatttttcaactcttaattctcacatgtaaagtgcctatctttgcctctttagttgatcttgggttgggctgttaatccaaattctctgggctttgctttgaagtatgataagcggtagctttactacagtaccatattctacattgctacagacgtgaaattatcctgatttgtctttgatgtgatgtctggcaagtgtagattacgccacagaggacctgattggagtggattgcatagccatgatgactgttatcttgctcatcaacggttggcaatagtagacccaacttctggagatcagccactttacaacgaggacaagaccattgttgtcgcggtatggaaatatttattatattttttcttagatattaccggtacatggactttggacaatatttgtgcgttttgatcttctatagtacaaacttttctcgtaggataaatttcctctactttgaatggatcttaacaggaaaatcggaataaacaatttgtgatttcgaaatttttagttcttacatatttggaacatatgcggtaaagtaactaggcgattgactaggtgaatacggagaatttcctctactgttatcaatatttactcagctcatttgaaagatctctgggtcgtctagcagtacagctatttaggcaacttaaccaaatccccttcccatgtgtttcactttcttgacttattttatattttgacattttgcaatgccgaaaataaattgtttttccattttatgacctttaaaatgttgaggtgaagggctaggcggtactttaattgacttatcagcaacaatgtattagatacagagcacaaacttgtttgggtgcctaatttggtattttttgagaacgatggagtaaatggaaatatgaattagcaaaatattgaaatagtagatctgaaaaatgaatgactgaatttagaggaggaccttttcttctttccttgctgaagtggttataagagaattatgtgaagaacatgtaagctttagggtagtgttacaccataacagtcatgctttccctcaactattagtgtagctgatgaatgctttcagaaatgtggctgtttttttgctgcgtattttatgtctttcatcatctcaaaatttattgtgttttaacactgcttggaaggcctagagtcttccgaaaatagtaggttgagaaaacatgagtaagcaaatttcttcctgctggcaattaatcgttttatatgaggtgaagccacacaatctaacatggtcttagacataaaaatattatttttgacatagagcagacagaaatcttgggtctagttgctatctcccttcctaataagatgccctctatcctcattgctatatcccaaaacagattcacactagaatgaataaatctctccttgcagccacatttatattaataagtagtgtaacaccaaaacatttgtgcttctgcaaagtctgttgaagtggacgattaagtaataaattgtgtgagtttgactgaagagaaagtaataaattgggtgagcttctcacatttttcaggataatgtcaattcatctgattgaactttttgaaggacatctccttttctaaaagtttttgttcacttcctgaaaatggttgctttcccaaatttattctcctgtattttccttgtgctttttttttttaccaggatgactctggccttcccttttgcaaatagtgacttcattaccttttaacaaaagaccaaatagcctttgtatgcttgaaatcgaagcagaatttacggctcaatttctctaggagatgtgtagcagtaacataaaacgtggttaaagcaacttgtaagcactgccactgtccaatttgacatgactctattactatttagagtgtcaaatagtgtttcttttgactaccaatttttcaatttcttttttataccttttttctttgactgtgacttatagtatattttatgtagtttcagaatatatattttagtttttaaagaattgatgtctgaaagtacagccaattagatggtttcaccctcgtactttaaatcttataagttgtcttaggttaaaggaataatatgcatgattacattgtttggattgatattcagaagatcaaatgtgccggacaaaagcttgcagaagatataattctgtcattttgttgtttagataaactacccttgtcaaatacctaaaactaattcatacttttcaggatccttcttgattggactactctgatcttcttccatatattgatatcctattctttttgcttttaggttaacggggagatatataaccatagagacttacgggaaaagctgaagtcccaccagtttcgaactggcagtgactgtgaagttattgcacatcttgtaagtactttcagaatatatttagataactatacatgacttctcagtttctgtgcgcagaatgatttcaaaataccatgactcaaatttttcatcaaaagcgaaaaaggtggattgtagtaggagaatgagattttttgtcataatttctttcttatggtacttgaactccgcaatctacctatcgcagtatgaagactatggagaagacttcgttcacatgttggatggaatgttctcctttgtgcttcttgacacccgtgataaaagtttcattgctgctcgggatgcaattggcattacacccctttatatgggatggggtcttgatggtatgtaattgttccatttcataatttttggtatattgtccttttaataaatagcccgtgagctatatgaagttcctttgttgtttccttgttttttatttgactgatgtatctttgttgttttgcctactattaggggtgtcaatggttcggttcggccggttattttataaaatttgtaccataccaatttttaggttattctattatgtataaccaaaattagactttttgaaaccgtcccaatcatgtcggtttctcttcggtatcggtacggttcggttaattttcggtattttttaatatcatgtaaaattcaccagtagaagtagaattgcaataacatacattcttttataggacttagcaaaactctctagatatttttactgtttaaagggtgatgaattaaaaaaaaagaaaagaaagatggctagaatatagatccatcaactattatacaacagcgtaaaagaaatcaaacaaagacaaagaaaatattaatcacacgagttgaaagatataccaaggtaggactcaagaataaagtctatagaagattaaatattcaaaaagataaatctaaattatatgaaaggaaacatattcaattcattgtagtttgctactcataatcgctagaatactttgtgtcttgctaataaagatacttgaaataatttagtttaagtagaagtagcataataggttttatgaattagtattttgagtttaattacttgttggcttgtaataattttcataattccaaggctcaaagaaaatttattgcattattatttttaaacttactaaataaatatattttctacatgtaaaatttattcggtatggttctgtattttttcggtttatttttataaaataaaaaacctaccctaattatcggtgcggttgtagatttatataaaaacctacggtttcttaaaaagaaagctaaaaatcggttcggtgcggtacggttcggtcggtttagtcgattttcgaatatccattgacacccctacctactataatgctggtctcatgctgttgttacaaacaaatgatgtataatagtgcatagtacatgttttcttgccgctgtagatcaacaatgtaccaagaacatctgtcttagctagttcaacctaatcttcagttatctatcattccagtgaataacacatatagagttaggccgacattacctaaggccaactatacaacattgtgaggtgcgactaaggaactctgaacactgatcgggagaaaaagcaacttatccgtgtttgttttgaagtactatgaggaggcagcaaccttgctggatttatcacttgacagtaaatgtcaatggttgaggaacatttttaatgaccggaggagaaacagagaagagaagaaataattgattttaaaaaagcagaatagaaaattttgtgaagtcgtatgttaatttcaatgaagaggatcaacaggtgcaatatttgacttggcacctagcatgtagaagtttctggttggggttgagcatggtaggcactgttgcttttacaactgtcattagagtcggaaacatattgtattcttgagggagcgttgctatggataaaaaaggcctatctactacttgatatttgggcattcagaactagttgtctaggaggaagaaaaggatttaaaatggtgagtattgtaggctttctttgccgtccaggatgcctgggctagttccctgaattgggatactggtggaggatttcctgaaattgatcatggccctgatatgttaggggataagaacattgaaggatcgaaggaaggtgccggcgctaagttttgctagaaatgaacagttccattatggcttttgtcaaagatgggaatggaattggccccaacttctaggcatctccatccatcgtcaggtgaactaaagtaaggttacactggtagaatttttcattatttcatattaagtaaggtgaagccgtcaacaatcattgcgtgagaaggaaccaggaacaaaatctagaagatgtctactaaagtgctaatagataatctgcaaaatttagcacggtaaggctagtgtagatctggttcaaattttaaattggtccaaaatgcgttaataaatgtcgggcctcgttttggtcccccgaaaggttgctaaataaagacttgacaaagaaacaggtgaaggagaagcccaaacatttgaacaaactacacctagcctaaacaaagtgattgtttgcgaggaggaaaagcgagcatgcggagagagcctcaaacacaaaaaatggcctagtcttgtaatgagtctagccaaatcatgtaagctccgactttcaagagagatcaacagaaaacttcgtcaatccggataaatcaagaaaattatgataatggaaaggaggctggacattctactgccgacgcaacccctggttttggagtcagtgagacaagtggaactaaagaagaatatggatcaaaagagtttatccagttgacacctgatagactcactacatcagttttcagttcaagaacatttcaattcttgctgataaagatagttgccgataaagataggcaaactgttacctaccaatcaaaggaggttgtttcaattgagttagaggtatcaaaggaggagccagtatatatagagcaagttttgatgaagggagttgcctctatttgagaccaccaggtactcaagtgcttcccgtagttcctaggcattttatgatggatatgatagagaaatgtcttattgacaagaatatactagggctacagggataatagggttgggaaggtgaggacttctttggttgctgtcaggaacttggaaacaaagggagaaaggggttggaatcaaaggttttgatttcgtttttgatttataaggaaataatgggtagtggatggagtcgaggataggaaagaggtttaccagttgcaccattgaatatgtaattagagacactatcctcttgttagggaacacgactatcacaagcacgtaagggtgagtgaaagctccgacatggacgtggtttattcgggtatgttggcacgaagccttcgtgtctaggtttggtgttgggttaagtgcaactccttaggaaggcaaataaacttgggacagatttgatagttgtcggaagttgcacaatgatgatattttctttatccaaacgctacgaatgattgcacatatagtaactagagttatttctcatttgagcttttatatgtaagaatcaaataaaatgattgatgaaatgcatgaaagctcaaatgagaaataactagttgtgagaatgcacgggggaaaaatatattatattgattaaagtgttgtacaaccctatttagatacagtaattacataataataggtatctacttcccgatgtgggacactagacatgactaactacttaacaatccccctcaagccggtgcatataaatcatatgtatcgagcttgttacagatgtaactaatacgagaaccagtaagagacttagtgaaaatatctgctagctaatcattcgactttacaaactttgtaacaatatctcttgagagtattttttctctaacaaagtgacagtcgatctcaatgtgtttagtcctttcatagaacaccggatttgacgcaatatgaagagcagcttgattatcacacaccagttccatcttgctgatttctccgaacttcaactccttgagcaactgcttgatccaaactagaatcacacgttgccacaaccatggcccgatattcggcttcggcgctagatcgagcaactacattctgtttcttgctcttccaagagactaaattacctcctactagaacacaatatccagacgtagaacgtcaatcagcaggtaatcctgcccaatcagcatctgtgtacccaacaatctgctcgtggcctcgatcctcaaatcgtaaccctttgcctggagctgactttatataccgaagaatgcgaacaactgcatcccagtgactatcacagggagaaaccataaactaacttgcaacactcaccggaaaagaaatattaggtctagtcactgtgtggtaatttaatttgccaaccaacctcctctatctcgtaggatctctaagatgctccccctgtccatgcagaagcttagcattcggatccatagaagtgtcaactggtctacaacccatcattccagtctcctcaagaatgtctaaggcatatttccgctgtgaaataacaatacctgagctagactgagcgacctcaatacctagaaaatacttcaatctgcccagatccttagtctggaagtgctgaaagagatgctgcttcagattagtaataccatcttgatcattgccaataataataatatcatcaacataccactcaactttggcatttctatctcaatttggctggaatatcctggacaatgcccaataacactgcgtctcttctcagtagctggaacaatggaggggcaaatattaaacagaagaaatggtggaaagtagttcctgcttgtatctggtgggtcatgtggaaggaaaagaatgcagagtttttgaagataggcctagctcttagcctcttaccagaaaattaagcttaattgttttttgttatttcatttttggtgcaaagagagctatagggaggatgtacattctttactagatttgctagaagaaatgtaatcgattagaataggcaaggcagttgcagatcttttggggtctgattgtggcttacactatctcctgtaaatatttttaatcttttatataatttgttaccattctaaaaaaaaacataaaccaccagataaatatacagattaggagcagaatgtcgataaagcacagagtgatccgcttcactacgagtcatgccgaactcctgaataattatgctgaacttaccaaaccaagctcgagggaactgtttcaaaccatatagtgacctgcgcaatcggcatacaagaccactagacttcccttaagcaacaaaaccaggtgattgctccatataaatttcttcctcaagatcaccgtggaggaaagcattcttagtgtctaactgataaagaggtcattgacgtacaacaaccatgaacaagaagagacaaccgatgctactttagccacgggagagagtgtcactataatcaagcccaaaaatctgagtgtatccttttgcaacaagacgagccttaagacgatcaacttggccatccgggctgactttgactgcataaacccaacgataaccaacaatagacttacttgaaggaagaggaacaagcttccaagtgccactcacatgtaaagcagacatctcctcaatcatagtttgtcgccatcctggatgagatagtgcctcacctgtagactttgggatagaaacaattgacaaagatgatataaaagcataatgaggtgatgacagacgatgataacttaaatcgacataatgaggattaagattaagagtggatcatacacctttccgaagtgcaatcggtgtactaggaagagacaagtccgcagtaggagcagggtcaagtgtaggatgtgaatcagttgggcctgatgctgggtgcggacgacgatgatatgtcaagagtggtgttccggtggcggggaatctaggaggagccacactagactccccaacggttggaatgggtaagacttatgtggcggaaggtgaaggaggagctatagtaagctctttaaaggtcggtataggtaagacctcagatatatcaaggtggtcagaagaggtaaagaaaggtttagactcaaaaaatgtgacgtcagatgacataaagtaattacgaaaatcaagtgagtaacaacgatatcccttttgaacacgagaataaccaaggaagacatacttgagagtacgaggagctaacttatctttcctaggggctaaattatgaacgaaacaagagctccccaaaacacgaggaggaacagagtataagggtgactgggaaacaatactgcatacggaatctgattctggatgggagatgaaggcattcgattaaccaaataacaagctgtgagaactgtatcgcctaaaaaacgcaacggaacatgagattcaaagagaagtgtgcgagcagtttcaatgatgtgcctattctttctctctgcaaccccattttgctgaggggtataaggacaagaggtctgatgaataattccttgagaagtcataaagtgctaaaattgagaggataactattctaaggcattatcactgcgaaaagtgcgaatagaaacaccaaattgatttttaatttcatcacaaaaattctggaatatagaaaacaactcagaacgatctttcattaagaaaatccaagtacatcttgaatgatcatcaatgaaactatcaaactaacgaaatcccaaggttgagttgactctactatgaccctatatatcagaatgaactaaagaaaaaacagactatgcatgactctcaatactacgaggaaaggaggctcaggtatgtttcccgagctgacatgactcacactctaatgtagataaactaggcactatcttctgaagcttggatgtcctaaacatctgtgaattaggtccggaggatctgtaactagacatgccttggaggaattgagtgggttaaggtagtaaaggccttctaattcaagtcctgttccaatcgtctgtccgtactgcggtcctgcataataaaagaatcattaataaaatatataccacaatggagggcacaagtcaaacgactaacagatgcaagattaaaaggacaaccagggacataaagaacggaatctagagtgacagagggtaggggattcgcttgtccaactccttttgcttcagtttgagacccgttggctaaagtaacagtggaaagagactgtgaatacgcaatatttgacaaaagtgatttattaccagagatatgatcagaagcggttgagtccacaacccattgtccaagagtactagactgggaaacacaagcaaaagaattaccagcaacaaaagcatcagtctgagcaacagaggctacttgtggagatgtctgcttacttgctcgatactgaaggaacttattatactccccttccgataaagaaaatacccggttacctatagtctcggtctgagcaacataagcatttttgggtggacgaccatgtaaagaatagcacacgtcacgagtgtgtccaagtttatgacaataagagcacttgggtctagatcttccaaaacgacctcctcctcgtctattctccatagtttgagatgcccgattgttcacttactgggatacgagaacagatgagtcaggtctctgtgatgagctcactgggtgacttggtgctgcagcaaggcgaagtaatcgagagaataattcatcaactgtggggacagtcggactagccaaaatctggtcacgtactgaatcaaggtcattagggagtccatcaagtgtaagaactagaaacatcttctgtcgttgctcttgttgctttttaatactagcagaaactggcatcaacgtctcaaattcttccatgattgcctgtacttgtcccctagtaagtagacatatccaattcctgtttcttcaagcttgtcattcgcgatattacatcatagaaacgagatatgtcattagtgtataaattacgagccttttcccaaactaaataacatgtctggaatggacggaacaagggcatcaacttggaatcaatagatcgccacaggatactacataactgagcatcgaccttcttcaaaagtgttttggccttttcatcaccttcgctagccttttttgttaaatgatcttgaactccttgacctttacaacacaactcgacagacgaagcccaagctaagtagtttgaacttctcattaaaggttctgaggtaatcataatagcagaacttccagaactcgtgtttttagacccaaatacatccactcccaaagacattattggattgaaaagagatctagcaaattagcaccaaataaaacaaagaatcaactgtggttgcccaaaaactgccggaaaactactgtagttgccggaaaattttcaaagtgctcggaatcaaaaaataaaaatatgggcaggctcggaatggtagagcgatcagactaacctaaaggagtttttctgaaaaaattgacggaacgggctccatgcgctggtgcgtggagtagatctcgccggagaagactgtctccgatcggcgcgtggcggtgcgtgaggcggcttatgacggaggtgttcgctggggtttggtcgccggaagttgggggaccttgtggtggtgttggtttttgcacaacaccgatggaattggttttgacgaaaaatagccctaaaaggtcaccggaattgaagcacggcgacggctggttttttttttccccggatgttttctcactgccgctctgataccatgtgagaatgcacgggagaaaaatatattatattgattaaagtgttgtacaaccctatttatatacagtaattacataataataggtatctacttcccgatgtgggacactaaacatgactaactacttaacactagtaacctcatggttctcatcctacttcattgaccacttggccacatccttgggtgcctgcactgtagttgtagctgtaatgaagatagttttttttcttttctggatactcttatattgtgacaatttttctctatttgctgttaatggatgaggttatgtgactctccggtggtgagaatgttaatgaacatcaagctcacatgtcatttcattttctgattgatcttgctcttcactctttacgagtttaatgtgttagtccttccaagttctgtccatctatctttctggtgtgcttaacccatgtatattggaccacactttgtattcatgctgattgcaacaaagcctgactgatcctgaactcctcctttatgccttaggtagaaattttgtgtgtttagtgttacatgtttcgcttcaaaggaaaaagggagaaacttaaggtagaatcctgctccttgacctttggatccttgcacattgaagtggacagacatgtttgctttgctttgatgagttgttttagtagtcggtctaaacaactgctgatgtattctgctttattgtaagtgtctataatttttatctttccacaatttgctttcagctctttctgcatgtagagaaaataacagaagcttctatttgatcatgtaatattgactgcgcaagttcagttttaaacaactttggctatgcatatgcaagcaatgacctgaaataaatattattttttatgcagggtctgtatggttttcttcagagatgaaagccttaagtgatgactgtgaacgatttgttagtttccttcccggtcacatttattcaagcaaaaatggtatgcccatacgaagctcatttctgagcttgtgcttcatttatgttgatagttgatcatgtttcctatacgatcagcgtacttttgctatttatttcagatttactttaggaccctgtcggaaacagtcttgaaacccttattgatggtgtaattgttatctcttgatcaaccaaaggagtaattacatgtaattattacaaatttgcaataaaataacttttaggataaccagtggagcttaatgaatcttttaggatatttgggatgctagagtgatgtacttttgataatgtcggttaatataaaatcctctgactttgtactgatgtactggctgagcagattttatattactttctagtccctctgcaaataaaaaaattgtgagactgcaatataacaccattttttaatgcatttacaatgtttaaacttgatgtagcagtgaagaacatgtcgaccttaaacagaggttctatttgtcctagataaaggaacatcagcgtaatgaaacattacaggagagaagctctattgagtttttgttaatctagagggagcccgtaaatagaacaccaattcatctgttatatgacttctttttcattttggaacgttttaaaagtttgacaacattttttgatctccgtacactaaactttttgatgtattaaactaactatttccactactttaatattttcttccttaaacaactcatgttagcatcttaaactgtgtgtgttcaataccctcataaaaaattaaacgagtatttttttatatttgcttatgcccttggagattgggatggggaagttcatcatttgaggtgaaaggtaaattcttttggatggacaaaaagtgaacctcgtctttggcatggagggactatttaaatcgtcatcattaccaaagtcatcctttcaaatgtttactttcaagaactattcatgctatttctcaagtatattatgggtgcaggaggactcagaagatggtacaacccaccatggtactcagaaaccattccttctactccatatgatcaccttgtcttacggaaagcttttgagaaggtcattctattttggatgtcgcttgtttctgagttgtacataatatttaggtgccactttgttatattttgaacttcatcaaaagcctttggttttttgaggacacatatccccttgactgactgggcgctgaatttccttgatgtattgtgcttattttgattgtcctcttctttgtaatactcttgcctataaataaaccttctttatcaaaagaaaatgataaaatgttaataacactttgtgatatatgttgaaggggagtcttggcataactggtaaagttgctgccatgtgaccaggaggtcacgggttcaagccgtggaaacagcctcttgcagaaatgtagggtaaggctgcatacaatcgacccttgtggtccgacccttccccggaccccgcgcataacgggagcttagtgcaccgggctgtcctttttttttgcatagactttacatgatattttatttcaatctttatcaattcttttttaccttcacatgcaaattatactaccgtatgtgaaatatgcaatagtttttcccgcaattttcatctttcatgtacctctctctactatccttgtattactactgatttggtgcattagatgttaattttattcccaacctttactttatgcaggctgtagtcaagcgacttatgacggatgtaccatttggtgtgcttctctcaggtggactagattcttcacttgttgctgcagtggccaaccgttatttggctgatactgaagctgcgcgacaatggggatcacagttgcataccttttgcgtaggcttgaaggtggtgatgctcccttttgtaaacttgctctgattagggaattgctagctgtatcatcccaattcacatggaagttctagaccattattgctgtttcttttccatatatagggattttctgaaaatagtaaaaaggaaatggaatttctcattattaaagataccaaatgcctttttgggaaatgatcttacagttgaatctgaaatttccttttgacattgtgattaattctgatatgaatttggtccatcctggctttccttctcttgattgtagattttttttctctttttgcaattatatgcctttgtatttcatatctcttattatgctattattttattatgcatttttatggtactaatatatcggctcctgttgctttttttgagccgagattttgtttatttctgctatcacgtcctcctgaaaaagtgctactatcttgtattcatgcgggagtttgaaatttgcaagtgtataactttacaaaatgattcgaactgatcacagttcattgctaataaacttttataaaaagcaggtattgttgtgatataatgtgacacttaaataaatttaaacctacatcctttcgaaatctctaatagcttcactaatatttcttgtgctaaattcttccaacacctcttatcgatgccaaatttggcagaccttagtgcaattgaaagtgtttagtgttctttatgcagtaggtatgttattgtccaaaatgacagaaataaagttagagatttgttgaaaagccacttggtgtgattgagcagttcggagatgagtgatttatccatctttttatacttttagtgttttgttttcttttatattgttttctcttttgcttttgtcattgtaattaaggttgtcttttgtttgactttagggttctcctgatctgaaagctgccagagaggtggcagactaccttggaactcgtcaccatgagtttcactttactgtgcaggtaactttcttcataaagacatcctatgctgattttccttataggagtataactgcttaagtcaaattggctatctacttctttctattgctacatcaggaaggaattgatgcactagatgaagtcatttatcatgttgaaacatatgatgtgaccactatcagagccagtacaccaatgtttctcatgtctcggaagataaagtccttgggtgtgaaaatggttctatctggtgaaggttctgatgaaatttttggcggttatttatatttccacaaggcacccaacaantttacacaaaaatgacacaagttatgcagaaaaaatagtcctcaatgtcatgggagaccagggttcaaatctgaggttaaaaaaaatactaggtgaactcttctaatccgtttaagccttggtagatagagttaactagtatatgagctagttagagctagcaggtacctggtagattagtgcaggtgcacacgagctgactcaaaaccaccattattaaaaaataagaaagttttcaatgttgttccaccgtgaattcatgatgtgccgactttcctgtgtttatctctggatagatgaagagtgtttttggatcttgattaattctgtttggcccctttgaggttctgcaagctatttctagactcaaacttgctgcgatctatgttaggtctagagaatatctgccaaataaaacaatggaaggtacactcaagtattttatctccatattacaagtcagtgttagttgaatgggtctgatatatgggacagaaaatggcttcatacccataacctcattaaaatgttgctgatgtgctgatctcatctgtacatctagtataatcctaattacaaattaatgcatctctttgcggtgactatagtatgctgaagaacttttcttgaaacattgcagatcagacctgatctcggaagaatagaaaagtgggttctacgcaatgcttttgacgatgatcagaatccttatctgccaaaggtttgtttgttttcttgtatcaggtccaaaatcaaaatctttgtttattcatttgcataaacaggcagtagactcccaatacattgtgtacacttatgcacaaatgtatgagatgtgtattatatgtgtcacatataagggtatttccacatggtgtttattgatctagagaaagcgtacgacagggttcctagggaggtcctatggagatgcctagaggttaaaggggtcccggttgactgcattagggtgattaaagacatgtatgatggagctaagactcgaattaggacagtaggaggcaactccgaccattttccggttgttacggggttgcatcaagggtctgcgttcaaccctttcctatttgccttggtgatggatgctataaccgcatcatattcaaggggaggtgccatggtgcatgttatttgctgatgacatagtcctaattgacgagacacgacgcggcgtcaacgagaggctagaggtttggagacatgcccttgagtctaaaggtttcaggctgagcaggacgaagacggaatatctcgagtgcaannnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnatcgtatatgggtagggtggatgaagtggaggttagcaacgggagtcttgtgtgacaagaaagtgtcaccgttactaantctcgagtgcaactttggggccgagccgatggaagcatgagtggaagtgaggctcgactcacaagtcatccctaagaggggtagtttcaagtaccttgggtcagttattcaggggaccggggagatcgacgaggatgtcacacatcgtataggggtggggtggatgaagtggnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnnataggggtagggtggatgaagtggaggttagcaacgggagtcttgtgtgacaagaaagtgtcaccgttactaaaaggtaagttttacagagcagtggttaggcctgctatgttgcatgggaccgagtgttggccggttaaaaattcacacatctagaagatgaaagtagcagagatgaggatgctgaggtggatgtgcgggcatacaaggatggacaagattaggaatgaagttattcgagagaaggtnttgagccgagggtctcctggaaacagcctctctgcccctcggggtaggggtaaggtctgcgtacatactaccctccccagaccccattggtgggattatactgggatgttgttgttgttgttgttgttgttgtacatataagggtattatatgtacacactcctcccacctttcacttatatattcacgtaaaacacatgcatgtgtaaaacctatatatgtgtgtgtgatatatataataaaatgatgtttgaaaagacattatgatcaagtagttgatcttcatctgctttccgtttttagcatatcttgtataggcagaaggaacagttcagtgatggagttggctacagttggattgatggtttgaaggatcacgcaagcagactggtgagctttttagttgttttctcttccctcctcttccattctttatcccttcctttgtggttttgttgaacacatctcgtatatttggtccaggtttctgattctatgttagcgaatgcaagtttcgtttacccgcataacacacccacgacaaaggaaggatactactatagaactatttttgagcgatatttccccaaggtgggtcttagatagtcttcttttccttgttattcatgttcaatttttgatattttgtcccaaacaatgttttctttttcttccttctgttaacaaggtcgcttctatcttcatatttgttgataaaggtaatcacattgataactattatttggagcagaatgctgcgagggaaacagttccaggtggtccaagtgtggcatgcagcactgcaaaagcagtagaatgggacgcagcttggtccaagaatctagatccatctggacgagctgcactcggtgttcatgcagctgcttatgaggatgcatcagaggttaaaaccacagtcgcaacagatacccctcagaaacttgaagttgataaagctgcagtagctgtttga
SEQ ID NO:10 - NtASN3-S, предсказанная аминокислотная последовательность
MCGILAVFGCIDNSQAKRSRIIELSRRLRHRGPDWSGLHSHDDCYLAHQRLAIVDPTSGDQPLYNEDKTIVVAVNGEIYNHRDLREKLKSHQFRTGSDCEVIAHLYEDYGEDFVHMLDGMFSFVLLDTRDKSFIAARDAIGITPLYMGWGLDGSVWFSSEMKALSDDCERFVSFLPGHIYSSKNGGLRRWYNPPWYSETIPSTPYDHLVLRKAFEKAVVKRLMTDVPFGVLLSGGLDSSLVAAVANRYLADTEAARQWGSQLHTFCVGLKGSPDLKAAREVADYLGTRHHEFHFTVQEGIDALDEVIYHVETYDVTTIRASTPMFLMSRKIKSLGVKMVLSGEGSDEIFGGYLYFHKAPNKEEFHQETCRKIKALHLYDCLRANKSTSAWGVEARVPFLDKEFVNVAMNMDPESKMIRPDLGRIEKWVLRNAFDDDQNPYLPKHILYRQKEQFSDGVGYSWIDGLKDHASRLVSDSMLANASFVYPHNTPTTKEGYYYRTIFERYFPKNAARETVPGGPSVACSTAKAVEWDAAWSKNLDPSGRAALGVHAAAYEDASEVKTTVATDTPQKLEVDKAAVAV
SEQ ID NO:11 - NtASN3-T, последовательность геномной ДНК
atgtgtggaatactagcagttttcggttgcattgataattctcaggccaagcgttcccgaatcatcgaactttctagaaggtagaatttttttaaaaaaaa
aattgcattccaatttttctacagcattcttattctttggggtgacaatggtatgcatgt
gttttggatttgctattcgtggacttgttcgctttctcgtggtcttcgatttgtttttgt
taatttataatcagtgtttttatatggatttcttttttgaatctctcaaatatatgacat
ttgatttccacgagaaaatttggcatttttatggttcttatgatatgcggttaacccttc
gccttgcatcgcttcagcgctttaaaagacgaaatgatggctattagtaatactgctaga
gtggttattgaattacattaaataacagattgctagtagatcaaattcagaatgacagtg
aaatatctctggaaacttgaatccccgttcaattcgctggtaaaacctggcaaaaaaata
gctttatctcaaaggagagtatgtcactgtgaaattggtggtatgtgttgtttaagcttt
aaggcatgatatgctgctcttaggtgatgtcaggatgataataccatccttttcacacta
gactcagctttgaagcactttcatctcagttaatcggtggaggaaaatctagtttgcttt
gggcataatataccaaatgtcttccttttgttttctagtgttgtttctcaagaattgaaa
aaccaacctacaatatagtgatataattaactagttagctcttcctgtttttctgaaaat
ttccaataattgagaagcgtttacaggtcaattagtaaagggagaacggaacattttttt
cctctaaagaacttctctttttccatacagaatgactttctatacaaggcctaagtctgc
tttagtgtctgcgattaagcatcttctgggaaacttcacacttgaaagaagtagaactag
gagggggcaggtctagcaagataattgcttcagtcatttctcataaaaaaacattgtttc
attcattatgtgcattcatatagagttggttaagctgctaagctgagctaatagtatgat
agctatgaatctcgacaaagtaggttgcttcttagggctttattctttggctccgtaaag
aatttcttttggaagggcttcagagtgggtaggactgttctgatgataagctgtaacacc
aacgaaacaagcctttgtagctgtggaagtccgagggggtggtggtggtggtggtggtgt
gagagttagagtggtggaggagaggggaggagtgaagagaagtttatattttaaagaata
gggccttgagaacttttagtagtgagcngggggggggggggggcagaggggaggagtgaa
gagaagtatatattttaaagaatagggccttgagaacttatagtagtaagcatgcattat
tattattgttgttattattattattattattatttttattattatgtaagatgtaattac
ttagctaatgaggtttttccaactatttccaaattgtgaaatgtgttgaaagcataatta
agtatataaaagcgtgttctttctaatatctttagctttaagaggagataatcacaaaat
ttatcttgataatatagcagacaatttttcctagttcaaatctcacgtcacccttcataa
aaaatttccacgtgcttggcttgcatgttaggggacgtcttgacataattaaacaaatat
aattgtccctctctaacagcttaagcttttagatgaggcggtcaacaattcaacaatttg
catttgtttgataagaagttatattaaaagaacacttaaaaggtgtcagtatgtgcacaa
actcaaggtgtagactaactagaagtcaattcaaattttctgcttgttctctctgtttta
cttcttttgcactatagaaattgcatttccagcattcccgaaagctgcttcctttgcata
tagacatcacatttccagcgtgtctgataagcgttcttgcttattcctgtccctctactt
ggttgattttcctctcacttagttcactgtcttcttgcacgcattatcaaattgtgttct
caattctcttgccttcttttcatctgaccccacaaaatacctcccaaaaagcctttagtt
tctgcttttccgagtctgaaattatctgcttattactgcctttcaagtattctgatacgt
tgaccattgttttccccagtagtaatgcattgttctctgtccaatctctatttttttttt
tgtgaaccactattctttgttgcttctgtttcgtcatcctcagttccccatcttctcaat
ttgattttcctcatttaaaaagcgtgtgccccttttcttgtgctaccctctcaaaaatca
gttgctgtcatctcgtttgatctgtcatgaacccagttgatgccacatgtttagttcttt
acctctggacaacttttacttcctgagaaccacaatttcctttccttaagaaattcttag
aaattggttatgtaaactgtaaagggaaaatttcccagcaatcaccatttggcctctacg
tggcttgcttttcaacttaccgacacgtgttcatccccttttaacaaatgctgaccatta
tcttctgcttgttggggcatgcttcttacccatggtgttaaccctgatctttcgtatcac
atttttcaactcttaattctcatcttataaagtgcatatccttgcctctttagttgatct
tgggttgggctgttaatccaaattccatgggctttgctttgaagtatgataagcattagc
tttactacagtagcatattctacgttgctacagacatgaaattttcctgatttgtctttg
atgtgatgtctggcaagtgtagattacgccacagaggacctgactggagtggattgcata
gccatgatgactgttatcttgctcatcaacggttggcaatagtagacccaacttctggag
atcagccactttacaatgaggacaagaccattgttgtcgcggtagggaaatatttattat
atcatttcttagatagttaagtattttaccagtacatggactttggacaatatttgtgcg
ttttgatcttctatagcacaaacttttctcgtaggataaatttcctctactttgagtgaa
tcttaacaggaaaatcggaataaacaatttgtgatttcgaaatttttagttcttacatat
ttgaaacatgtgcggtgaagtaactaggcgcttgactaggtgaatacggagaatttcctc
tactgttatcgatattttctcagttcatttgaaagatctctgggtcgtctagcagtacaa
ctatttaggcaacttaatcaaatccccttcccatgagtttcactttcttgacttgtttta
tattttgacattttgcaatgccgaaaataaattgtttttccattttatgacctttaaaat
gttgaggtgaagggctaggcggtactttaatcgacttatcggcaacaatgtattagatac
agagcacaaacttgtttgggtgcctaaattgatatttttgaaaagtcgatggagtaaatg
gaaatatggattagcaaaaatattgaaatagtagatctaaacaatggatgactgaattta
gaggaggaccttttcttctttccttgttgaagtggttataagagaattattggaagaaaa
tgtaagctttagggtagtgttgcaccataacagtcatgctttccctcaattattagtgta
gctgatgaatgcttttggaaaggtggctgattttttgctgcgtattttatgtctttcatc
atctcaaaatttgttgtgttttaacagtgcttggaaggcctagagtcttctgaaaaaagt
aggctgagaaaacatgagtaagcaaatttcttcctgctggcaattaatctttgtatatga
ggtgaagccacataatctaacatggtattagacatataaatattatttttgacatagagc
agacagaaatcttgggtctagttgctatctcccttcctaataagatgccctcattgctat
atcccaaaacagattcaccctagaacgaataaatctctccttgcagccacatatatatta
ataagtagtgtaacactaaaacatttgtgcttctgcaaagtctgttgaagtgggcgatta
agtaataaattgtgtgagtttgactgaagagaaagtaataaattgtgtgagcttctcaca
ttttcatgataatgtcaattcatctgattgaactttttgaaggacatctccttttctaaa
agtttttgttcacttcctgaaaatggttgctttcccaaatgtattctcctgtattttcct
tgtgctttatgttgaccaggatgactctggccttccctttttcaaatggtgacttcatta
ccttttaacaaaagatcaaatagcctttgtatgcttgaaatcgaagcagaatttagggct
caatttctctaggagatgtgtagcagtaacataacacgtggttaaagcaacttttaagca
ctgccactgtcaaatttgatatgacgctattactatttagagtgtcaaatagtatttctt
ttgactaccaatttttcaatatcttttttataccttttttctttgactgtgacttatagt
atattttatgtagtttcagaatatatattttagtttttaaagaattgatgtctgaaagta
cagccaattagatggtttcacccttgtacttcaaatcttataagttgtttaggttaaagg
aataatatgcatgattacattgtttggattgaaattcagaagatcaaatgtgcaggcaaa
aagcttgcggaagatataattctgtcattttgttgtttagattaacttcccctgtcaaat
agctaaaactaattcatactttctcaggatccttcttgattggactactctgatcttctt
ccatatattgatatatcatattccttttgcttttaggttaacggggagatatataaccat
agagaattacgggaaaagctgaagtcccaccagtttcgaactggcagtgactgtgaagtt
attgcacatcttgtaagtactttcagaatatattcaaataaatatacatgacttctcagt
ttctgtgcgaagaatgatttcaaaataccatgactcaaatttctcatcaaaagagaaaaa
ggtggattgtagtaagagaatgagattttttgtcataatttctttcttatggtatttgaa
ctccacgatctacctatcgcagtatgaagactttggagaagacttcgttcacatgttgga
tggaatgttctcctttgtgcttcttaacacccgtgataaaagtttcattgctgctcggga
tgcaattggcattacgcccctttatatgggatggggtcttgatggtatgtaatttttcca
tttcataatttttggtatattgcccttttaataaatagcctgtgagttatgtgaagttcc
tttgttgtttccttgttttttatttgcctgatgcatctttgttgttttgcctactataat
gctggtttcatgctgttgttacaaacaaatgatgtataatagtgaacagtacatgttttc
ttgccgctgtagatcaacaatgtaccaagaacatctgtcttaagctagttcaacctaatc
ttcagttatctatcattccagcgaataacacatgtagagttaaccggacattacctaagg
ccagctatacaacattgtgaggtgtgactaaggaactctgaacactgtttggagaaatgg
aaagcaacatatccgtgtttgttttgaagtactataaggaggcagcaaccttgctggatt
tatcacttgacagcaaatgtcaatggttgaggaacattttttatggctggaggagaaaca
gagaagagaaaaaataattgattttaaaaaacagaggagaaaattttgtgaagtcgtatg
ttaatttcaatggctggaggtgcaatatttgacttggcaactagcgtgtagaagtttctg
gttggggttgagcatggaagacactgttgcttttacaactgtcattagagtcggaaacat
attgtattcttgagggagcgttgcaatggataaaaaagtcctacctactacttgatattt
gggcattcagaactggttgtctaaagctgagaaggaagaaaaggatttaaaatggtgagt
attgtaggctttcttttctgtccagggtgcctggactagttccctgaattgggatactgg
tggaggatttcctgaaattgatcatggccaaagagatatgtcaggggataagaacattga
aggatcaaaggaaggtgctgcgctaagttttgctagaaatgaacagttccactatggctt
ttgtcaaagatgggaattgaattggccccaacttctgggcatctccatccatcgtcaggt
gaacaaaagtaaggttacaatggtagaatttttcattatttcaaacaagtaaggtgaagt
cgtcagcaatcattgcctgagaaggaaccaggaacaaaatctagaagatgtctgctaaag
tgctaatagataatctgccaaatttagcacggtaaggctagtgcagatctggttcaaatt
ttaaattggtccaaaatgctttaataaatgtcggacctcgttttggtcccccaaaaggtt
gctatataaagacttgacaaagaaacaggtgaaggagaagcccaaacaattgaacaaact
acacctagcctaaagaaagtggttgatagtttggtcggaggaggaagagcgagcctgcgg
agagagcctcaaacataaaaaatgacctagtcttgtaatgagtctggccaaatcaagtaa
gctctgactttcaagagagatcaacagaaaacttcgtcagtctagataaatcaagaaaat
tatgataatgaaaaggaggctggacattctactgccgaccaaccctggttttggtgtcag
tgagacaagtggaactaaagaagaatatggatcaaaagagtttatccggttgacacctga
taaactcactacatcagttttcagttcaaaaacatttcaattcttgctgataaagacagg
caaactgtcacctaccaatcaaaggaggttgtttcaattgagttagaggtatcaaaggag
gcggcagtatatatggagcaagttttgatgaagggagttgcttctattggagaccaccag
gtactcaagtgcttcccgtagttcctaggcattttataattgatatgacagagaaatgtc
ttattgacaagaatagactaggactacagggataatagtgttgggaaggtgaggacttct
ttggttgctgtcaggaacttggaaacaaagggagaaaggggttggaatcaaaggttttga
tttcatttttgatttatgaggaaataatgggtagtggatggagtcgaggatgggaaagag
gtttaccaattgcaccattgaatatgtaattagagacactatcctcttatttgggaacac
gactagggtgagggaacgctccgacagggatgtggtttattcgggtatgttggcacgaag
cttttgtgtctaggtttggtgttggcttaagtgcaacaccttaggaagacaaacaaactt
aggacagatttgatagttgtcggaaattgcacagtgatgatattttcttcttccaaacgt
tgggaatgattgcacatgtagtaactagagttatttctcatttaagcttttatatgtaag
aatcaaataaaataattgatgaaatgcatgaaagcaactgtcttacgcctgaatgctttt
aaaaggcatgctttacaaagtttttggtacatctatctgacatgacacactaacattcac
tcttatatgtgtacatatttaactaagtccaagcctaactatctaactggagatatcaaa
gttcttagctctttaaagctttcttaactttaggaagatcctgcagtgttgtttttttat
aaccgtggtatccaggctagcttgcacacatctcgactaattccaccgggtgcttgctat
ctcccaccaacacagattacctggtaactcatgggaagaaatcacctattgttttgagtc
tgctggcatttgaagttttggactactaacctcatggttctcatcccacttcattgacca
cttggccacatccttgggtgcctgcactgtagctgtaacgaagatagtttttttttccgg
gatactcttatattgtgataatttttttgtattcgctgttaatggatgaggttatgtgac
tctccggtggtgagagtgttaatgaacatcaagctcatatgtcatttcattttctgattg
atcttgctcttcactctttacgagtttaatgtgttagtccttcccagttttttttaagag
cgagaagcgcaaaaaagcgacaagggctcgcctcgcttcaaaagcgaagcgcaaagcgaa
gcgcacactttattaaagtgaagcgcaattcttaaaaaacataatgtaaaccttgcaaag
acacaatataaaattataaataatcaaaaagttcaaattgtcaaaatcaaagctactaaa
ttactagaatcaacctcttattcttctactcttcttgttcttcttcaagattgtcaaaat
cttgaattcctctattatcttcatattgctcgtcatcttcctcctccccctcctcttcct
cttcatgatcactttcatcttcatcaattagggatagggatcgacttgtagtagccacac
tttttcccttcctaatcgagcttgaacttgaagtattcccccttaaaccataaggattct
ccccaactccactagcaaccgcaacatcaccccaagtgaaattagaatcgccttcaaata
cttcttcatcttcacaattttcggggactccggttagccactcattagcctcatcaatat
tgtccaaaagaattggatcaattagattgcggtggttgtagcgacgcctcaatgctctat
tgtattttatgaacactagattatggaggcgcttcaaggttagtcgattcctcttctttg
tatgaatctgcaaacaagatgtgtcaactaattagtaggagttgggacaattgagatata
atttactttatctcaaaacacgaaataattctttttatttctcacgtgttcaaaaacgct
ccagttcctttcacatccggatgagctacaagttaaacttagaactctgatggcgaaagt
ctgtaaattcggagtctctacaccatattggtcccaccactcaactatagaagtgaaaaa
aaaaaattcaagagttaataagtataaaaaatacattaaagttagagctataaaatatag
aagcacttggtcacctggcgatttcgtctttctttgtttaatagcaagtcggagcttaaa
aagtccctcagctgccttgtaaatagcaagctgatctactatcttttcttgcatgtcttc
atctggggtcaacttgataacaacctcatggaatcctgtccacacttctctagccaatga
attattctcatgctgatcataaaagagtgacgggttcagaataagtccagctgcatgcaa
aggtctatgaagttgctcactccatcttgcatcaatgatctgaaagacctttgcatattt
ctgctcatcagtgaatgatgcttgaatagcctccttggccctatccatagcttcatagag
gtagcccattggtggtttttgctccccatccaccaaacggagtactttaaccaaagggcc
accaattttaagagcatgaaggacattattccaaaaagaataagaaagaataatgcgtgc
aacatctttccctgcactttcctttgcaaatttactcttgctccattcctctgaagtgaa
caactttctcaaattggatttttgcaagtggatactatgtaaagtcaagaaagcagtggc
gaaccttgtcttgcccggtttcaccaaatttttttgtccggtgaatcttctcatcatatt
caataacaagggccgctgagaaatataagaatgtaccctaacgccctggccaaaaactgt
agagaagggtttttccttgaaaatgtccccgaagattaagttgatacaatgagccgcaca
tggagtccaatagacattcttgtacgctccttccaccatgccacccgctttcacattttc
actcgcattatcagtgaccacttgaacaactttgcttgggccaatcttttcaatggtgtt
ctgaaacaaggtgaacattttgatgtggtcagtggatgagtcgctagcatcaatggactc
aagaaacaaacttcccttgggagaattcaccaacacgttaataatcattttcccagttct
tgccgtccacttatccatcataatggagcagccatacttgttccacgcaactttatgttc
ctccacaattttattagtctcctccacttccttatttagataaggacctctgatttcatg
ataagtgggaggcttcattccaggtccgtattgaccaacggcctcaataaagtctccaaa
agtgtcagtatagttgacacaattgaaggggagcccagcatcatagacccatcgtgcaaa
agctctaactgcacgatccctcaaaatgtccttagcaattttttgtacatcttttccacc
gctctttccttccggcttctttgggaaatagcaatctataggacctttagtcctactcgt
acccgtcgatccatggcttgaagagattgcatctcttccccgttttgttggaagtgacaa
ttcttcaatatcatcatcaccatcatcatcaagattggtcaccaatggttgatgactcat
ttgatttttttgctccttcttcttctcaacaaaattctttatttcatccctcacctccgg
tggacatttcggacaacttgtgacgtttctatcgccaccaataagatggaatttaaagcg
ataaattccacccgttgtaatcttgttacaaaacttgcatacaatatttgtattcttctc
attaactctatcgccatatcgccaagccgggtctttatctttcgatctttgagacatttt
gaaatccctattaagatataagaaaatacataatcaaataaactgaaaatacatataata
tatataataattaattttataaaccgaaatacacattaaaaaaatcaaataaactaaaat
ataacatatataattttctaaattgaaatacatataaaattaatcaaataaactgaaaac
ataacatatatataataattaattttatatactgaaaaatgcattaagaaaatcaaataa
actgaaaaatataacatatataaataataattaattttatatactgaaataaaaatcaaa
taaactaaaatataacatatatatataataattaattttctaaaatgaaatacatataaa
attaatcaaataaactgaaaatataacatatataaataataattaatattatatactgaa
atatacattaaaaaatcaaataaactgaaaaatataacatatataaataataattaattt
tatatgctgaaatatacattaaaaaaatcaaataaactgaaaaatataacatatataaat
aataattaattttataaactaaaatacatattaaaagtaataaataagaagaataaaggg
gggaaaaaaacaaaatgggttagagataaaaggaagaagaagacgctgaagtctgaacag
tgcaaactggactgtcagagtctcagagataaacgaagaagaagaaggaagaaagaagga
aaaagaaagaaaaaaaagcagaatctgacgggaccaagaagaagaaagaaagaagaaaga
aaaaaaagagaagaagaaaattacctggactggtcagcaatgtcaatgaagttgaaccct
aggctttggtcgacctcgatcgaagaagagagaagaagaaatggccattttttgtcaatt
tagggtctgttttgtataatagaaaaacagacccaaagttttttttaaaaaaaaaagggc
ctgcgcttttttaacaaaagcgatcgcttcgtcgcttcctccgttgaagcgtgcgcttcc
ctcggtcgagtcgcctcaggcagctagaagcgacactgagtcgcgtcgcgtcgcttcgcg
ctttaagcgcgaagcgatcgcttttctaaacactggtccttccaagttctgtccatcttt
ctggtgtgcttaacccatgtatattggaccacactttgtattcatgctgattgcaacaaa
gcctgactgatcctgaactcctcctttttgccttgggtagaattatttttgtgtttttag
tgttacatatttcaatttaaaggaaaaagggagaagcttaaggtagaatcctgctccttg
acctttggatccttacacattgaagtggacagacatgtttgctttgctttgctttgatga
gttgttttagtagtcagtctaagcaactgctgatgtattctgctttattgtaagtgtcta
taatttttatctttccacaatttgctttcagctctttctgcatataagagaaaataacag
aagcttctattcgatcatgtaatattgactgcacaagtgcatatttaaacaactttggct
atgcgtatgcaagcaataataaatattactttttatgcagggtctgtatggttttcttca
gagatgaaagccttaagtgatgactgtgaacgatttgttagtttccttcccggtcatatt
tattcaagcaaaaatggtatgcccatacgaagctcatttctgagcttgcgcttcatttat
gatcagcgtacttttcctatttatttcagatttactttaggaccctgttggaaacagtct
tgaacccttattgatggtgtaattattatctcttgatcaaccaaaggagtaattacatgt
aattattacaaatttgcaacaaaatcttttaggataatcagtggagcttcatgaatcttt
taggatagttgggatgctagagtgatgtacttttgataatgtcggttaacataaatcctc
tgactttgtactgatgtactggctgagcagattttatattactttctaatccctctacaa
ataaaaaaattgtgagactgcaatataacaccatttttttaatgcatttacaatgtttaa
acttgatttagcaacgtagaacatgtcgaccttaaacagaggttctatttgtcctagata
aaggaagatcagcataatgaaacattacaggagaggagctctattgagtttttgttaatc
tagagggagcccgtaaatagtacaccaattcatctcttatatgacttctttttcattttg
gaacgttttaaaagtttgactacattttttgatctccgtacactaaactttttgatgtat
taaactaactatttccactactttaatattttcttccttaaacaactcctgttagcatct
taaactctgtgtgttcaataccctcatataaaattatacgagtattttttttatatttgc
ttatgtccttggagtttgggatggggaagctcatcatttgaggcgaaaggtaaattcttt
tgaatggacaaaaagtgaacctcggtctttggcatggagggactatttaagtcgccatca
ttaccaaagtcatcctttcaagtgtttgctttcaagaactattcatgctatgtctcaagt
atattatgtgtgcaggaggactcagaagatggtacaacccaccatggtactcagaaacca
ttccttctactccatatgatcaccttgtcttacggaaagcttttgagaaggtcattctat
tttggaagtcgcttgtttctgagttgtatataatatttaggtgccactttgttatatttt
aaacttcatcaaaagcctttggattctttgaggacacatatccccttgactgactgggag
ctgaatttccttgatgtatagtgcttattttgattgtcctctcctttgtaatactcttgc
ctataaataaaccttccttatcaaaagaaaatgataaaatgttaaaaatactttgtgata
gatgttgactatagactttaaatgatattttatctcaatctttatcaattcttttttacc
ttcacatgcaaattatactacagtatgtgaaatatgcaatagtttgtcctgcaattttca
tctttcatgtacctctctctactatccgtgtattactactgatttgatgcattaaatgct
aattttattcccaacctttactttatgcaggctgtagtcaagcgacttatgacggatgta
ccatttggtgtgcttctctcaggcggactagattcttcacttgttgctgcagtggccaac
cgctatttggctgatactgaagctgcgcgacaatggggatcacagttgcataccttttgc
gtaggcttgaaggtggtgatgctcctttttgtaaacttgctctgatttgggaattgctag
ctgtatcatcccaatttacatggaagttctagaccattattgctgtttcttttccatata
tagggattttctgaaaatagtaaaaaggaaatggaatttctctttattaaagataccgaa
tgcctttttgggaaatgatcttacagttctgaatcgaaaatttccttttgacattgtgat
taattctgatatgaatttggtccatcctggctttccttctcttgattgtagattttcctt
ctcttgttacaattatatgcctgtctttttgtcttactctctgagacaggcaatgtaact
ccagttgcaaggtttggtaatattgacttgtttatttctgttatcacatcctcctgaaaa
agtgctactagcttgtattcatgcgggaatttgaaattttcaagtgtatagctttacaaa
atgatcagaactgatcacagtcattgctctaaataaaattttctaaaaagcaggtattgt
tgtgatataatgtgacacttaaataaatttaaacctacatcctttcgaaatctgtaatag
ctccactaatatttcttgtgctaaactattccaacacctcttatcgatgccaattttgac
agaccttagtgcacttaaaagtgtttagtgttctttatgcagtaggtatgttattgtcca
aaatgacagaaataaagttagagatttgttgaaagccacttggtgtgattgagcagttcg
gagatgagtgatttatccattttatatacttgtagtgttttgttttcttttgcttttgtc
attgtaattaaggttgtcttttgtttgacattagggttctcctgatctgaaagctgccag
agaggtggcagactaccttggaacccgtcaccatgagtttcactttacagtgcaggtaac
tttccacaaagacatcctaccatgtgaccaggaggtcacgggtttgagctgtggaaacaa
ccttttgcagaaatgcagcgtaaggttgcgtacaatagacccttgtggtccggcccttcc
ccgaccccgcacatagcgggagcttagtgcaccgggctgcccatcctaagctgattttcc
ttataggagtataaccgcttaagtcaaattggctatctacttctttctattgctacatca
ggaaggaattgatgcactagatgaagtcatttatcatgttgaaacatatgatgtgaccac
tatcagagccagtacaccaatgtttctcatgtctcggaagataaagtctttgggtgtgaa
aatggttctatctggtgaaggttctgatgaaatttttggcggttatttatatttccacaa
gggacccaacaaagaggagtttcaccaagaaacttgtagaaaggtggatctcattgtcat
ttcttcagtcattcaggattattggaagtaactgtatttttactagaatacttctttgaa
ctatttctagctctgatggacgctttgatgttaatccagattaaagcacttcatctttat
gattgcttgagggccaacaaatctacttcagcttggggtgttgaagctcgtgtacctttc
ttggataaagaatttatcaatgttgcaatgaacattgatccagagtggaaaatggtaacc
tacagtgcttccgtctcatttctccaccccccacccccccaaaaaaaagaacagaaaaga
attgaaaagagagattcttttcttccttcaaggaagaagggggtaacaaaagaatagctg
caagttatgcagaaaaaatagtcttccatgtcatgggagaccagggttcaaatctgattg
ttaaaaaaaacactaggtgaactcttctaatctgtttaagccttggtggacatagttaac
tagtatatgagctagttagagctgactcgaaaccaccattattcaaaaaattaagaaagt
cttcaatgttgttccaccgtgaattcatgatgtgtcgactttcatgtgtttatctctgga
tagatgaagagtttttttggatcttgattaattctgtttggcccctttgaggttctgcac
gctatttctagactcaaacttgctgcgacctatgttaggtctagagaatatctgcctaat
aaaacaatggaagttacattcaagtattttacctccatattacaagtcagtgttagttaa
atgggtctgatatatgggatagaaaatggcttcatacccataacctcgttaaaatgttgc
tgatgtgctgatctcatctgtacatctagtataatcctaattacaaattaatgcatctct
ttgcggtgactatagtatgctgaagaacctgtcttgaaacattgcagatcagacctgatc
tcggaagaatagaaaagtgggttctacgcaatgcttttgacgatgatcagaatccttatc
tgccaaaggtttgtttgttttcttgtatcaggtccaataatcataatctttgtttattca
tttgcataaaaggcagtagactcccaatacattgtgtacgcttatgcacaaatgtatgag
atgtgtattatatgtgtcacatataagggtattatatgtacacactcctcccaccttcca
catatattatgtgtgtgtgtgtgtgtgtataatgaaatgatgtttgaaaagacattatga
tcaagtagttgatcctcatctgctttctgtttttagcatatcttgtataggcagaaggaa
cagttcagtgatggagttggctacagttggattgatggcttgaaggatcacgcaagcagt
ctggtgagctctttagttgttttctcttccctcctcttccattctttatcccttcctttt
gtggttttgttgaacacatctcatatacttggtccaggtttctgattctatgttagcgaa
tgcaagttttgtttacccgcataacacacccacgacaaaggaaggatactattatagaac
tatttttgagcgatatttccccaaggtgggtcttacatagtcttcttttccttgttattc
atgttcagtttatgatattttgtcccaaacaatgttttctttttcttcctttttctgtta
acaagatcgcttctatcttcgtatttgttgataaagttaatcacattgttaactattatt
tggagcagaatgctgcgagggaaacagttccaggtggtccaagtgtggcatgcagcactg
caaaagcagtagaatgggacgcagcttggtccaagaatctagatccatctggtcgagctg
cactcggtgttcatgcagctgcttatgaggatgcatcagaggttaaaaccacagtcgcga
cagatactgctcagaaacttgacgttgataaagctgcagtagctgtttga
SEQ ID NO:12 - NtASN3-T, предсказанная аминокислотная последовательность
MCGILAVFGCIDNSQAKRSRIIELSRRLRHRGPDWSGLHSHDDCYLAHQRLAIVDPTSGDQPLYNEDKTIVVAVNGEIYNHRDLREKLKSHQFRTGSDCEVIAHLYEDYGEDFVHMLDGMFSFVLLDTRDKSFIAARDAIGITPLYMGWGLDGSVWFSSEMKALSDDCERFVSFLPGHIYSSKNGGLRRWYNPPWYSETIPSTPYDHLVLRKAFEKAVVKRLMTDVPFGVLLSGGLDSSLVAAVANRYLADTEAARQWGSQLHTFCVGLKGSPDLKAAREVADYLGTRHHEFHFTVQEGIDALDEVIYHVETYDVTTIRASTPMFLMSRKIKSLGVKMVLSGEGSDEIFGGYLYFHKAPNKEEFHQETCRKIKALHLYDCLRANKSTSAWGVEARVPFLDKEFVNVAMNMDPESKMIRPDLGRIEKWVLRNAFDDDQNPYLPKHILYRQKEQFSDGVGYSWIDGLKDHASRLVSDSMLANASFVYPHNTPTTKEGYYYRTIFERYFPKNAARETVPGGPSVACSTAKAVEWDAAWSKNLDPSGRAALGVHAAAYEDASEVKTTVATDTPQKLEVDKAAVAV
SEQ ID NO: 13 - последовательность ДНК праймера NtASN1, ASN1-f
tcacatatattccctcataacacac
SEQ ID NO: 14 - последовательность ДНК праймера NtASN1, ASN1-r
aagaagcatcccactctacagctt
SEQ ID NO: 15- последовательность ДНК праймера NtASN5, ASN5-f
atatcttccctcacaacactccaact
SEQ ID NO: 16 - последовательность ДНК праймера NtASN5, ASN5-r
Ctaccggaaggatcaaggttgt
SEQ ID NO: 17 - кодирующая последовательность ДНК из NtASN1-S, использованная для сайленсинга обеих копий NtASN1-S и NtASN1-T
tcacatatattccctcataacacacccattacaaaggaagcatactactataggatgattttcgagcgctttttcccacagaattcagctgggctaaccgttcctggaggagcaagtgtggcgtgtagcacagctaaagctgtagagtgggatgcttcttggtcaaagaaccttgatccttcaggcagggctgctattggtgtacataactcggcttatgagaatcatgtacctgctatggctaatgggaatttgaccaaaaaaatcattggtcgtgtgccttctatggtagaagttggtgctgctcccgagctcacaataaagagttag
SEQ ID NO: 18 - кодирующая последовательность ДНК из NtASN5-T, использованная для сайленсинга обеих копий NtASN5-S и NtASN5-T
gaccagattggagtgggatatatcaacatggtgatttttacttagcacatcaacgtttagcaattatcgatcctacttctggtgatcagcctctgtttaatcaagataagactattgttgttacagtcaatggagaaatttacaatcatgagaaacttcgtaatcttatgcctaatcacaagttcagaaccggaagtgattgtgatgttattgcacatctttatgaagaatatggagaaaattttgtggacatgttggatggggtgttctcttttgtattgttggatacgcgcgataacagctttcttgctgctcgtgatgcgattggaattactcccctctatattggttggggacttgatgg
SEQ ID NO: 19 - последовательность ДНК стоп-мутанта ASN1-S: ASN1-S_W156*
atgtgcgggatcttggctgttttgggttgttctgatgattctcaggccaaaagggttcgtgttctcgagctctctcgcaggtaggttgaagcatcgtggaccagattggagtgggctgtatcaacatggggactgttacttggcacatcagcgtctagctattgttgatcctgcttccggtgatcaacctctgtttaacgaagataagacgattgttgttacggtaggtaaatggagagatctacaatcacgagcaacttcgtaagcaaatgcctaatcataagttccggactggcagtgactgtgatgtcattgcacacctagtagtatgaagaacatggagaagattttgtggacatgctggatgggatcttcgcttttgtgttattggatactcgagataacagctttcttgttgctcgtgatgccattggaattacttccctttatattggttggggacttgatggtagggtctgtatgaatatcatctgagcttaagggcttgaatgatgactgcgaacattttgaagttttcccaccagggcacttgtactctagcaagaatggcggctttaggaggtggtacaatcctccttggttctctgaggccattccttccactcgttatgatcccttagttctcaggcgtgcctttgaaaatgtaggctgttatcaaaaggttgatgactgatgtcccctttggtgttctcctctccgggggactcgattcatccttggttgcttcgattactgctcgctacttggctggtacaaaggctgccaagcagtggggagcacagcttcattccttctgtgttggccttgaggtagggctcaccggatctcaaggctgcaagagaagttgctgactacttgggaaccgttcaccacgagtttcacttcaccgttcaggtaggatggaattgatgcaattgaagatgttatttaccatattgagacatacgatgtaacgacaatcagagcaagcactcctatgttccttatgtcgcgtaagattaagtcactaggagtgaagatggtcatatctggggaaggatctgatgaagtgtttggtggctacttgtactttcacaaggctcccaacaaggaagagttccacaaggaaacatgtcgcaaggtagattaaagcgcttcaccaatatgactgcttaagagcaaataagtcaacatctgcatggggtttagaagctagagtccctttcctagataaggagttcatcaatgttgccatgagtattgatccagagtggaagttgagattaaaccagagcaaaggaggattgaaaagtgggctctaaggagggcctttgatgatgaggagcatccttatctcccaaaggtagcacatcctgtataggcaaaaagaacaattcagtgatggcgtgggcatagttggatagatggactcaaagcacatgctgaacaacatgtaggtgaccaataggatgatgtttaatgcttcacatatattccctcataacacacccattacaaaggaagcatactactataggatgattttcgagcgctttttcccacaggtagaattcagctgggctaaccgttcctggaggagcaagtgtggcgtgtagcacagctaaagctgtagagtgggatgcttcttggtcaaagaaccttgatccttcaggcagggctgctattggtgtacataactcggcttatgagaatcatgtacctgctatggctaatgggaatttgaccaaaaaaatcattggtcgtgtgccttctatggtagaagttggtgctgctcccgagctcacaataaagagt
SEQ ID NO: 20 - аминокислотная последовательность стоп-мутанта ASN1-S: ASN1-S_W156*
MetCysGlyIleLeuAlaValLeuGlyCysSerAspAspSerGlnAlaLysArgValArgValLeuGluLeuSerArgArgLeuLysHisArgGlyProAspTrpSerGlyLeuTyrGlnHisGlyAspCysTyrLeuAlaHisGlnArgLeuAlaIleValAspProAlaSerGlyAspGlnProLeuPheAsnGluAspLysThrIleValValThrValAsnGlyGluIleTyrAsnHisGluGlnLeuArgLysGlnMetProAsnHisLysPheArgThrGlySerAspCysAspValIleAlaHisLeuTyrGluGluHisGlyGluAspPheValAspMetLeuAspGlyIlePheAlaPheValLeuLeuAspThrArgAspAsnSerPheLeuValAlaArgAspAlaIleGlyIleThrSerLeuTyrIleGlyTrpGlyLeuAspGlySerValSTOP
SEQ ID NO: 21 - последовательность ДНК стоп-мутанта ASN1-T: ASN1-T_W156*
atgtgcgggatcttggctgttttgggttgttctgatgattctcaggccaaaagggttcgtgttctcgagctctctcgcaggtaggttgaagcatcgtggaccagattggagtgggctgtatcaacatggggactgttacttggcacatcagcgtctagctattgttgatcctgcttccggtgatcaacctctgtttaacgaagataagacgattgttgttacggtaggtaaatggagagatctacaatcacgagcaacttcgcaagcaaatgcctgatcataagttccggactggaagtgactgtgatgtcattgcacacctagtagtatgaagaacatggagaagattttgtggacatgctggatgggatcttcgcttttgtgttactggatactcgagataacagctttcttgttgctcgtgatgccattggaattacttccctttatattggttggggacttgatggtagggtctgtatgaatatcatctgagcttaagggcttgaatgatgactgcgaacattttgaagttttcccaccaggacacttgtactctagcaagaatggcggctttaggaggtggtacaatcctctttggttctctgaggctattccttccactccttatgatcccttagttctcaggcgcgcctttgaaaatgtaggctgttatcaaaaggttgatgactgatgtcccttttggtgttctgctctccgggggactcgattcatccttggttgcttcgattactgcccgctacttggctggcacaaaggctgccaagcagtggggagcacagcttcattccttctgtgttggccttgaggtagggatcaccggatctcaaggctgcaagagaagttgctgactacttgggaaccgttcaccacgagtttcacttcaccgttcaggtaggatggaattgatgcaattgaagatgttatttaccatattgagacatacgatgtaacgacaatcagagcaagcactcctatgttccttatgtcgcgtaagattaagtcactaggagtgaagatggttatatctggggaaggctctgatgaagtgtttggtggctacttgtactttcacaaggctcccaacaaggaagagttccacaaggaaacatgtcgcaaggtagattaaagcacttcaccaatatgactgtttaagagcaaataagtcaacatctgcatggggcttagaagctagagtgcctttcctagataaggagttcatcaatgttgccatgagtattgatccagagtggaagttggtagattaaaccagagcaaaggaggattgagaagtgggctctaaggagggcctttgatgatgaggagcatccctatctcccaaaggtagcacatcctatacaggcagaaagaacaattcagtgatggcgtaggctatagttggatagatggactcaaagcacatgctgaacaacatgtaggtgaccaataggatgatgcttaatgcttcacatatattccctcataacacaccgattacaaaggaagcatactattataggatgattttcgagcgctttttcccacaggtagaattcagctgggctaaccgttcctggaggagcgagtgtggcgtgtagcacagctaaagctgtagagtgggatgcttcttggtcaaagaaccttgatccttcaggaagggctgctattggtgtacataactcagcttatgagaatcatgaacctgctatggctaatgggaatttggccacaaaaatcattggccgtgcgccgtctatggtagaagttggtgctgctcatgagctcacaataaggagt
SEQ ID NO: 22 - аминокислотная последовательность стоп-мутанта ASN1-T: ASN1-T_W156*
MetCysGlyIleLeuAlaValLeuGlyCysSerAspAspSerGlnAlaLysArgValArgValLeuGluLeuSerArgArgLeuLysHisArgGlyProAspTrpSerGlyLeuTyrGlnHisGlyAspCysTyrLeuAlaHisGlnArgLeu
AlaIleValAspProAlaSerGlyAspGlnProLeuPheAsnGluAspLysThrIleValValThrValAsnGlyGluIleTyrAsnHisGluGlnLeuArgLysGlnMetProAspHisLysPheArgThrGlySerAspCysAspValIleAlaHisLeuTyrGluGluHisGlyGluAspPheValAspMetLeuAspGlyIlePheAlaPheValLeuLeuAspThrArgAspAsnSerPheLeuValAlaArgAspAlaIleGlyIleThrSerLeuTyrIleGlyTrpGlyLeuAspGlySerValSTOP
SEQ ID NO: 23 - последовательность ДНК стоп-мутанта ASN5-S: ASN5-S_Q66*
atgtgtggaatcttggctttgttgggttgttcagatgattctcaggccaaaagggttcgagttcttgagctctctcgcaggcaggttgaagcatcgtggaccagattggagtgggatatatcaacatggtgatttttacttagcacatcaacgtttagcaattatcgatcctgcttctggtgatcagcctctgtttaattaagataagacgattgttgttacagtaggtcaatggagagatttacaatcatgagaaacttcgtaatcttatgcctaatcacaagttcagaactggaagtgattgtgatgttattgcacatcttgtagtatgaagaatatggagaaaattttgtggacatgttggatggggtgttctcttttgtattgttggatacgcgcgataacagctttcttgctgctcgtgatgcaattggaattactcccctatatattggttggggacttgatggtaggctctgtgtggatttcatctgagctaaaaggattaaatggtgactgtgaacattttgaagttttccctcccggtcacttgtactcgagcaagaatggcgggtttaggagatggtacaatcctcaatggttctctgaggctattccatcaaatccttacgaccccttagttttgagacgtgccttcgaaaatgtaggctgttatcaaacgattgatgaccgatgtcccctttggtgttctgctctccgggggacttgattcgtctttggttgcttctgtcactgctcgctacttggctggaacaaaagctgctaagcaatggggagcacagcttcattccttctgtgttggtctcgaggtagggctcaccagatctcaaggctgcaagagaagttgctgactatttgggaaccgttcaccacgagttcaccttcacagttcaggtaggatggaattgatgctattgaagatgttatttaccatatcgagacatacgatgtaacaacgatcagagcaagcactcctatgttccttatgtcgcgtaagattaaatcactgggagttaagatggtcatatcaggggaaggctcagatgaactgtttggcggctatttgtacttccacaaggctccgaacaaggaagaattccatgtggagacatgtcacaaggtagataaaagcgcttcaccaatacgactgtttgagagcaaataaggcaacatcagcatggggcttagaagctagagtaccatttctggataaagagttcatcaacgttgctatgagtatcgatcctgaatggaagatggtagattaaacacgatcatggtaggatcgagaagtgggttcttaggaaggcttttgatgatgaggagcaaccctatctcccaaaggtagcatattctgtaccggcagaaagaacaattcagtgatggcgtaggctatagttggatcgatggactcaaagcacatgctgaacaacatgtaggtgactgataggatgatgcttaatgctgcacatatcttccctcacaacactccaactacaaaggaagcatactattacaggatgattttcgagaggttcttcccacaggtagaattcagcaaggctaactgttcctggaggaccgagtatagcttgcagcacagctaaagctattgagtgggatgcttcgtggtcgaacaaccttgatccttccggtagggctgcaatcggtgtacataactcggcttatgacgatcatctccccgatgttggtaatgggaatttggacacaacgatcatcgataatgtgccgaggatggtaggagtgggtgctgctgcagagctcacaataaggagc
SEQ ID NO: 24 - аминокислотная последовательность стоп-мутанта ASN5-S: ASN5-S_Q66*
MetCysGlyIleLeuAlaLeuLeuGlyCysSerAspAspSerGlnAlaLysArgValArgValLeuGluLeuSerArgArgLeuLysHisArgGlyProAspTrpSerGlyIleTyrGlnHisGlyAspPheTyrLeuAlaHisGlnArgLeuAlaIleIleAspProAlaSerGlyAspGlnProLeuPheAsnSTOP
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Филип Моррис Продактс С.А.
<120> СНИЖЕНИЕ СОДЕРЖАНИЯ АСПАРАГИНА В РАСТЕНИЯХ
<130> P10251EP
<140> 15184528.6
<141> 2015-09-09
<160> 24
<170> PatentIn version 3.5
<210> 1
<211> 3646
<212> ДНК
<213> Nicotiana tabacum
<400> 1
atgtgcggga tcttggctgt tttgggttgt tctgatgatt ctcaggccaa aagggttcgt 60
gttctcgagc tctctcgcag gtaaattata tttcccttct tttctgtatt gttaatgtta 120
ttgtgttgcg ttattatatt gattcttttt atgagaaagt agtttcctga acgtttgcat 180
tttagaataa aagaagacta aagcttaaga tagattatat atttagatat tcttgacgtg 240
gacctatatc ttagattcac aatataattt tctcggacca actgttttag catgacttgt 300
aaatgatgtt gatatgaaaa gctgtgattt aattagtagg agtaattcat ttccatgtta 360
caattactta tctttctcct ccttgtgttc ttgccactgc tagaggcaga tctataattc 420
ctacttagag tattgaacac attgtacttt tgaaattgta agtgtgcagt ttaatatttg 480
ttaaaatttt gaaggttttt ttaattacat gtatatcgtg acgttatata gcctctctac 540
ccgcataagg taggggtaat gtctgcgtac aaataggctg atttcactct gtgttcagtt 600
gaatcaatat atgcggcatc caccactgtt tgcaaatgca tgggaatgga aaagtcatgc 660
ctcatcactg tcacacatga tttttattta tttatttatt atatatgatt ctttaaatct 720
attagtgggt actgaatctt gctaactgat cgattgatga aacaggttga agcatcgtgg 780
accagattgg agtgggctgt atcaacatgg ggactgttac ttggcacatc agcgtctagc 840
tattgttgat cctgcttccg gtgatcaacc tctgtttaac gaagataaga cgattgttgt 900
tacggtgggt gcttttaatt tcatgtttac tctcatttct tgctcagttt actgatgaaa 960
acgaaactaa tgctgataac tatcaaccaa tcaaccacat acaagtaggg gtcgactata 1020
tgattcatta tatcctttcc tctattcaaa tagttgatga ttaatttcat gccaaaaatt 1080
tcaggtaaat ggagagatct acaatcacga gcaacttcgt aagcaaatgc ctaatcataa 1140
gttccggact ggcagtgact gtgatgtcat tgcacaccta gtgagtactc atcttccaac 1200
ttaggaaaca tgtatgtata tgtaaaatta ttccctgatt atattatgta cattaatcag 1260
tatgaagaac atggagaaga ttttgtggac atgctggatg ggatcttcgc ttttgtgtta 1320
ttggatactc gagataacag ctttcttgtt gctcgtgatg ccattggaat tacttccctt 1380
tatattggtt ggggacttga tggtaagatt ccatatgatt ttctcactta gattttgtta 1440
ttgcaaatga atatgttgaa tccttatatg tactaattgt tgtaattctg ttccttctca 1500
gggtctgtat ggatatcatc tgagcttaag ggcttgaatg atgactgcga acattttgaa 1560
gttttcccac cagggcactt gtactctagc aagaatggcg gctttaggag gtggtacaat 1620
cctccttggt tctctgaggc cattccttcc actcgttatg atcccttagt tctcaggcgt 1680
gcctttgaaa atgtgagtta atattgttgt tgttgaaatg gatccaacat gttggtttta 1740
aggctatata tcattttgca ggctgttatc aaaaggttga tgactgatgt cccctttggt 1800
gttctcctct ccgggggact cgattcatcc ttggttgctt cgattactgc tcgctacttg 1860
gctggtacaa aggctgccaa gcagtgggga gcacagcttc attccttctg tgttggcctt 1920
gaggtcagat aatcttgaga cttgtgtgat tacaacatta ataacgacaa cacctcattc 1980
acaagctagt aaggtcagct atatgaaatc tgataccgat ttgcttgctt tcaccgaaca 2040
gggctcaccg gatctcaagg ctgcaagaga agttgctgac tacttgggaa ccgttcacca 2100
cgagtttcac ttcaccgttc aggttttgat ggagcaactc tattttcact tgatctatca 2160
tcctcttcgt ttctcctttg tttacctgat ggcaaatgac atgtgttgta tttgcaggat 2220
ggaattgatg caattgaaga tgttatttac catattgaga catacgatgt aacgacaatc 2280
agagcaagca ctcctatgtt ccttatgtcg cgtaagatta agtcactagg agtgaagatg 2340
gtcatatctg gggaaggatc tgatgaagtg tttggtggct acttgtactt tcacaaggct 2400
cccaacaagg aagagttcca caaggaaaca tgtcgcaagg tataagaaag tgcttccagt 2460
tcctagttta aataggacaa ggagttgaga aaatgttgat tttttcccta tgcagattaa 2520
agcgcttcac caatatgact gcttaagagc aaataagtca acatctgcat ggggtttaga 2580
agctagagtc cctttcctag ataaggagtt catcaatgtt gccatgagta ttgatccaga 2640
gtggaagttg gtaagtagtt gacccccaat tgttgatcga ataaacaatc aactgtccct 2700
aaaactatgt atttattcat ttcagattaa accagagcaa aggaggattg aaaagtgggc 2760
tctaaggagg gcctttgatg atgaggagca tccttatctc ccaaaggtgt actaaatcgc 2820
cattttcaaa ctgatttgca aatgattttg aatgttatat tattaatcaa actaacaatt 2880
tactcattac attccttgca gcacatcctg tataggcaaa aagaacaatt cagtgatggc 2940
gtgggctata gttggataga tggactcaaa gcacatgctg aacaacatgt gcgtttcaat 3000
gactaatcaa cgactttcta ctgctcttat ttatcttcca atgcttgttt gttttaaagc 3060
tgaccgagga aacaaatttt ttcctttaat aggtgaccaa taggatgatg tttaatgctt 3120
cacatatatt ccctcataac acacccatta caaaggaagc atactactat aggatgattt 3180
tcgagcgctt tttcccacag gtaatttgtt cagataatag tgagaattta tgataaatgg 3240
actaatttac gtttgttgtc gattttgcaa ccttccaaca gtgtgagcaa gctcacataa 3300
tcggaatttt ttgttcaaat aataatgcat tggaaatcat aatgctaaat tgagatttac 3360
ttttaaccat gttcctctac catttttggt tctgcagaat tcagctgggc taaccgttcc 3420
tggaggagca agtgtggcgt gtagcacagc taaagctgta gagtgggatg cttcttggtc 3480
aaagaacctt gatccttcag gcagggctgc tattggtgta cataactcgg cttatgagaa 3540
tcatgtacct gctatggcta atgggaattt gaccaaaaaa atcattggtc gtgtgccttc 3600
tatggtagaa gttggtgctg ctcccgagct cacaataaag agttag 3646
<210> 2
<211> 590
<212> PRT
<213> Nicotiana tabacum
<400> 2
Met Cys Gly Ile Leu Ala Val Leu Gly Cys Ser Asp Asp Ser Gln Ala
1 5 10 15
Lys Arg Val Arg Val Leu Glu Leu Ser Arg Arg Leu Lys His Arg Gly
20 25 30
Pro Asp Trp Ser Gly Leu Tyr Gln His Gly Asp Cys Tyr Leu Ala His
35 40 45
Gln Arg Leu Ala Ile Val Asp Pro Ala Ser Gly Asp Gln Pro Leu Phe
50 55 60
Asn Glu Asp Lys Thr Ile Val Val Thr Val Asn Gly Glu Ile Tyr Asn
65 70 75 80
His Glu Gln Leu Arg Lys Gln Met Pro Asn His Lys Phe Arg Thr Gly
85 90 95
Ser Asp Cys Asp Val Ile Ala His Leu Tyr Glu Glu His Gly Glu Asp
100 105 110
Phe Val Asp Met Leu Asp Gly Ile Phe Ala Phe Val Leu Leu Asp Thr
115 120 125
Arg Asp Asn Ser Phe Leu Val Ala Arg Asp Ala Ile Gly Ile Thr Ser
130 135 140
Leu Tyr Ile Gly Trp Gly Leu Asp Gly Ser Val Trp Ile Ser Ser Glu
145 150 155 160
Leu Lys Gly Leu Asn Asp Asp Cys Glu His Phe Glu Val Phe Pro Pro
165 170 175
Gly His Leu Tyr Ser Ser Lys Asn Gly Gly Phe Arg Arg Trp Tyr Asn
180 185 190
Pro Pro Trp Phe Ser Glu Ala Ile Pro Ser Thr Arg Tyr Asp Pro Leu
195 200 205
Val Leu Arg Arg Ala Phe Glu Asn Ala Val Ile Lys Arg Leu Met Thr
210 215 220
Asp Val Pro Phe Gly Val Leu Leu Ser Gly Gly Leu Asp Ser Ser Leu
225 230 235 240
Val Ala Ser Ile Thr Ala Arg Tyr Leu Ala Gly Thr Lys Ala Ala Lys
245 250 255
Gln Trp Gly Ala Gln Leu His Ser Phe Cys Val Gly Leu Glu Gly Ser
260 265 270
Pro Asp Leu Lys Ala Ala Arg Glu Val Ala Asp Tyr Leu Gly Thr Val
275 280 285
His His Glu Phe His Phe Thr Val Gln Asp Gly Ile Asp Ala Ile Glu
290 295 300
Asp Val Ile Tyr His Ile Glu Thr Tyr Asp Val Thr Thr Ile Arg Ala
305 310 315 320
Ser Thr Pro Met Phe Leu Met Ser Arg Lys Ile Lys Ser Leu Gly Val
325 330 335
Lys Met Val Ile Ser Gly Glu Gly Ser Asp Glu Val Phe Gly Gly Tyr
340 345 350
Leu Tyr Phe His Lys Ala Pro Asn Lys Glu Glu Phe His Lys Glu Thr
355 360 365
Cys Arg Lys Ile Lys Ala Leu His Gln Tyr Asp Cys Leu Arg Ala Asn
370 375 380
Lys Ser Thr Ser Ala Trp Gly Leu Glu Ala Arg Val Pro Phe Leu Asp
385 390 395 400
Lys Glu Phe Ile Asn Val Ala Met Ser Ile Asp Pro Glu Trp Lys Leu
405 410 415
Ile Lys Pro Glu Gln Arg Arg Ile Glu Lys Trp Ala Leu Arg Arg Ala
420 425 430
Phe Asp Asp Glu Glu His Pro Tyr Leu Pro Lys His Ile Leu Tyr Arg
435 440 445
Gln Lys Glu Gln Phe Ser Asp Gly Val Gly Tyr Ser Trp Ile Asp Gly
450 455 460
Leu Lys Ala His Ala Glu Gln His Val Thr Asn Arg Met Met Phe Asn
465 470 475 480
Ala Ser His Ile Phe Pro His Asn Thr Pro Ile Thr Lys Glu Ala Tyr
485 490 495
Tyr Tyr Arg Met Ile Phe Glu Arg Phe Phe Pro Gln Asn Ser Ala Gly
500 505 510
Leu Thr Val Pro Gly Gly Ala Ser Val Ala Cys Ser Thr Ala Lys Ala
515 520 525
Val Glu Trp Asp Ala Ser Trp Ser Lys Asn Leu Asp Pro Ser Gly Arg
530 535 540
Ala Ala Ile Gly Val His Asn Ser Ala Tyr Glu Asn His Val Pro Ala
545 550 555 560
Met Ala Asn Gly Asn Leu Thr Lys Lys Ile Ile Gly Arg Val Pro Ser
565 570 575
Met Val Glu Val Gly Ala Ala Pro Glu Leu Thr Ile Lys Ser
580 585 590
<210> 3
<211> 4564
<212> ДНК
<213> Nicotiana tabacum
<220>
<221> misc_feature
<222> (3137)..(3137)
<223> n представляет собой a, c, g или t
<400> 3
atgtgcggga tcttggctgt tttgggttgt tctgatgatt ctcaggccaa aagggttcgt 60
gttctcgagc tctctcgcag gtaaattata ttcccttctt ttctctattg ttaatgttat 120
tgtgttgcat gcgttattaa ttctttttat gagaaagtag tttcctgagc gtttggagct 180
gctactgatt gcttgcattt tagaataaaa cttaagatgg attatatatt tagatattct 240
tgacgtggac ctatatctta gattcacaat tttctcggac caactgtttt agcatgactt 300
gtatatgatg ttgatatgaa aagctgtggt ttagtaattc atttccatgt tacaattact 360
aatctttctc ctatctgtgt tcttgccact gctagaggca gatctataat tcgtaattag 420
agtattgaac acattgtact tttcaaaatt ataagtgcgc aatttaatat ttgttaaaat 480
tttgaaggct tttttaatta catgtatatc gtgtctcgta tagcctctct acccatgtaa 540
ggtctgcgta cacattaccc gccccatgcc ctcccgtgaa attttactgg ggtttgctgt 600
tgttgttggt atcgtgtgtc gcatatactt tgttctgttg aatcaatata tgcgtcatct 660
gccactgttt gcaaatgcat ggaatggaaa agtcatgcct catcactgtc acacaggatt 720
tttatttatt tatttattta ttatatatga ttctttttct tgatgtgaaa tctattagtg 780
ggtactgaat ctggctaact gactaatgaa acaggttgaa gcatcgtgga ccagattgga 840
gtgggctgta tcaacatggg gactgttact tggcacatca gcgtctagct attgttgatc 900
ctgcttccgg tgatcaacct ctgtttaacg aagataagac gattgttgtt acggtgggtg 960
cttttaattt catgtttact ctcatttctt gctcagttta ctgatgaaca tgaaactaat 1020
gactatcaat cagccaatca accacgctta attccataca agtaggggtc gactatatga 1080
ttcattatat cctttcctct attccggttc ctttttattc aaatagttga tgattaattt 1140
catgccaaaa atttcaggta aatggagaga tctacaatca cgagcaactt cgcaagcaaa 1200
tgcctgatca taagttccgg actggaagtg actgtgatgt cattgcacac ctagtgagta 1260
atctagatta ctcatcttcc aacttaggaa acatgtatgt attcccctga ttatttcatc 1320
atctttatgt acattaatca gtatgaagaa catggagaag attttgtgga catgctggat 1380
gggatcttcg cttttgtgtt actggatact cgagataaca gctttcttgt tgctcgtgat 1440
gccattggaa ttacttccct ttatattggt tggggacttg atggtaagat tccatattat 1500
tttctcactt agattttgtt attgcaaatg aatatgttga atctttatat gtagtaactg 1560
ttgtaatttg gttccttctc agggtctgta tggatatcat ctgagcttaa gggcttgaat 1620
gatgactgcg aacattttga agttttccca ccaggacact tgtactctag caagaatggc 1680
ggctttagga ggtggtacaa tcctctttgg ttctctgagg ctattccttc cactccttat 1740
gatcccttag ttctcaggcg cgcctttgaa aatgtgagtt attattgttg ttgttgaaat 1800
ggatccaaca tgttggttct aaggctggat atcattgtgc aggctgttat caaaaggttg 1860
atgactgatg tcccttttgg tgttctgctc tccgggggac tcgattcatc cttggttgct 1920
tcgattactg cccgctactt ggctggcaca aaggctgcca agcagtgggg agcacagctt 1980
cattccttct gtgttggcct tgaggtcaga taatcttgag atttctgtga ttacaacatt 2040
aataacgacc acacctcatt cacaagctag taaggtcagc tatatgaatc tgataccgat 2100
ttgcttgctt tcaaacgaac agggatcacc ggatctcaag gctgcaagag aagttgctga 2160
ctacttggga accgttcacc acgagtttca cttcaccgtt caggtattga tggagcaact 2220
ctattttgac ttgatctatc atcctctgcg ttcccttttg tttacctgat ggcaaatgac 2280
atgtgtcgta tttgcaggat ggaattgatg caattgaaga tgttatttac catattgaga 2340
catacgatgt aacgacaatc agagcaagca ctcctatgtt ccttatgtcg cgtaagatta 2400
agtcactagg agtgaagatg gttatatctg gggaaggctc tgatgaagtg tttggtggct 2460
acttgtactt tcacaaggct cccaacaagg aagagttcca caaggaaaca tgtcgcaagg 2520
tataaaaaag tgtttccagt tccttgttta aaaccttatt taaataggat aagcaacaac 2580
aacaacaaca acccagtata atcccactag tggggtctga ggagggtatt gtgtatgcag 2640
accttacccc taccctgggg tagagaggct gtttcagata gaaactcgac tccctccctc 2700
caaaaacctt atttaaatag gacaaggagt tgagaaaatg ttgatctttt tcctccatct 2760
ctatgcagat taaagcactt caccaatatg actgtttaag agcaaataag tcaacatctg 2820
catggggctt agaagctaga gtgcctttcc tagataagga gttcatcaat gttgccatga 2880
gtattgatcc agagtggaag ttggtaagta gttgaacccc aattgttgat cgaataaacg 2940
atcaactgtc cctaaaacct tgtattctat ccatttcaga ttaaaccaga gcaaaggagg 3000
attgagaagt gggctctaag gagggccttt gatgatgagg agcatcccta tctcccaaag 3060
gtggtactaa atcgcccttt tcaaactgat ttgcaagtga tttcgaatgt tatattgtta 3120
accaaactaa caatttnaag gctctgatga agtgtttggt ggctacttgt actttcacaa 3180
ggctcccaac aaagaagagt tccacaagga aacatgtcgc aaggtataaa aaagtgtttc 3240
cagttccttg tttaaaacct tatttaaata ggataagcaa caacaacaac aacaacccag 3300
tataatccca ctagtggggt ctgaggaggg tattgtgtat gcagacctta cccctaccct 3360
ggggtagaga ggctgtttca gatagaaact cgactccctc cctccaaaaa ccttatttaa 3420
ataggacaag gagttgagaa aatgttgatc tttttcctcc atctctatgc agattaaagc 3480
acttcaccaa tatgactgtt taagagcaaa taagtcaaca tctgcatggg gcttagaagc 3540
tagagtgcct ttcctagata aggagttcat caatgttgcc atgagtattg atccagagtg 3600
gaagttggta agtagttgaa ccccaattgt tgatcgaata aacgatcaac tgtccctaaa 3660
accttgtatt ctatccattt cagattaaac cagagcaaag gaggattgag aagtgggctc 3720
taaggagggc ctttgatgat gaggagcatc cctatctccc aaaggtggta ctaaatcgcc 3780
cttttcaaac tgatttgcaa gtgatttcga atgttatatt gttaaccaaa ctaacaattt 3840
actcattaca tttcctgcag cacatcctat acaggcagaa agaacaattc agtgatggcg 3900
taggctatag ttggatagat ggactcaaag cacatgctga acaacatgtg cgtttcaatg 3960
actaatcaac aactttctaa tgttcttatt tatcttccaa tgtttgtttg ttttaaagct 4020
gacttggaat ttttttgttt cctttaatag gtgaccaata ggatgatgct taatgcttca 4080
catatattcc ctcataacac accgattaca aaggaagcat actattatag gatgattttc 4140
gagcgctttt tcccacaggt aatttgttca tgtattgtga ggattcatga taaattagct 4200
aacttatgct ggttttcaac cttgcaacct tccgataagt cacattgttg gagtttgttg 4260
ttcaaatgcg agatttactt ttaaccatgt tcctctggta tttttggttc tgcagaattc 4320
agctgggcta accgttcctg gaggagcgag tgtggcgtgt agcacagcta aagctgtaga 4380
gtgggatgct tcttggtcaa agaaccttga tccttcagga agggctgcta ttggtgtaca 4440
taactcagct tatgagaatc atgaacctgc tatggctaat gggaatttgg ccacaaaaat 4500
cattggccgt gcgccgtcta tggtagaagt tggtgctgct catgagctca caataaggag 4560
ttag 4564
<210> 4
<211> 590
<212> PRT
<213> Nicotiana tabacum
<400> 4
Met Cys Gly Ile Leu Ala Val Leu Gly Cys Ser Asp Asp Ser Gln Ala
1 5 10 15
Lys Arg Val Arg Val Leu Glu Leu Ser Arg Arg Leu Lys His Arg Gly
20 25 30
Pro Asp Trp Ser Gly Leu Tyr Gln His Gly Asp Cys Tyr Leu Ala His
35 40 45
Gln Arg Leu Ala Ile Val Asp Pro Ala Ser Gly Asp Gln Pro Leu Phe
50 55 60
Asn Glu Asp Lys Thr Ile Val Val Thr Val Asn Gly Glu Ile Tyr Asn
65 70 75 80
His Glu Gln Leu Arg Lys Gln Met Pro Asp His Lys Phe Arg Thr Gly
85 90 95
Ser Asp Cys Asp Val Ile Ala His Leu Tyr Glu Glu His Gly Glu Asp
100 105 110
Phe Val Asp Met Leu Asp Gly Ile Phe Ala Phe Val Leu Leu Asp Thr
115 120 125
Arg Asp Asn Ser Phe Leu Val Ala Arg Asp Ala Ile Gly Ile Thr Ser
130 135 140
Leu Tyr Ile Gly Trp Gly Leu Asp Gly Ser Val Trp Ile Ser Ser Glu
145 150 155 160
Leu Lys Gly Leu Asn Asp Asp Cys Glu His Phe Glu Val Phe Pro Pro
165 170 175
Gly His Leu Tyr Ser Ser Lys Asn Gly Gly Phe Arg Arg Trp Tyr Asn
180 185 190
Pro Leu Trp Phe Ser Glu Ala Ile Pro Ser Thr Pro Tyr Asp Pro Leu
195 200 205
Val Leu Arg Arg Ala Phe Glu Asn Ala Val Ile Lys Arg Leu Met Thr
210 215 220
Asp Val Pro Phe Gly Val Leu Leu Ser Gly Gly Leu Asp Ser Ser Leu
225 230 235 240
Val Ala Ser Ile Thr Ala Arg Tyr Leu Ala Gly Thr Lys Ala Ala Lys
245 250 255
Gln Trp Gly Ala Gln Leu His Ser Phe Cys Val Gly Leu Glu Gly Ser
260 265 270
Pro Asp Leu Lys Ala Ala Arg Glu Val Ala Asp Tyr Leu Gly Thr Val
275 280 285
His His Glu Phe His Phe Thr Val Gln Asp Gly Ile Asp Ala Ile Glu
290 295 300
Asp Val Ile Tyr His Ile Glu Thr Tyr Asp Val Thr Thr Ile Arg Ala
305 310 315 320
Ser Thr Pro Met Phe Leu Met Ser Arg Lys Ile Lys Ser Leu Gly Val
325 330 335
Lys Met Val Ile Ser Gly Glu Gly Ser Asp Glu Val Phe Gly Gly Tyr
340 345 350
Leu Tyr Phe His Lys Ala Pro Asn Lys Glu Glu Phe His Lys Glu Thr
355 360 365
Cys Arg Lys Ile Lys Ala Leu His Gln Tyr Asp Cys Leu Arg Ala Asn
370 375 380
Lys Ser Thr Ser Ala Trp Gly Leu Glu Ala Arg Val Pro Phe Leu Asp
385 390 395 400
Lys Glu Phe Ile Asn Val Ala Met Ser Ile Asp Pro Glu Trp Lys Leu
405 410 415
Ile Lys Pro Glu Gln Arg Arg Ile Glu Lys Trp Ala Leu Arg Arg Ala
420 425 430
Phe Asp Asp Glu Glu His Pro Tyr Leu Pro Lys His Ile Leu Tyr Arg
435 440 445
Gln Lys Glu Gln Phe Ser Asp Gly Val Gly Tyr Ser Trp Ile Asp Gly
450 455 460
Leu Lys Ala His Ala Glu Gln His Val Thr Asn Arg Met Met Leu Asn
465 470 475 480
Ala Ser His Ile Phe Pro His Asn Thr Pro Ile Thr Lys Glu Ala Tyr
485 490 495
Tyr Tyr Arg Met Ile Phe Glu Arg Phe Phe Pro Gln Asn Ser Ala Gly
500 505 510
Leu Thr Val Pro Gly Gly Ala Ser Val Ala Cys Ser Thr Ala Lys Ala
515 520 525
Val Glu Trp Asp Ala Ser Trp Ser Lys Asn Leu Asp Pro Ser Gly Arg
530 535 540
Ala Ala Ile Gly Val His Asn Ser Ala Tyr Glu Asn His Glu Pro Ala
545 550 555 560
Met Ala Asn Gly Asn Leu Ala Thr Lys Ile Ile Gly Arg Ala Pro Ser
565 570 575
Met Val Glu Val Gly Ala Ala His Glu Leu Thr Ile Arg Ser
580 585 590
<210> 5
<211> 5276
<212> ДНК
<213> Nicotiana tabacum
<400> 5
atgtgtggaa tcttggcttt gttgggttgt tcagatgatt ctcaggccaa aagggttcga 60
gttcttgagc tctctcgcag gcaatttccc ctttcttttc tcttttattt tattactcca 120
tcatataatt attctacttt cacctttttt gttttttaat ttcgatttgt ttgctttttg 180
ctttcttcat ttaatgttct tgcattgctc tagctcctat acgaataata ggagctaaag 240
taatagagtg aaggattatt ttggagcaac ggtaaagtta tttttgtata gttcacaggt 300
tggagccgta aaagcagtta ctagtgtttg cattagggta agttgtctat caggaaagat 360
atttgtagca attgaccacc ctttggcaaa aaattatgcg taaaatatta ctacttgtta 420
ttgattcaaa aaatatgact ttgaacatcc ttgtatagcc tagtggcaaa gggtgttcaa 480
aatttttgaa cacctttatt gaattttctg gtttcgtcgt tgctgtctac aacacactct 540
cgaagtgcgg ccttccctca aaccctacgt gaatatggaa cgcctagtgt gccgggatgc 600
cctttaggct gagtaataga gtactatgtt gatataccta aaatacggaa ctggcacgag 660
agataaattc agaatttata gcttaattat ttaaaattta tcatttgatt tatttcaacc 720
agagttcatt ctagtagagt ttatgttgca tatttgtctg atttgcacca tttgttttaa 780
tgttcttgca ttactctaat ttatttatga ataataagct aaactaattg agtttatgtt 840
gatgtaactg acctgagttg gcagtgatgt ttggctactt gaatttaaaa caagattaag 900
acttgtgagt tggcagtaat tgtttggata cttgaattta gaacaagaat aaaacaagat 960
taattaagga tacattattc ctaacccgga ccctaattct atagattgat tattaaattt 1020
ttcttggacc aaccgtcacc gtgacctgtt catgatggtg atggcaaaag ccataataca 1080
cttgatttct gttttcaatt tttaaggttc ttattattat acttattata tactttgaaa 1140
attatgggtc aaaattttaa tatttgttat tccttccgta tgtttgactc aacttggagt 1200
tcatgaaaaa aaaaaaaaat ttttttaaac ttgtggtctt aaacaagtca tcgatatatg 1260
tgtggttgtg aatcgcctcg ttaggataaa gtagaaattt taaagttaaa ctatttcgaa 1320
ataaaaaagt atgatagtgt ttttgggaca tactaaaaag gaaataacac ataaaatgga 1380
acaaagaagt atattttagt gatttttcac atatatatca gtattttgta tcaaaaatgc 1440
tggttcagtt gaactcattg aactgggtgc tccatccacc tccgagtact gcaacatgtg 1500
accggtggca gatgcaaccc ttacgtacca gggtcatctg aacctagtat aaatatattt 1560
gtgaaaaatc cctaaaattt caataattag tagatttaaa accttaattt taaaactaca 1620
atgagttcag ttttaaaatc tttatatgtc aaacctatca agattaaatc ctggatccac 1680
cttttgcatg tgacagtaca atagcctttt gtaattgaaa caaatatttg aattttggta 1740
actatttgat cacataggtt gaagcatcgt ggaccagatt ggagtgggat atatcaacat 1800
ggtgattttt acttagcaca tcaacgttta gcaattatcg atcctgcttc tggtgatcag 1860
cctctgttta atcaagataa gacgattgtt gttacagtga gtgcctctaa tttacctctt 1920
ttttttttct tttttttttt acgatcgcgt tgtttagatt atttatggat gaaaatttgt 1980
atatttgatt attgattgtg tgctaaattt tgcaggtcaa tggagagatt tacaatcatg 2040
agaaacttcg taatcttatg cctaatcaca agttcagaac tggaagtgat tgtgatgtta 2100
ttgcacatct tgtgagttag tttacttttc tcattgacca acttaggcca gtgaattata 2160
ttaggaattt cgggtgtttg gatatggtca aattagctta taagcactta ttatcaattt 2220
taacattttt atccatacac gtaactgttc attcataaag tcattttagc acttgatgct 2280
catcaaccta tgttgttggg actcttcgaa aatgtcagcc cgatagcatt tcggagagtc 2340
cgagcaacat agcaatcaac tacttttaat cagttaaacg aacatgctca tagtgataac 2400
ctctggaaaa tatgttatag taaattagaa aaatttatac gatgtctcac tatcggtatg 2460
tattgatcag tatgaagaat atggagaaaa ttttgtggac atgttggatg gggtgttctc 2520
ttttgtattg ttggatacgc gcgataacag ctttcttgct gctcgtgatg caattggaat 2580
tactccccta tatattggtt ggggacttga tggtaagatt ttctatacaa ttttccagtt 2640
agaaattaaa actgaagcta ttcctattta ctattgaaga atcttgatac atgttatttg 2700
gtatttcgtc gtgttgttat gtctcaggct ctgtgtggat ttcatctgag ctaaaaggat 2760
taaatggtga ctgtgaacat tttgaagttt tccctcccgg tcacttgtac tcgagcaaga 2820
atggcgggtt taggagatgg tacaatcctc aatggttctc tgaggctatt ccatcaaatc 2880
cttacgaccc cttagttttg agacgtgcct tcgaaaatgt gaggcttgtt aatgaatcta 2940
gtgtacaatc ttggatgttt ttctctcgac tttggtttta gcactcaaaa agttgtctgt 3000
acaaacgatc ttggctataa ggagtggtgg ttaattttgc aggctgttat caaacgattg 3060
atgaccgatg tcccctttgg tgttctgctc tccgggggac ttgattcgtc tttggttgct 3120
tctgtcactg ctcgctactt ggctggaaca aaagctgcta agcaatgggg agcacagctt 3180
cattccttct gtgttggtct cgaggtcaga ctatcaatcc tgagcggaga catatttggg 3240
gcgggttctg tgggttcaac tgaacccatt gcttttggtt cgaaccatat atacaaatcc 3300
aagaaaatat aagatgtgta catataatat gacactcatt ctgagcctac tgacgctaga 3360
tactggactc gcctttggtc ttgagacttt ataatagaaa atagatattc aataccaatt 3420
tctgtttttc ttttctgact gacagggctc accagatctc aaggctgcaa gagaagttgc 3480
tgactatttg ggaaccgttc accacgagtt caccttcaca gttcaggttt gtaaatcgtg 3540
ttgctactcc gatttattca tgtgagtctc ttctgttttc ctcctagaaa gcaacaagtg 3600
ttcatgtttt gcaggatgga attgatgcta ttgaagatgt tatttaccat atcgagacat 3660
acgatgtaac aacgatcaga gcaagcactc ctatgttcct tatgtcgcgt aagattaaat 3720
cactgggagt taagatggtc atatcagggg aaggctcaga tgaactgttt ggcggctatt 3780
tgtacttcca caaggctccg aacaaggaag aattccatgt ggagacatgt cacaaggtaa 3840
taaaacacgt cggctccctt ggcaagtctg attctccttg tcattgtttg tctcgggttg 3900
gaataaattg ctataattcg agaagaaatg ttaatcctgc tctttcactc gcatgcagat 3960
aaaagcgctt caccaatacg actgtttgag agcaaataag gcaacatcag catggggctt 4020
agaagctaga gtaccatttc tggataaaga gttcatcaac gttgctatga gtatcgatcc 4080
tgaatggaag atggtaacaa aactgagctc caattgtcaa tgagatgaga agcataactg 4140
aatatgcacg ttctctaaaa cttgtttttc atccgtttca gattaaacac gatcatggta 4200
ggatcgagaa gtgggttctt aggaaggctt ttgatgatga ggagcaaccc tatctcccaa 4260
aggtccacaa agtcatgttt ttcctactga tttgccgatt agtatggatc aaaactaaca 4320
tgttttgttc ttttatgtac tctgcagcat attctgtacc ggcagaaaga acaattcagt 4380
gatggcgtag gctatagttg gatcgatgga ctcaaagcac atgctgaaca acatgtgagc 4440
ttctaaataa atccagaact attacatcgt tatcgtccaa ttttcgtttg ttttgctgtc 4500
agctgatctt ggaaacaatt cttttctctc aggtgactga taggatgatg cttaatgctg 4560
cacatatctt ccctcacaac actccaacta caaaggaagc atactattac aggatgattt 4620
tcgagaggtt cttcccacag gtacttatag cgcgagacta gaatcatata tctcttaaat 4680
ggaaacatga aaaataaagg aggtgttttt tctccttgga aaaagtattt actactgatt 4740
aaagcacctt cattcaccca tcttgctact tcgcgtactt tactaagttg tggttctaaa 4800
aaagaagatt tagcttggta tattccatca cgaggaagcc acaaaaagat tctacttata 4860
agaagttgga tacttttaat ggtgtgagac cttttgggaa aaaccatgcg ggcttggccc 4920
aaagccgaca atatcacacc atgttaagag tatctttgct tcgttttagc caaacagtac 4980
aataagtcaa tttttaaacc atgttgttac tattgttgga tttgcagaat tcagcaaggc 5040
taactgttcc tggaggaccg agtatagctt gcagcacagc taaagctatt gagtgggatg 5100
cttcgtggtc gaacaacctt gatccttccg gtagggctgc aatcggtgta cataactcgg 5160
cttatgacga tcatctcccc gatgttggta atgggaattt ggacacaacg atcatcgata 5220
atgtgccgag gatggtagga gtgggtgctg ctgcagagct cacaataagg agctag 5276
<210> 6
<211> 590
<212> PRT
<213> Nicotiana tabacum
<400> 6
Met Cys Gly Ile Leu Ala Leu Leu Gly Cys Ser Asp Asp Ser Gln Ala
1 5 10 15
Lys Arg Val Arg Val Leu Glu Leu Ser Arg Arg Leu Lys His Arg Gly
20 25 30
Pro Asp Trp Ser Gly Ile Tyr Gln His Gly Asp Phe Tyr Leu Ala His
35 40 45
Gln Arg Leu Ala Ile Ile Asp Pro Ala Ser Gly Asp Gln Pro Leu Phe
50 55 60
Asn Gln Asp Lys Thr Ile Val Val Thr Val Asn Gly Glu Ile Tyr Asn
65 70 75 80
His Glu Lys Leu Arg Asn Leu Met Pro Asn His Lys Phe Arg Thr Gly
85 90 95
Ser Asp Cys Asp Val Ile Ala His Leu Tyr Glu Glu Tyr Gly Glu Asn
100 105 110
Phe Val Asp Met Leu Asp Gly Val Phe Ser Phe Val Leu Leu Asp Thr
115 120 125
Arg Asp Asn Ser Phe Leu Ala Ala Arg Asp Ala Ile Gly Ile Thr Pro
130 135 140
Leu Tyr Ile Gly Trp Gly Leu Asp Gly Ser Val Trp Ile Ser Ser Glu
145 150 155 160
Leu Lys Gly Leu Asn Gly Asp Cys Glu His Phe Glu Val Phe Pro Pro
165 170 175
Gly His Leu Tyr Ser Ser Lys Asn Gly Gly Phe Arg Arg Trp Tyr Asn
180 185 190
Pro Gln Trp Phe Ser Glu Ala Ile Pro Ser Asn Pro Tyr Asp Pro Leu
195 200 205
Val Leu Arg Arg Ala Phe Glu Asn Ala Val Ile Lys Arg Leu Met Thr
210 215 220
Asp Val Pro Phe Gly Val Leu Leu Ser Gly Gly Leu Asp Ser Ser Leu
225 230 235 240
Val Ala Ser Val Thr Ala Arg Tyr Leu Ala Gly Thr Lys Ala Ala Lys
245 250 255
Gln Trp Gly Ala Gln Leu His Ser Phe Cys Val Gly Leu Glu Gly Ser
260 265 270
Pro Asp Leu Lys Ala Ala Arg Glu Val Ala Asp Tyr Leu Gly Thr Val
275 280 285
His His Glu Phe Thr Phe Thr Val Gln Asp Gly Ile Asp Ala Ile Glu
290 295 300
Asp Val Ile Tyr His Ile Glu Thr Tyr Asp Val Thr Thr Ile Arg Ala
305 310 315 320
Ser Thr Pro Met Phe Leu Met Ser Arg Lys Ile Lys Ser Leu Gly Val
325 330 335
Lys Met Val Ile Ser Gly Glu Gly Ser Asp Glu Leu Phe Gly Gly Tyr
340 345 350
Leu Tyr Phe His Lys Ala Pro Asn Lys Glu Glu Phe His Val Glu Thr
355 360 365
Cys His Lys Ile Lys Ala Leu His Gln Tyr Asp Cys Leu Arg Ala Asn
370 375 380
Lys Ala Thr Ser Ala Trp Gly Leu Glu Ala Arg Val Pro Phe Leu Asp
385 390 395 400
Lys Glu Phe Ile Asn Val Ala Met Ser Ile Asp Pro Glu Trp Lys Met
405 410 415
Ile Lys His Asp His Gly Arg Ile Glu Lys Trp Val Leu Arg Lys Ala
420 425 430
Phe Asp Asp Glu Glu Gln Pro Tyr Leu Pro Lys His Ile Leu Tyr Arg
435 440 445
Gln Lys Glu Gln Phe Ser Asp Gly Val Gly Tyr Ser Trp Ile Asp Gly
450 455 460
Leu Lys Ala His Ala Glu Gln His Val Thr Asp Arg Met Met Leu Asn
465 470 475 480
Ala Ala His Ile Phe Pro His Asn Thr Pro Thr Thr Lys Glu Ala Tyr
485 490 495
Tyr Tyr Arg Met Ile Phe Glu Arg Phe Phe Pro Gln Asn Ser Ala Arg
500 505 510
Leu Thr Val Pro Gly Gly Pro Ser Ile Ala Cys Ser Thr Ala Lys Ala
515 520 525
Ile Glu Trp Asp Ala Ser Trp Ser Asn Asn Leu Asp Pro Ser Gly Arg
530 535 540
Ala Ala Ile Gly Val His Asn Ser Ala Tyr Asp Asp His Leu Pro Asp
545 550 555 560
Val Gly Asn Gly Asn Leu Asp Thr Thr Ile Ile Asp Asn Val Pro Arg
565 570 575
Met Val Gly Val Gly Ala Ala Ala Glu Leu Thr Ile Arg Ser
580 585 590
<210> 7
<211> 5926
<212> ДНК
<213> Nicotiana tabacum
<400> 7
atgacgtagt ttagcttgtt cgtccacggc ataaaaacgc ctataaattc ggtggtcatt 60
ttggttcatt tacagaatcc cacgatcatc ttttgtgtat tatcattttg ccttctttac 120
gtgttcttct ctcccaaaat cctgcaaatc tttaactatc ttcccatatt tctctccctc 180
tttctctttc tttttgttat ctactttact acttctatta gtagagcttt atccaattaa 240
agtcatctat attggatata gtttttacta ctactttatc ttgtttggga aagaaaagaa 300
taaaaaatta gagtgtctat attaagtata aaaaggtgaa aaataaaaaa gtagtgatca 360
atcatgtgtg ggatcttggc tttgttgggt tgtccagatg attctcaagc caaaagggtt 420
cgagttcttg agctctctcg caggcaattc ccctttcttt tctcttttat tttattactc 480
catcatatta ttcaactata atttcacctt ttttgttttt caattaatat ttgtctgctt 540
ttgctttctt catttaatgt tcttgcattg ctctagttcc tttatgaata ataggctaaa 600
gtaatagagt gaaggagtag agtcggagca acggtgaagc tgtctttaca tgatctatag 660
ctcacgggtt caccatagaa gcagccaatg gcgaagctag aaaattcaat aagggtgttc 720
tttcctttgc cagtgggcta tgcaagggtg ttcaaagtct atttttaatc aataacaagt 780
aatattttat cttatagtga aagcaaccac taatacttgc atttaggtaa ttagctgtct 840
atcaagggaa gttatgtgtg gtaagtagtg gcggagccag aattttcgtt aaggggtgtc 900
aaaatatata aaagtaaaca taccaggaaa ttaaggggag tcaatacata gtatatatac 960
atataattta tttttttacc tagctataca gtatattttt tccgcgaagt gacacccctt 1020
cctataaggt ggctccgcca ctggtggtaa gggtggtcaa ttgaacaccc ttcgccgaaa 1080
aattatactc tggatggtgt aaaatattac tacttgttac tgattcaata aataaatttt 1140
gaaaaccctt gcatagtcta gtagcaaaga gtgttcaaaa ttttgaacac atttattgaa 1200
ttctctggtt tcgccactgc tgtctacatc acatccctta aggtgcgatc ttttttcgaa 1260
tcatgtgtga atacgggatg ccttgtgttc caggctgccc ctttaggcta aagtaataga 1320
gtacgtctta attcttatgt tgatcatata cctgaaactc ggaactggca cgagagatat 1380
aaattcagaa tttatagcaa atttatcatt atgttgcatt tgttttaatg ttcttgcatt 1440
actctaattt ttttatgaat aataagctaa actaattgag tttatgttga tgtaactgac 1500
ctgagtttgg ctacttgaaa gttaaaacaa gattaagact tgtgagttgg cagcgattgt 1560
ttggatactt gaacacgttt aaaacaagat taagacatta tgatggatac attattccta 1620
acctgggtgg gacctaattc tatggattca ttattaaaat ttcttggaca aaccatcacc 1680
atgatctgtt catgatgttt atggcaaaag ccataataca ctttatttct gtatcacaac 1740
tactttatac acatgttata gagttttaaa attattttaa tatttgttat tccttgagtc 1800
tcctgatatt tgactccgca tgaaattcaa gaaaaaaaaa agaagatttt taaaatttat 1860
agtcgaaaac aagccatcga tatatgtgtg gttgttaata acctcgttag gcgtaaagta 1920
gaaattttaa agttaaacta tttctaaata agaaaatatg atattgtttt tgcatagact 1980
aataaggaaa gtatgacaag taaaataaaa caaagaagta tattttaatg attttttccg 2040
tgtcaaaaat gtcagtttag ttgaacttga actcattgaa actggtgacc agtggtggat 2100
gcaaacctta ggtacggagg tcatccgaac ctagtatagt tatatttgtg aaaaatcact 2160
aaaatttcaa taattactag atatgaaacc ttaattttaa aactacaatg agtgcagttt 2220
taaaatcttt aaatgtcaaa ccgatcaaga ttaaattatg gatccacctt taaatgtcaa 2280
accgacagta caatagcctt ttgcaattga aacaaatata aacattacat ttattaaact 2340
agttttaatt ttggttacta tttgattaca taggttgaag catcgtggac cagattggag 2400
tgggatatat caacatggtg atttttactt agcacatcaa cgtttagcaa ttatcgatcc 2460
tacttctggt gatcagcctc tgtttaatca agataagact attgttgtta cagtgagtgc 2520
ctctaattta cccttttttt tgtggacgaa tgcgttgttt agattacttt aattatggct 2580
gaaaatttgt atatttgatt attgattgtg tgccaaatgt tgcaggtcaa tggagaaatt 2640
tacaatcatg agaaacttcg taatcttatg cctaatcaca agttcagaac cggaagtgat 2700
tgtgatgtta ttgcacatct tgtgagttag tttactttac tcatcaacca acttaggcta 2760
gtgaattcaa aggcggagct agaattttga gtatatgagt tctggaattt aggacaagat 2820
aagttactgg gttcggatag attatttaga catattaagt agatttctta acacaaatac 2880
acggcccgag ccgaagctat tgggttctgc cgaacccgta gctagacttg tagctccgcc 2940
attgagtgaa ttatataagg aatttcgggt gtttgggtaa acttataagc tggtcaaatt 3000
agcttataag cacttattat caatttcaac atttctatcc aaacacgtaa ctattcattc 3060
gattttagca cttgatgctt atcgactact tttaatcagt taaacgaaca ggctcatagt 3120
gacaaactct agaaaatatg gtattgtaaa ttagtagaag aatttatacg atgtctcact 3180
atcgatatgt attgatcagt atgaagaata tggagaaaat tttgtggaca tgttggatgg 3240
ggtgttctct tttgtattgt tggatacgcg cgataacagc tttcttgctg ctcgtgatgc 3300
gattggaatt actcccctct atattggttg gggacttgat ggtaagattt tctatacaat 3360
tttccaagta gaaattaaaa ctgaagccat tccaatttgc tattgtagaa tcttgatata 3420
tgttatttgg tatttcaggc tctgtgtgga tttcatctga gttaaagggc ttaaatgatg 3480
attgtgaaca ttttgaagtt ttccctcccg ggcacttgta ctcgagcaag aatggcgggt 3540
ttaggagatg gtacaatcct caatggttct ctgaggctgt tccatcaaat ccttatgacc 3600
ccttagttct gaggcgtgcc ttcgaaaatg tgagaaatga atctaggagg tttccctctc 3660
gactttggac ttagcactct aaaaatttgt ctgtacaaac gatcttggct ataaggagtg 3720
gttgttaatt tcgcaggctg ttattaaacg gttgatgacc gatgtaccct ttggtgttct 3780
gctctccggg ggacttgatt cgtctttggt tgcttctgtc actgctcgct acttggctgg 3840
aacaaaagct gctaagcaat ggggagcgca gcttcattcc ttctgtgttg gtctcgaggt 3900
cagtcaatca atcttgagta gaaaaaattt agggtcggtt ctgtgggttt aacttaactc 3960
attgcttttg gctcgaacca tatatgcaaa tccaaaaaga tttaagatgt atacatataa 4020
tataacactc attctgagcc tactgacgct agatactgga ctgtgccttt ggtcttgaga 4080
cttttgtaat agaaaattga tattcaatac caatttctgt tcttttgtga ctaacagggc 4140
tcaccagatc tcaaggctgc aagagaagtt gctgactatt tgggaaccgt tcaccacgag 4200
ttcaccttca cagttcaggt ttgtaaatcg cgatgctact tcaatttatt catgtgaggc 4260
tcttctgttt tcctactaga aagcaacatg tgtgcatgtt ttggcaggac ggaattgatg 4320
ctattgaaga tgttatttac catatcgaga cgtatgatgt aacaacgatc agagcaagca 4380
cccctatgtt ccttatgtcg cgtaagatta aatcacttgg agtgaagatg gtcatatcag 4440
gggaaggctc agatgaactg tttggtggct acttgtactt ccacaaggct cccaacaagg 4500
aagaattcca cacggagaca tgtcacaagg taataaaaca catcggcaag tctgattctc 4560
cttgtcattg tttgtctcgg gttggaataa attgctataa ttcgagatat ggaatgagtt 4620
aagaaatgtt aatcatgttt ttccattcgc atgcagataa aagcgcttca ccaatacgac 4680
tgtttgagag caaataaggc aacatcagca tggggcttag aagctagagt accatttctg 4740
gataaagagt tcatcaatat tgctatgagt atcgatcctg aatggaagat ggtaagaaaa 4800
ctgagctcca attgtcaatg agacgagaag cataactgaa tatgcgcgtt ctctaaaacc 4860
tgtttttcat ccgtttcaga ttaaacacga tcaaggtagg atcgagaagt gggttcttag 4920
gaaggctttt gatgatgagg agcaccccta tctcccaaag gtccacaaac tcatattttt 4980
cctactgatt tgccaataat tatggatcaa aactaacatg tttgttctgt tatgtgctct 5040
gcagcatatt ttgtaccggc agaaagaaca attcagcgat ggtgtaggct atagttggat 5100
cgatgggctc aaagcacatg ctgaacaaca tgtgagcttc cttaataaat ccagaactaa 5160
tgcagcgtta tcgtccaatt tttgtttgtt ttactatcaa ctaattttag tcatgttagt 5220
atgtttacct taggtactat gtttttaagg agtttttagt cgtgtcaaga atttattcgc 5280
tggtaaaaaa tagattgaac ttccagtatt ttctactttt attatgcata tgattggact 5340
gagatgaggt taaatggaac aacatggttg aggattaata taaccaattc caacttggtt 5400
ggaattgagg cgtcgtagtt gttgttagat ctcagaaaca attcttttct ctcaggtgac 5460
tgataggatg atgcttaatg cttcacatat cttccctcac aacactccaa ctacaaagga 5520
agcatactat tacaggatga tttttgagag gttctttcca caggtaatat tagtgagaga 5580
ttaaaatcta tagattttgt ttatggctga ggtttaaatc atagatttct taagctcaaa 5640
aactaaaacc atgtccctgc tactgttgga tttgcagaat tcagcaaggc taactgttcc 5700
tggaggaccg agtatagctt gcagcacggc taaagctatt gagtgggacg cttcttggtc 5760
gaacaacctt gatccttccg gtagggctgc tatcggtgta cataactcag cttatgacga 5820
tcatctaccc gatgttagta atgggaattt ggacacaacg atcatcgata atgtgccaag 5880
gatggtagga gtgggtgctt ctgcagagct cacaataagg agctag 5926
<210> 8
<211> 565
<212> PRT
<213> Nicotiana tabacum
<400> 8
Met Thr Leu Lys His Arg Gly Pro Asp Trp Ser Gly Ile Tyr Gln His
1 5 10 15
Gly Asp Phe Tyr Leu Ala His Gln Arg Leu Ala Ile Ile Asp Pro Thr
20 25 30
Ser Gly Asp Gln Pro Leu Phe Asn Gln Asp Lys Thr Ile Val Val Thr
35 40 45
Val Asn Gly Glu Ile Tyr Asn His Glu Lys Leu Arg Asn Leu Met Pro
50 55 60
Asn His Lys Phe Arg Thr Gly Ser Asp Cys Asp Val Ile Ala His Leu
65 70 75 80
Tyr Glu Glu Tyr Gly Glu Asn Phe Val Asp Met Leu Asp Gly Val Phe
85 90 95
Ser Phe Val Leu Leu Asp Thr Arg Asp Asn Ser Phe Leu Ala Ala Arg
100 105 110
Asp Ala Ile Gly Ile Thr Pro Leu Tyr Ile Gly Trp Gly Leu Asp Gly
115 120 125
Ser Val Trp Ile Ser Ser Glu Leu Lys Gly Leu Asn Asp Asp Cys Glu
130 135 140
His Phe Glu Val Phe Pro Pro Gly His Leu Tyr Ser Ser Lys Asn Gly
145 150 155 160
Gly Phe Arg Arg Trp Tyr Asn Pro Gln Trp Phe Ser Glu Ala Val Pro
165 170 175
Ser Asn Pro Tyr Asp Pro Leu Val Leu Arg Arg Ala Phe Glu Asn Ala
180 185 190
Val Ile Lys Arg Leu Met Thr Asp Val Pro Phe Gly Val Leu Leu Ser
195 200 205
Gly Gly Leu Asp Ser Ser Leu Val Ala Ser Val Thr Ala Arg Tyr Leu
210 215 220
Ala Gly Thr Lys Ala Ala Lys Gln Trp Gly Ala Gln Leu His Ser Phe
225 230 235 240
Cys Val Gly Leu Glu Gly Ser Pro Asp Leu Lys Ala Ala Arg Glu Val
245 250 255
Ala Asp Tyr Leu Gly Thr Val His His Glu Phe Thr Phe Thr Val Gln
260 265 270
Asp Gly Ile Asp Ala Ile Glu Asp Val Ile Tyr His Ile Glu Thr Tyr
275 280 285
Asp Val Thr Thr Ile Arg Ala Ser Thr Pro Met Phe Leu Met Ser Arg
290 295 300
Lys Ile Lys Ser Leu Gly Val Lys Met Val Ile Ser Gly Glu Gly Ser
305 310 315 320
Asp Glu Leu Phe Gly Gly Tyr Leu Tyr Phe His Lys Ala Pro Asn Lys
325 330 335
Glu Glu Phe His Thr Glu Thr Cys His Lys Ile Lys Ala Leu His Gln
340 345 350
Tyr Asp Cys Leu Arg Ala Asn Lys Ala Thr Ser Ala Trp Gly Leu Glu
355 360 365
Ala Arg Val Pro Phe Leu Asp Lys Glu Phe Ile Asn Ile Ala Met Ser
370 375 380
Ile Asp Pro Glu Trp Lys Met Ile Lys His Asp Gln Gly Arg Ile Glu
385 390 395 400
Lys Trp Val Leu Arg Lys Ala Phe Asp Asp Glu Glu His Pro Tyr Leu
405 410 415
Pro Lys His Ile Leu Tyr Arg Gln Lys Glu Gln Phe Ser Asp Gly Val
420 425 430
Gly Tyr Ser Trp Ile Asp Gly Leu Lys Ala His Ala Glu Gln His Val
435 440 445
Thr Asp Arg Met Met Leu Asn Ala Ser His Ile Phe Pro His Asn Thr
450 455 460
Pro Thr Thr Lys Glu Ala Tyr Tyr Tyr Arg Met Ile Phe Glu Arg Phe
465 470 475 480
Phe Pro Gln Asn Ser Ala Arg Leu Thr Val Pro Gly Gly Pro Ser Ile
485 490 495
Ala Cys Ser Thr Ala Lys Ala Ile Glu Trp Asp Ala Ser Trp Ser Asn
500 505 510
Asn Leu Asp Pro Ser Gly Arg Ala Ala Ile Gly Val His Asn Ser Ala
515 520 525
Tyr Asp Asp His Leu Pro Asp Val Ser Asn Gly Asn Leu Asp Thr Thr
530 535 540
Ile Ile Asp Asn Val Pro Arg Met Val Gly Val Gly Ala Ser Ala Glu
545 550 555 560
Leu Thr Ile Arg Ser
565
<210> 9
<211> 21008
<212> ДНК
<213> Nicotiana tabacum
<220>
<221> misc_feature
<222> (18072)..(18072)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (19355)..(19462)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (19541)..(19541)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (19714)..(19786)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (20035)..(20035)
<223> n представляет собой a, c, g или t
<400> 9
atgtgtggaa tactagctgt ttttggttgc attgataact ctcaggccaa gcgttcccga 60
atcatcgaac tttccagaag gtttcttctt tttattgctt tccgattttc tacagcattc 120
ttattcttct ggggtgtcaa tggtatgcat gtgttttgga tttgctattc gtgaacttgt 180
tcgctttctc gtggtcttcg atttattttt gtcaatttat aatcagcgtt ttttgtatgg 240
atttctcttt tgaatctctc aaatatatga catttgattt cctcgagaaa atttggcatt 300
tttatggttc ttatgatatg tggttaaccc ctcgctctgc atcgcttcag cgctttaaaa 360
cacgaagtga tggctattag taatactgct agagtggtta ttgaattaca ttaaataacg 420
ggttgctagt agatcaaatt cagaatgaca gtgaaatatc tctggaaact tgaatccccg 480
ttcgattcgc tggtaaaacc tggctaaaaa atagctttat ctcaaaggag agtatgtcac 540
tgtgaaattg gtggtatgtg ttgtttaagg catgatatgc tgctcttagg tgatgtcaag 600
atgataatac catccttttc acactagact cagctttgta gcacttacat ctcagttaat 660
cgatggagga aaatctagtc tgctttgggg catattatac caaatgtctt ccttttgttt 720
tctagtgttg tttctcaaga attgaaaaac caatctacaa tatagtgata taattaacta 780
gttaactctt cctgtgtttt ctgaaaattt ccaataattg agaagtgttt actggtcaat 840
tagtaaaggg agaacggaat atttttttcc tctaaagaac ttctcttttt ccatacagaa 900
tgactttcta tacaaggcct aagtctaatt tagtgtctgc gattaagcat cttctgggaa 960
cttcacactt gaaagaagta gaactaggag ggggcaggtc tagaaagata attgcttcag 1020
tcatttctca taaaaaacat tgcttcagtc attatgtgca ttcatataga gttggtcaag 1080
ctgctaagct gagctaatag tatgatagct gtgactctcg acaaagtagg ttgcctctta 1140
gggctttatc ctttggctcc gtaaagaatt tctttgggag ggctccagag tgggtaggat 1200
tgttctcatg gtaagctgta acaccaacga aacaagcctt tgtagctgtg gaagtccgag 1260
ggggtggtgg tggtggtggt ggtgtgagag ttagagtggt ggaggagagg ggaggagtga 1320
agagaagttt atattttaaa gaatagggcc ttgagaactt ttagtagtga gcatgcatta 1380
ttattattgt tattgttatt attattatta ttattattat tattattata ttattattat 1440
tattatatta ttgttatgta attacttagc taatgaggtt tttccaactg tttccaaatt 1500
aggaaatgtg ttgaaagcat agttaagtat ataaaagcgt gttctttcta atatccttag 1560
cttttagagc agataatcac aaaatttatc atgataatat agcagacaat ttttcctagt 1620
tcaaatctca tgccaccctt cataaaaaat ttccacgtgc ttggctggca tgttagggga 1680
cgtcttggca taattaaaca aatagaattg tccctttcta acagcttaag cttttagatg 1740
aggcggtcaa caattcaaca atttgcattt gtttgataag aagatattaa aagaacctta 1800
aaaggtgtca gtatgtgaac aaactcaagg ggtagactaa ctagaagtca atttaaattt 1860
tctgcttgtc tgtgttttac ttcttttgca ctatagaaat tgcatttcca gcattccgaa 1920
agctgcttcc tttgcatata gacttcacat ttccagcgtt tctgataagc gttcttgctt 1980
attcctgtcc ttctacttgg ttgattctcc tctcacttag ttcactatct tcttgcacgc 2040
attttcaaat tgttttatca attctcttgc tttcttttca tctgacccca caaaatacct 2100
cccaaaagcc tttctgcttt tccgagtctg aaattatctg cttattactg cctttcaagt 2160
attctgatgc gttgactgtt tttttccctt gtagtaatgc atggttcgct gtccaatctc 2220
tattttgttt tttgtgaacc actattcttt gttgcttctg tttcgtcatc ctcagttccc 2280
catcttctca gtttgatttt cctcatttaa aaagcgtgtg ccccttttct tgtgctaccc 2340
tctaaaaaat cagttgctgt catctcgttt gatctgtcat gatcccagtt gatgccacat 2400
gtttagttct ttacctctgg acaactttta cttcctgaga accacgattt cttttcctga 2460
agaaattctt agaaattggt tatgtaaagg aaattcccag caatcaccat ttggcctcta 2520
cctggcttgc ttttcaactt accgacacgt gttcatcccc ttttaacaga tgctgaccat 2580
tatcttctgc ttgttggggc atacttctta cccatggtgt taaccctgat ctttcgtatc 2640
acatttttca actcttaatt ctcacatgta aagtgcctat ctttgcctct ttagttgatc 2700
ttgggttggg ctgttaatcc aaattctctg ggctttgctt tgaagtatga taagcggtag 2760
ctttactaca gtaccatatt ctacattgct acagacgtga aattatcctg atttgtcttt 2820
gatgtgatgt ctggcaagtg tagattacgc cacagaggac ctgattggag tggattgcat 2880
agccatgatg actgttatct tgctcatcaa cggttggcaa tagtagaccc aacttctgga 2940
gatcagccac tttacaacga ggacaagacc attgttgtcg cggtatggaa atatttatta 3000
tattttttct tagatattac cggtacatgg actttggaca atatttgtgc gttttgatct 3060
tctatagtac aaacttttct cgtaggataa atttcctcta ctttgaatgg atcttaacag 3120
gaaaatcgga ataaacaatt tgtgatttcg aaatttttag ttcttacata tttggaacat 3180
atgcggtaaa gtaactaggc gattgactag gtgaatacgg agaatttcct ctactgttat 3240
caatatttac tcagctcatt tgaaagatct ctgggtcgtc tagcagtaca gctatttagg 3300
caacttaacc aaatcccctt cccatgtgtt tcactttctt gacttatttt atattttgac 3360
attttgcaat gccgaaaata aattgttttt ccattttatg acctttaaaa tgttgaggtg 3420
aagggctagg cggtacttta attgacttat cagcaacaat gtattagata cagagcacaa 3480
acttgtttgg gtgcctaatt tggtattttt tgagaacgat ggagtaaatg gaaatatgaa 3540
ttagcaaaat attgaaatag tagatctgaa aaatgaatga ctgaatttag aggaggacct 3600
tttcttcttt ccttgctgaa gtggttataa gagaattatg tgaagaacat gtaagcttta 3660
gggtagtgtt acaccataac agtcatgctt tccctcaact attagtgtag ctgatgaatg 3720
ctttcagaaa tgtggctgtt tttttgctgc gtattttatg tctttcatca tctcaaaatt 3780
tattgtgttt taacactgct tggaaggcct agagtcttcc gaaaatagta ggttgagaaa 3840
acatgagtaa gcaaatttct tcctgctggc aattaatcgt tttatatgag gtgaagccac 3900
acaatctaac atggtcttag acataaaaat attatttttg acatagagca gacagaaatc 3960
ttgggtctag ttgctatctc ccttcctaat aagatgccct ctatcctcat tgctatatcc 4020
caaaacagat tcacactaga atgaataaat ctctccttgc agccacattt atattaataa 4080
gtagtgtaac accaaaacat ttgtgcttct gcaaagtctg ttgaagtgga cgattaagta 4140
ataaattgtg tgagtttgac tgaagagaaa gtaataaatt gggtgagctt ctcacatttt 4200
tcaggataat gtcaattcat ctgattgaac tttttgaagg acatctcctt ttctaaaagt 4260
ttttgttcac ttcctgaaaa tggttgcttt cccaaattta ttctcctgta ttttccttgt 4320
gctttttttt tttaccagga tgactctggc cttccctttt gcaaatagtg acttcattac 4380
cttttaacaa aagaccaaat agcctttgta tgcttgaaat cgaagcagaa tttacggctc 4440
aatttctcta ggagatgtgt agcagtaaca taaaacgtgg ttaaagcaac ttgtaagcac 4500
tgccactgtc caatttgaca tgactctatt actatttaga gtgtcaaata gtgtttcttt 4560
tgactaccaa tttttcaatt tcttttttat accttttttc tttgactgtg acttatagta 4620
tattttatgt agtttcagaa tatatatttt agtttttaaa gaattgatgt ctgaaagtac 4680
agccaattag atggtttcac cctcgtactt taaatcttat aagttgtctt aggttaaagg 4740
aataatatgc atgattacat tgtttggatt gatattcaga agatcaaatg tgccggacaa 4800
aagcttgcag aagatataat tctgtcattt tgttgtttag ataaactacc cttgtcaaat 4860
acctaaaact aattcatact tttcaggatc cttcttgatt ggactactct gatcttcttc 4920
catatattga tatcctattc tttttgcttt taggttaacg gggagatata taaccataga 4980
gacttacggg aaaagctgaa gtcccaccag tttcgaactg gcagtgactg tgaagttatt 5040
gcacatcttg taagtacttt cagaatatat ttagataact atacatgact tctcagtttc 5100
tgtgcgcaga atgatttcaa aataccatga ctcaaatttt tcatcaaaag cgaaaaaggt 5160
ggattgtagt aggagaatga gattttttgt cataatttct ttcttatggt acttgaactc 5220
cgcaatctac ctatcgcagt atgaagacta tggagaagac ttcgttcaca tgttggatgg 5280
aatgttctcc tttgtgcttc ttgacacccg tgataaaagt ttcattgctg ctcgggatgc 5340
aattggcatt acaccccttt atatgggatg gggtcttgat ggtatgtaat tgttccattt 5400
cataattttt ggtatattgt ccttttaata aatagcccgt gagctatatg aagttccttt 5460
gttgtttcct tgttttttat ttgactgatg tatctttgtt gttttgccta ctattagggg 5520
tgtcaatggt tcggttcggc cggttatttt ataaaatttg taccatacca atttttaggt 5580
tattctatta tgtataacca aaattagact ttttgaaacc gtcccaatca tgtcggtttc 5640
tcttcggtat cggtacggtt cggttaattt tcggtatttt ttaatatcat gtaaaattca 5700
ccagtagaag tagaattgca ataacataca ttcttttata ggacttagca aaactctcta 5760
gatattttta ctgtttaaag ggtgatgaat taaaaaaaaa gaaaagaaag atggctagaa 5820
tatagatcca tcaactatta tacaacagcg taaaagaaat caaacaaaga caaagaaaat 5880
attaatcaca cgagttgaaa gatataccaa ggtaggactc aagaataaag tctatagaag 5940
attaaatatt caaaaagata aatctaaatt atatgaaagg aaacatattc aattcattgt 6000
agtttgctac tcataatcgc tagaatactt tgtgtcttgc taataaagat acttgaaata 6060
atttagttta agtagaagta gcataatagg ttttatgaat tagtattttg agtttaatta 6120
cttgttggct tgtaataatt ttcataattc caaggctcaa agaaaattta ttgcattatt 6180
atttttaaac ttactaaata aatatatttt ctacatgtaa aatttattcg gtatggttct 6240
gtattttttc ggtttatttt tataaaataa aaaacctacc ctaattatcg gtgcggttgt 6300
agatttatat aaaaacctac ggtttcttaa aaagaaagct aaaaatcggt tcggtgcggt 6360
acggttcggt cggtttagtc gattttcgaa tatccattga cacccctacc tactataatg 6420
ctggtctcat gctgttgtta caaacaaatg atgtataata gtgcatagta catgttttct 6480
tgccgctgta gatcaacaat gtaccaagaa catctgtctt agctagttca acctaatctt 6540
cagttatcta tcattccagt gaataacaca tatagagtta ggccgacatt acctaaggcc 6600
aactatacaa cattgtgagg tgcgactaag gaactctgaa cactgatcgg gagaaaaagc 6660
aacttatccg tgtttgtttt gaagtactat gaggaggcag caaccttgct ggatttatca 6720
cttgacagta aatgtcaatg gttgaggaac atttttaatg accggaggag aaacagagaa 6780
gagaagaaat aattgatttt aaaaaagcag aatagaaaat tttgtgaagt cgtatgttaa 6840
tttcaatgaa gaggatcaac aggtgcaata tttgacttgg cacctagcat gtagaagttt 6900
ctggttgggg ttgagcatgg taggcactgt tgcttttaca actgtcatta gagtcggaaa 6960
catattgtat tcttgaggga gcgttgctat ggataaaaaa ggcctatcta ctacttgata 7020
tttgggcatt cagaactagt tgtctaggag gaagaaaagg atttaaaatg gtgagtattg 7080
taggctttct ttgccgtcca ggatgcctgg gctagttccc tgaattggga tactggtgga 7140
ggatttcctg aaattgatca tggccctgat atgttagggg ataagaacat tgaaggatcg 7200
aaggaaggtg ccggcgctaa gttttgctag aaatgaacag ttccattatg gcttttgtca 7260
aagatgggaa tggaattggc cccaacttct aggcatctcc atccatcgtc aggtgaacta 7320
aagtaaggtt acactggtag aatttttcat tatttcatat taagtaaggt gaagccgtca 7380
acaatcattg cgtgagaagg aaccaggaac aaaatctaga agatgtctac taaagtgcta 7440
atagataatc tgcaaaattt agcacggtaa ggctagtgta gatctggttc aaattttaaa 7500
ttggtccaaa atgcgttaat aaatgtcggg cctcgttttg gtcccccgaa aggttgctaa 7560
ataaagactt gacaaagaaa caggtgaagg agaagcccaa acatttgaac aaactacacc 7620
tagcctaaac aaagtgattg tttgcgagga ggaaaagcga gcatgcggag agagcctcaa 7680
acacaaaaaa tggcctagtc ttgtaatgag tctagccaaa tcatgtaagc tccgactttc 7740
aagagagatc aacagaaaac ttcgtcaatc cggataaatc aagaaaatta tgataatgga 7800
aaggaggctg gacattctac tgccgacgca acccctggtt ttggagtcag tgagacaagt 7860
ggaactaaag aagaatatgg atcaaaagag tttatccagt tgacacctga tagactcact 7920
acatcagttt tcagttcaag aacatttcaa ttcttgctga taaagatagt tgccgataaa 7980
gataggcaaa ctgttaccta ccaatcaaag gaggttgttt caattgagtt agaggtatca 8040
aaggaggagc cagtatatat agagcaagtt ttgatgaagg gagttgcctc tatttgagac 8100
caccaggtac tcaagtgctt cccgtagttc ctaggcattt tatgatggat atgatagaga 8160
aatgtcttat tgacaagaat atactagggc tacagggata atagggttgg gaaggtgagg 8220
acttctttgg ttgctgtcag gaacttggaa acaaagggag aaaggggttg gaatcaaagg 8280
ttttgatttc gtttttgatt tataaggaaa taatgggtag tggatggagt cgaggatagg 8340
aaagaggttt accagttgca ccattgaata tgtaattaga gacactatcc tcttgttagg 8400
gaacacgact atcacaagca cgtaagggtg agtgaaagct ccgacatgga cgtggtttat 8460
tcgggtatgt tggcacgaag ccttcgtgtc taggtttggt gttgggttaa gtgcaactcc 8520
ttaggaaggc aaataaactt gggacagatt tgatagttgt cggaagttgc acaatgatga 8580
tattttcttt atccaaacgc tacgaatgat tgcacatata gtaactagag ttatttctca 8640
tttgagcttt tatatgtaag aatcaaataa aatgattgat gaaatgcatg aaagctcaaa 8700
tgagaaataa ctagttgtga gaatgcacgg gggaaaaata tattatattg attaaagtgt 8760
tgtacaaccc tatttagata cagtaattac ataataatag gtatctactt cccgatgtgg 8820
gacactagac atgactaact acttaacaat ccccctcaag ccggtgcata taaatcatat 8880
gtatcgagct tgttacagat gtaactaata cgagaaccag taagagactt agtgaaaata 8940
tctgctagct aatcattcga ctttacaaac tttgtaacaa tatctcttga gagtattttt 9000
tctctaacaa agtgacagtc gatctcaatg tgtttagtcc tttcatagaa caccggattt 9060
gacgcaatat gaagagcagc ttgattatca cacaccagtt ccatcttgct gatttctccg 9120
aacttcaact ccttgagcaa ctgcttgatc caaactagaa tcacacgttg ccacaaccat 9180
ggcccgatat tcggcttcgg cgctagatcg agcaactaca ttctgtttct tgctcttcca 9240
agagactaaa ttacctccta ctagaacaca atatccagac gtagaacgtc aatcagcagg 9300
taatcctgcc caatcagcat ctgtgtaccc aacaatctgc tcgtggcctc gatcctcaaa 9360
tcgtaaccct ttgcctggag ctgactttat ataccgaaga atgcgaacaa ctgcatccca 9420
gtgactatca cagggagaaa ccataaacta acttgcaaca ctcaccggaa aagaaatatt 9480
aggtctagtc actgtgtggt aatttaattt gccaaccaac ctcctctatc tcgtaggatc 9540
tctaagatgc tccccctgtc catgcagaag cttagcattc ggatccatag aagtgtcaac 9600
tggtctacaa cccatcattc cagtctcctc aagaatgtct aaggcatatt tccgctgtga 9660
aataacaata cctgagctag actgagcgac ctcaatacct agaaaatact tcaatctgcc 9720
cagatcctta gtctggaagt gctgaaagag atgctgcttc agattagtaa taccatcttg 9780
atcattgcca ataataataa tatcatcaac ataccactca actttggcat ttctatctca 9840
atttggctgg aatatcctgg acaatgccca ataacactgc gtctcttctc agtagctgga 9900
acaatggagg ggcaaatatt aaacagaaga aatggtggaa agtagttcct gcttgtatct 9960
ggtgggtcat gtggaaggaa aagaatgcag agtttttgaa gataggccta gctcttagcc 10020
tcttaccaga aaattaagct taattgtttt ttgttatttc atttttggtg caaagagagc 10080
tatagggagg atgtacattc tttactagat ttgctagaag aaatgtaatc gattagaata 10140
ggcaaggcag ttgcagatct tttggggtct gattgtggct tacactatct cctgtaaata 10200
tttttaatct tttatataat ttgttaccat tctaaaaaaa aacataaacc accagataaa 10260
tatacagatt aggagcagaa tgtcgataaa gcacagagtg atccgcttca ctacgagtca 10320
tgccgaactc ctgaataatt atgctgaact taccaaacca agctcgaggg aactgtttca 10380
aaccatatag tgacctgcgc aatcggcata caagaccact agacttccct taagcaacaa 10440
aaccaggtga ttgctccata taaatttctt cctcaagatc accgtggagg aaagcattct 10500
tagtgtctaa ctgataaaga ggtcattgac gtacaacaac catgaacaag aagagacaac 10560
cgatgctact ttagccacgg gagagagtgt cactataatc aagcccaaaa atctgagtgt 10620
atccttttgc aacaagacga gccttaagac gatcaacttg gccatccggg ctgactttga 10680
ctgcataaac ccaacgataa ccaacaatag acttacttga aggaagagga acaagcttcc 10740
aagtgccact cacatgtaaa gcagacatct cctcaatcat agtttgtcgc catcctggat 10800
gagatagtgc ctcacctgta gactttggga tagaaacaat tgacaaagat gatataaaag 10860
cataatgagg tgatgacaga cgatgataac ttaaatcgac ataatgagga ttaagattaa 10920
gagtggatca tacacctttc cgaagtgcaa tcggtgtact aggaagagac aagtccgcag 10980
taggagcagg gtcaagtgta ggatgtgaat cagttgggcc tgatgctggg tgcggacgac 11040
gatgatatgt caagagtggt gttccggtgg cggggaatct aggaggagcc acactagact 11100
ccccaacggt tggaatgggt aagacttatg tggcggaagg tgaaggagga gctatagtaa 11160
gctctttaaa ggtcggtata ggtaagacct cagatatatc aaggtggtca gaagaggtaa 11220
agaaaggttt agactcaaaa aatgtgacgt cagatgacat aaagtaatta cgaaaatcaa 11280
gtgagtaaca acgatatccc ttttgaacac gagaataacc aaggaagaca tacttgagag 11340
tacgaggagc taacttatct ttcctagggg ctaaattatg aacgaaacaa gagctcccca 11400
aaacacgagg aggaacagag tataagggtg actgggaaac aatactgcat acggaatctg 11460
attctggatg ggagatgaag gcattcgatt aaccaaataa caagctgtga gaactgtatc 11520
gcctaaaaaa cgcaacggaa catgagattc aaagagaagt gtgcgagcag tttcaatgat 11580
gtgcctattc tttctctctg caaccccatt ttgctgaggg gtataaggac aagaggtctg 11640
atgaataatt ccttgagaag tcataaagtg ctaaaattga gaggataact attctaaggc 11700
attatcactg cgaaaagtgc gaatagaaac accaaattga tttttaattt catcacaaaa 11760
attctggaat atagaaaaca actcagaacg atctttcatt aagaaaatcc aagtacatct 11820
tgaatgatca tcaatgaaac tatcaaacta acgaaatccc aaggttgagt tgactctact 11880
atgaccctat atatcagaat gaactaaaga aaaaacagac tatgcatgac tctcaatact 11940
acgaggaaag gaggctcagg tatgtttccc gagctgacat gactcacact ctaatgtaga 12000
taaactaggc actatcttct gaagcttgga tgtcctaaac atctgtgaat taggtccgga 12060
ggatctgtaa ctagacatgc cttggaggaa ttgagtgggt taaggtagta aaggccttct 12120
aattcaagtc ctgttccaat cgtctgtccg tactgcggtc ctgcataata aaagaatcat 12180
taataaaata tataccacaa tggagggcac aagtcaaacg actaacagat gcaagattaa 12240
aaggacaacc agggacataa agaacggaat ctagagtgac agagggtagg ggattcgctt 12300
gtccaactcc ttttgcttca gtttgagacc cgttggctaa agtaacagtg gaaagagact 12360
gtgaatacgc aatatttgac aaaagtgatt tattaccaga gatatgatca gaagcggttg 12420
agtccacaac ccattgtcca agagtactag actgggaaac acaagcaaaa gaattaccag 12480
caacaaaagc atcagtctga gcaacagagg ctacttgtgg agatgtctgc ttacttgctc 12540
gatactgaag gaacttatta tactcccctt ccgataaaga aaatacccgg ttacctatag 12600
tctcggtctg agcaacataa gcatttttgg gtggacgacc atgtaaagaa tagcacacgt 12660
cacgagtgtg tccaagttta tgacaataag agcacttggg tctagatctt ccaaaacgac 12720
ctcctcctcg tctattctcc atagtttgag atgcccgatt gttcacttac tgggatacga 12780
gaacagatga gtcaggtctc tgtgatgagc tcactgggtg acttggtgct gcagcaaggc 12840
gaagtaatcg agagaataat tcatcaactg tggggacagt cggactagcc aaaatctggt 12900
cacgtactga atcaaggtca ttagggagtc catcaagtgt aagaactaga aacatcttct 12960
gtcgttgctc ttgttgcttt ttaatactag cagaaactgg catcaacgtc tcaaattctt 13020
ccatgattgc ctgtacttgt cccctagtaa gtagacatat ccaattcctg tttcttcaag 13080
cttgtcattc gcgatattac atcatagaaa cgagatatgt cattagtgta taaattacga 13140
gccttttccc aaactaaata acatgtctgg aatggacgga acaagggcat caacttggaa 13200
tcaatagatc gccacaggat actacataac tgagcatcga ccttcttcaa aagtgttttg 13260
gccttttcat caccttcgct agcctttttt gttaaatgat cttgaactcc ttgaccttta 13320
caacacaact cgacagacga agcccaagct aagtagtttg aacttctcat taaaggttct 13380
gaggtaatca taatagcaga acttccagaa ctcgtgtttt tagacccaaa tacatccact 13440
cccaaagaca ttattggatt gaaaagagat ctagcaaatt agcaccaaat aaaacaaaga 13500
atcaactgtg gttgcccaaa aactgccgga aaactactgt agttgccgga aaattttcaa 13560
agtgctcgga atcaaaaaat aaaaatatgg gcaggctcgg aatggtagag cgatcagact 13620
aacctaaagg agtttttctg aaaaaattga cggaacgggc tccatgcgct ggtgcgtgga 13680
gtagatctcg ccggagaaga ctgtctccga tcggcgcgtg gcggtgcgtg aggcggctta 13740
tgacggaggt gttcgctggg gtttggtcgc cggaagttgg gggaccttgt ggtggtgttg 13800
gtttttgcac aacaccgatg gaattggttt tgacgaaaaa tagccctaaa aggtcaccgg 13860
aattgaagca cggcgacggc tggttttttt tttccccgga tgttttctca ctgccgctct 13920
gataccatgt gagaatgcac gggagaaaaa tatattatat tgattaaagt gttgtacaac 13980
cctatttata tacagtaatt acataataat aggtatctac ttcccgatgt gggacactaa 14040
acatgactaa ctacttaaca ctagtaacct catggttctc atcctacttc attgaccact 14100
tggccacatc cttgggtgcc tgcactgtag ttgtagctgt aatgaagata gttttttttc 14160
ttttctggat actcttatat tgtgacaatt tttctctatt tgctgttaat ggatgaggtt 14220
atgtgactct ccggtggtga gaatgttaat gaacatcaag ctcacatgtc atttcatttt 14280
ctgattgatc ttgctcttca ctctttacga gtttaatgtg ttagtccttc caagttctgt 14340
ccatctatct ttctggtgtg cttaacccat gtatattgga ccacactttg tattcatgct 14400
gattgcaaca aagcctgact gatcctgaac tcctccttta tgccttaggt agaaattttg 14460
tgtgtttagt gttacatgtt tcgcttcaaa ggaaaaaggg agaaacttaa ggtagaatcc 14520
tgctccttga cctttggatc cttgcacatt gaagtggaca gacatgtttg ctttgctttg 14580
atgagttgtt ttagtagtcg gtctaaacaa ctgctgatgt attctgcttt attgtaagtg 14640
tctataattt ttatctttcc acaatttgct ttcagctctt tctgcatgta gagaaaataa 14700
cagaagcttc tatttgatca tgtaatattg actgcgcaag ttcagtttta aacaactttg 14760
gctatgcata tgcaagcaat gacctgaaat aaatattatt ttttatgcag ggtctgtatg 14820
gttttcttca gagatgaaag ccttaagtga tgactgtgaa cgatttgtta gtttccttcc 14880
cggtcacatt tattcaagca aaaatggtat gcccatacga agctcatttc tgagcttgtg 14940
cttcatttat gttgatagtt gatcatgttt cctatacgat cagcgtactt ttgctattta 15000
tttcagattt actttaggac cctgtcggaa acagtcttga aacccttatt gatggtgtaa 15060
ttgttatctc ttgatcaacc aaaggagtaa ttacatgtaa ttattacaaa tttgcaataa 15120
aataactttt aggataacca gtggagctta atgaatcttt taggatattt gggatgctag 15180
agtgatgtac ttttgataat gtcggttaat ataaaatcct ctgactttgt actgatgtac 15240
tggctgagca gattttatat tactttctag tccctctgca aataaaaaaa ttgtgagact 15300
gcaatataac accatttttt aatgcattta caatgtttaa acttgatgta gcagtgaaga 15360
acatgtcgac cttaaacaga ggttctattt gtcctagata aaggaacatc agcgtaatga 15420
aacattacag gagagaagct ctattgagtt tttgttaatc tagagggagc ccgtaaatag 15480
aacaccaatt catctgttat atgacttctt tttcattttg gaacgtttta aaagtttgac 15540
aacatttttt gatctccgta cactaaactt tttgatgtat taaactaact atttccacta 15600
ctttaatatt ttcttcctta aacaactcat gttagcatct taaactgtgt gtgttcaata 15660
ccctcataaa aaattaaacg agtatttttt tatatttgct tatgcccttg gagattggga 15720
tggggaagtt catcatttga ggtgaaaggt aaattctttt ggatggacaa aaagtgaacc 15780
tcgtctttgg catggaggga ctatttaaat cgtcatcatt accaaagtca tcctttcaaa 15840
tgtttacttt caagaactat tcatgctatt tctcaagtat attatgggtg caggaggact 15900
cagaagatgg tacaacccac catggtactc agaaaccatt ccttctactc catatgatca 15960
ccttgtctta cggaaagctt ttgagaaggt cattctattt tggatgtcgc ttgtttctga 16020
gttgtacata atatttaggt gccactttgt tatattttga acttcatcaa aagcctttgg 16080
ttttttgagg acacatatcc ccttgactga ctgggcgctg aatttccttg atgtattgtg 16140
cttattttga ttgtcctctt ctttgtaata ctcttgccta taaataaacc ttctttatca 16200
aaagaaaatg ataaaatgtt aataacactt tgtgatatat gttgaagggg agtcttggca 16260
taactggtaa agttgctgcc atgtgaccag gaggtcacgg gttcaagccg tggaaacagc 16320
ctcttgcaga aatgtagggt aaggctgcat acaatcgacc cttgtggtcc gacccttccc 16380
cggaccccgc gcataacggg agcttagtgc accgggctgt cctttttttt tgcatagact 16440
ttacatgata ttttatttca atctttatca attctttttt accttcacat gcaaattata 16500
ctaccgtatg tgaaatatgc aatagttttt cccgcaattt tcatctttca tgtacctctc 16560
tctactatcc ttgtattact actgatttgg tgcattagat gttaatttta ttcccaacct 16620
ttactttatg caggctgtag tcaagcgact tatgacggat gtaccatttg gtgtgcttct 16680
ctcaggtgga ctagattctt cacttgttgc tgcagtggcc aaccgttatt tggctgatac 16740
tgaagctgcg cgacaatggg gatcacagtt gcataccttt tgcgtaggct tgaaggtggt 16800
gatgctccct tttgtaaact tgctctgatt agggaattgc tagctgtatc atcccaattc 16860
acatggaagt tctagaccat tattgctgtt tcttttccat atatagggat tttctgaaaa 16920
tagtaaaaag gaaatggaat ttctcattat taaagatacc aaatgccttt ttgggaaatg 16980
atcttacagt tgaatctgaa atttcctttt gacattgtga ttaattctga tatgaatttg 17040
gtccatcctg gctttccttc tcttgattgt agattttttt tctctttttg caattatatg 17100
cctttgtatt tcatatctct tattatgcta ttattttatt atgcattttt atggtactaa 17160
tatatcggct cctgttgctt tttttgagcc gagattttgt ttatttctgc tatcacgtcc 17220
tcctgaaaaa gtgctactat cttgtattca tgcgggagtt tgaaatttgc aagtgtataa 17280
ctttacaaaa tgattcgaac tgatcacagt tcattgctaa taaactttta taaaaagcag 17340
gtattgttgt gatataatgt gacacttaaa taaatttaaa cctacatcct ttcgaaatct 17400
ctaatagctt cactaatatt tcttgtgcta aattcttcca acacctctta tcgatgccaa 17460
atttggcaga ccttagtgca attgaaagtg tttagtgttc tttatgcagt aggtatgtta 17520
ttgtccaaaa tgacagaaat aaagttagag atttgttgaa aagccacttg gtgtgattga 17580
gcagttcgga gatgagtgat ttatccatct ttttatactt ttagtgtttt gttttctttt 17640
atattgtttt ctcttttgct tttgtcattg taattaaggt tgtcttttgt ttgactttag 17700
ggttctcctg atctgaaagc tgccagagag gtggcagact accttggaac tcgtcaccat 17760
gagtttcact ttactgtgca ggtaactttc ttcataaaga catcctatgc tgattttcct 17820
tataggagta taactgctta agtcaaattg gctatctact tctttctatt gctacatcag 17880
gaaggaattg atgcactaga tgaagtcatt tatcatgttg aaacatatga tgtgaccact 17940
atcagagcca gtacaccaat gtttctcatg tctcggaaga taaagtcctt gggtgtgaaa 18000
atggttctat ctggtgaagg ttctgatgaa atttttggcg gttatttata tttccacaag 18060
gcacccaaca antttacaca aaaatgacac aagttatgca gaaaaaatag tcctcaatgt 18120
catgggagac cagggttcaa atctgaggtt aaaaaaaata ctaggtgaac tcttctaatc 18180
cgtttaagcc ttggtagata gagttaacta gtatatgagc tagttagagc tagcaggtac 18240
ctggtagatt agtgcaggtg cacacgagct gactcaaaac caccattatt aaaaaataag 18300
aaagttttca atgttgttcc accgtgaatt catgatgtgc cgactttcct gtgtttatct 18360
ctggatagat gaagagtgtt tttggatctt gattaattct gtttggcccc tttgaggttc 18420
tgcaagctat ttctagactc aaacttgctg cgatctatgt taggtctaga gaatatctgc 18480
caaataaaac aatggaaggt acactcaagt attttatctc catattacaa gtcagtgtta 18540
gttgaatggg tctgatatat gggacagaaa atggcttcat acccataacc tcattaaaat 18600
gttgctgatg tgctgatctc atctgtacat ctagtataat cctaattaca aattaatgca 18660
tctctttgcg gtgactatag tatgctgaag aacttttctt gaaacattgc agatcagacc 18720
tgatctcgga agaatagaaa agtgggttct acgcaatgct tttgacgatg atcagaatcc 18780
ttatctgcca aaggtttgtt tgttttcttg tatcaggtcc aaaatcaaaa tctttgttta 18840
ttcatttgca taaacaggca gtagactccc aatacattgt gtacacttat gcacaaatgt 18900
atgagatgtg tattatatgt gtcacatata agggtatttc cacatggtgt ttattgatct 18960
agagaaagcg tacgacaggg ttcctaggga ggtcctatgg agatgcctag aggttaaagg 19020
ggtcccggtt gactgcatta gggtgattaa agacatgtat gatggagcta agactcgaat 19080
taggacagta ggaggcaact ccgaccattt tccggttgtt acggggttgc atcaagggtc 19140
tgcgttcaac cctttcctat ttgccttggt gatggatgct ataaccgcat catattcaag 19200
gggaggtgcc atggtgcatg ttatttgctg atgacatagt cctaattgac gagacacgac 19260
gcggcgtcaa cgagaggcta gaggtttgga gacatgccct tgagtctaaa ggtttcaggc 19320
tgagcaggac gaagacggaa tatctcgagt gcaannnnnn nnnnnnnnnn nnnnnnnnnn 19380
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn 19440
nnnnnnnnnn nnnnnnnnnn nnatcgtata tgggtagggt ggatgaagtg gaggttagca 19500
acgggagtct tgtgtgacaa gaaagtgtca ccgttactaa ntctcgagtg caactttggg 19560
gccgagccga tggaagcatg agtggaagtg aggctcgact cacaagtcat ccctaagagg 19620
ggtagtttca agtaccttgg gtcagttatt caggggaccg gggagatcga cgaggatgtc 19680
acacatcgta taggggtggg gtggatgaag tggnnnnnnn nnnnnnnnnn nnnnnnnnnn 19740
nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnnnnn nnnnnnatag gggtagggtg 19800
gatgaagtgg aggttagcaa cgggagtctt gtgtgacaag aaagtgtcac cgttactaaa 19860
aggtaagttt tacagagcag tggttaggcc tgctatgttg catgggaccg agtgttggcc 19920
ggttaaaaat tcacacatct agaagatgaa agtagcagag atgaggatgc tgaggtggat 19980
gtgcgggcat acaaggatgg acaagattag gaatgaagtt attcgagaga aggtnttgag 20040
ccgagggtct cctggaaaca gcctctctgc ccctcggggt aggggtaagg tctgcgtaca 20100
tactaccctc cccagacccc attggtggga ttatactggg atgttgttgt tgttgttgtt 20160
gttgttgtac atataagggt attatatgta cacactcctc ccacctttca cttatatatt 20220
cacgtaaaac acatgcatgt gtaaaaccta tatatgtgtg tgtgatatat ataataaaat 20280
gatgtttgaa aagacattat gatcaagtag ttgatcttca tctgctttcc gtttttagca 20340
tatcttgtat aggcagaagg aacagttcag tgatggagtt ggctacagtt ggattgatgg 20400
tttgaaggat cacgcaagca gactggtgag ctttttagtt gttttctctt ccctcctctt 20460
ccattcttta tcccttcctt tgtggttttg ttgaacacat ctcgtatatt tggtccaggt 20520
ttctgattct atgttagcga atgcaagttt cgtttacccg cataacacac ccacgacaaa 20580
ggaaggatac tactatagaa ctatttttga gcgatatttc cccaaggtgg gtcttagata 20640
gtcttctttt ccttgttatt catgttcaat ttttgatatt ttgtcccaaa caatgttttc 20700
tttttcttcc ttctgttaac aaggtcgctt ctatcttcat atttgttgat aaaggtaatc 20760
acattgataa ctattatttg gagcagaatg ctgcgaggga aacagttcca ggtggtccaa 20820
gtgtggcatg cagcactgca aaagcagtag aatgggacgc agcttggtcc aagaatctag 20880
atccatctgg acgagctgca ctcggtgttc atgcagctgc ttatgaggat gcatcagagg 20940
ttaaaaccac agtcgcaaca gatacccctc agaaacttga agttgataaa gctgcagtag 21000
ctgtttga 21008
<210> 10
<211> 581
<212> PRT
<213> Nicotiana tabacum
<400> 10
Met Cys Gly Ile Leu Ala Val Phe Gly Cys Ile Asp Asn Ser Gln Ala
1 5 10 15
Lys Arg Ser Arg Ile Ile Glu Leu Ser Arg Arg Leu Arg His Arg Gly
20 25 30
Pro Asp Trp Ser Gly Leu His Ser His Asp Asp Cys Tyr Leu Ala His
35 40 45
Gln Arg Leu Ala Ile Val Asp Pro Thr Ser Gly Asp Gln Pro Leu Tyr
50 55 60
Asn Glu Asp Lys Thr Ile Val Val Ala Val Asn Gly Glu Ile Tyr Asn
65 70 75 80
His Arg Asp Leu Arg Glu Lys Leu Lys Ser His Gln Phe Arg Thr Gly
85 90 95
Ser Asp Cys Glu Val Ile Ala His Leu Tyr Glu Asp Tyr Gly Glu Asp
100 105 110
Phe Val His Met Leu Asp Gly Met Phe Ser Phe Val Leu Leu Asp Thr
115 120 125
Arg Asp Lys Ser Phe Ile Ala Ala Arg Asp Ala Ile Gly Ile Thr Pro
130 135 140
Leu Tyr Met Gly Trp Gly Leu Asp Gly Ser Val Trp Phe Ser Ser Glu
145 150 155 160
Met Lys Ala Leu Ser Asp Asp Cys Glu Arg Phe Val Ser Phe Leu Pro
165 170 175
Gly His Ile Tyr Ser Ser Lys Asn Gly Gly Leu Arg Arg Trp Tyr Asn
180 185 190
Pro Pro Trp Tyr Ser Glu Thr Ile Pro Ser Thr Pro Tyr Asp His Leu
195 200 205
Val Leu Arg Lys Ala Phe Glu Lys Ala Val Val Lys Arg Leu Met Thr
210 215 220
Asp Val Pro Phe Gly Val Leu Leu Ser Gly Gly Leu Asp Ser Ser Leu
225 230 235 240
Val Ala Ala Val Ala Asn Arg Tyr Leu Ala Asp Thr Glu Ala Ala Arg
245 250 255
Gln Trp Gly Ser Gln Leu His Thr Phe Cys Val Gly Leu Lys Gly Ser
260 265 270
Pro Asp Leu Lys Ala Ala Arg Glu Val Ala Asp Tyr Leu Gly Thr Arg
275 280 285
His His Glu Phe His Phe Thr Val Gln Glu Gly Ile Asp Ala Leu Asp
290 295 300
Glu Val Ile Tyr His Val Glu Thr Tyr Asp Val Thr Thr Ile Arg Ala
305 310 315 320
Ser Thr Pro Met Phe Leu Met Ser Arg Lys Ile Lys Ser Leu Gly Val
325 330 335
Lys Met Val Leu Ser Gly Glu Gly Ser Asp Glu Ile Phe Gly Gly Tyr
340 345 350
Leu Tyr Phe His Lys Ala Pro Asn Lys Glu Glu Phe His Gln Glu Thr
355 360 365
Cys Arg Lys Ile Lys Ala Leu His Leu Tyr Asp Cys Leu Arg Ala Asn
370 375 380
Lys Ser Thr Ser Ala Trp Gly Val Glu Ala Arg Val Pro Phe Leu Asp
385 390 395 400
Lys Glu Phe Val Asn Val Ala Met Asn Met Asp Pro Glu Ser Lys Met
405 410 415
Ile Arg Pro Asp Leu Gly Arg Ile Glu Lys Trp Val Leu Arg Asn Ala
420 425 430
Phe Asp Asp Asp Gln Asn Pro Tyr Leu Pro Lys His Ile Leu Tyr Arg
435 440 445
Gln Lys Glu Gln Phe Ser Asp Gly Val Gly Tyr Ser Trp Ile Asp Gly
450 455 460
Leu Lys Asp His Ala Ser Arg Leu Val Ser Asp Ser Met Leu Ala Asn
465 470 475 480
Ala Ser Phe Val Tyr Pro His Asn Thr Pro Thr Thr Lys Glu Gly Tyr
485 490 495
Tyr Tyr Arg Thr Ile Phe Glu Arg Tyr Phe Pro Lys Asn Ala Ala Arg
500 505 510
Glu Thr Val Pro Gly Gly Pro Ser Val Ala Cys Ser Thr Ala Lys Ala
515 520 525
Val Glu Trp Asp Ala Ala Trp Ser Lys Asn Leu Asp Pro Ser Gly Arg
530 535 540
Ala Ala Leu Gly Val His Ala Ala Ala Tyr Glu Asp Ala Ser Glu Val
545 550 555 560
Lys Thr Thr Val Ala Thr Asp Thr Pro Gln Lys Leu Glu Val Asp Lys
565 570 575
Ala Ala Val Ala Val
580
<210> 11
<211> 17971
<212> ДНК
<213> Nicotiana tabacum
<220>
<221> misc_feature
<222> (1389)..(1389)
<223> n представляет собой a, c, g или t
<400> 11
atgtgtggaa tactagcagt tttcggttgc attgataatt ctcaggccaa gcgttcccga 60
atcatcgaac tttctagaag gtagaatttt tttaaaaaaa aaattgcatt ccaatttttc 120
tacagcattc ttattctttg gggtgacaat ggtatgcatg tgttttggat ttgctattcg 180
tggacttgtt cgctttctcg tggtcttcga tttgtttttg ttaatttata atcagtgttt 240
ttatatggat ttcttttttg aatctctcaa atatatgaca tttgatttcc acgagaaaat 300
ttggcatttt tatggttctt atgatatgcg gttaaccctt cgccttgcat cgcttcagcg 360
ctttaaaaga cgaaatgatg gctattagta atactgctag agtggttatt gaattacatt 420
aaataacaga ttgctagtag atcaaattca gaatgacagt gaaatatctc tggaaacttg 480
aatccccgtt caattcgctg gtaaaacctg gcaaaaaaat agctttatct caaaggagag 540
tatgtcactg tgaaattggt ggtatgtgtt gtttaagctt taaggcatga tatgctgctc 600
ttaggtgatg tcaggatgat aataccatcc ttttcacact agactcagct ttgaagcact 660
ttcatctcag ttaatcggtg gaggaaaatc tagtttgctt tgggcataat ataccaaatg 720
tcttcctttt gttttctagt gttgtttctc aagaattgaa aaaccaacct acaatatagt 780
gatataatta actagttagc tcttcctgtt tttctgaaaa tttccaataa ttgagaagcg 840
tttacaggtc aattagtaaa gggagaacgg aacatttttt tcctctaaag aacttctctt 900
tttccataca gaatgacttt ctatacaagg cctaagtctg ctttagtgtc tgcgattaag 960
catcttctgg gaaacttcac acttgaaaga agtagaacta ggagggggca ggtctagcaa 1020
gataattgct tcagtcattt ctcataaaaa aacattgttt cattcattat gtgcattcat 1080
atagagttgg ttaagctgct aagctgagct aatagtatga tagctatgaa tctcgacaaa 1140
gtaggttgct tcttagggct ttattctttg gctccgtaaa gaatttcttt tggaagggct 1200
tcagagtggg taggactgtt ctgatgataa gctgtaacac caacgaaaca agcctttgta 1260
gctgtggaag tccgaggggg tggtggtggt ggtggtggtg tgagagttag agtggtggag 1320
gagaggggag gagtgaagag aagtttatat tttaaagaat agggccttga gaacttttag 1380
tagtgagcng gggggggggg ggggcagagg ggaggagtga agagaagtat atattttaaa 1440
gaatagggcc ttgagaactt atagtagtaa gcatgcatta ttattattgt tgttattatt 1500
attattatta ttatttttat tattatgtaa gatgtaatta cttagctaat gaggtttttc 1560
caactatttc caaattgtga aatgtgttga aagcataatt aagtatataa aagcgtgttc 1620
tttctaatat ctttagcttt aagaggagat aatcacaaaa tttatcttga taatatagca 1680
gacaattttt cctagttcaa atctcacgtc acccttcata aaaaatttcc acgtgcttgg 1740
cttgcatgtt aggggacgtc ttgacataat taaacaaata taattgtccc tctctaacag 1800
cttaagcttt tagatgaggc ggtcaacaat tcaacaattt gcatttgttt gataagaagt 1860
tatattaaaa gaacacttaa aaggtgtcag tatgtgcaca aactcaaggt gtagactaac 1920
tagaagtcaa ttcaaatttt ctgcttgttc tctctgtttt acttcttttg cactatagaa 1980
attgcatttc cagcattccc gaaagctgct tcctttgcat atagacatca catttccagc 2040
gtgtctgata agcgttcttg cttattcctg tccctctact tggttgattt tcctctcact 2100
tagttcactg tcttcttgca cgcattatca aattgtgttc tcaattctct tgccttcttt 2160
tcatctgacc ccacaaaata cctcccaaaa agcctttagt ttctgctttt ccgagtctga 2220
aattatctgc ttattactgc ctttcaagta ttctgatacg ttgaccattg ttttccccag 2280
tagtaatgca ttgttctctg tccaatctct attttttttt ttgtgaacca ctattctttg 2340
ttgcttctgt ttcgtcatcc tcagttcccc atcttctcaa tttgattttc ctcatttaaa 2400
aagcgtgtgc cccttttctt gtgctaccct ctcaaaaatc agttgctgtc atctcgtttg 2460
atctgtcatg aacccagttg atgccacatg tttagttctt tacctctgga caacttttac 2520
ttcctgagaa ccacaatttc ctttccttaa gaaattctta gaaattggtt atgtaaactg 2580
taaagggaaa atttcccagc aatcaccatt tggcctctac gtggcttgct tttcaactta 2640
ccgacacgtg ttcatcccct tttaacaaat gctgaccatt atcttctgct tgttggggca 2700
tgcttcttac ccatggtgtt aaccctgatc tttcgtatca catttttcaa ctcttaattc 2760
tcatcttata aagtgcatat ccttgcctct ttagttgatc ttgggttggg ctgttaatcc 2820
aaattccatg ggctttgctt tgaagtatga taagcattag ctttactaca gtagcatatt 2880
ctacgttgct acagacatga aattttcctg atttgtcttt gatgtgatgt ctggcaagtg 2940
tagattacgc cacagaggac ctgactggag tggattgcat agccatgatg actgttatct 3000
tgctcatcaa cggttggcaa tagtagaccc aacttctgga gatcagccac tttacaatga 3060
ggacaagacc attgttgtcg cggtagggaa atatttatta tatcatttct tagatagtta 3120
agtattttac cagtacatgg actttggaca atatttgtgc gttttgatct tctatagcac 3180
aaacttttct cgtaggataa atttcctcta ctttgagtga atcttaacag gaaaatcgga 3240
ataaacaatt tgtgatttcg aaatttttag ttcttacata tttgaaacat gtgcggtgaa 3300
gtaactaggc gcttgactag gtgaatacgg agaatttcct ctactgttat cgatattttc 3360
tcagttcatt tgaaagatct ctgggtcgtc tagcagtaca actatttagg caacttaatc 3420
aaatcccctt cccatgagtt tcactttctt gacttgtttt atattttgac attttgcaat 3480
gccgaaaata aattgttttt ccattttatg acctttaaaa tgttgaggtg aagggctagg 3540
cggtacttta atcgacttat cggcaacaat gtattagata cagagcacaa acttgtttgg 3600
gtgcctaaat tgatattttt gaaaagtcga tggagtaaat ggaaatatgg attagcaaaa 3660
atattgaaat agtagatcta aacaatggat gactgaattt agaggaggac cttttcttct 3720
ttccttgttg aagtggttat aagagaatta ttggaagaaa atgtaagctt tagggtagtg 3780
ttgcaccata acagtcatgc tttccctcaa ttattagtgt agctgatgaa tgcttttgga 3840
aaggtggctg attttttgct gcgtatttta tgtctttcat catctcaaaa tttgttgtgt 3900
tttaacagtg cttggaaggc ctagagtctt ctgaaaaaag taggctgaga aaacatgagt 3960
aagcaaattt cttcctgctg gcaattaatc tttgtatatg aggtgaagcc acataatcta 4020
acatggtatt agacatataa atattatttt tgacatagag cagacagaaa tcttgggtct 4080
agttgctatc tcccttccta ataagatgcc ctcattgcta tatcccaaaa cagattcacc 4140
ctagaacgaa taaatctctc cttgcagcca catatatatt aataagtagt gtaacactaa 4200
aacatttgtg cttctgcaaa gtctgttgaa gtgggcgatt aagtaataaa ttgtgtgagt 4260
ttgactgaag agaaagtaat aaattgtgtg agcttctcac attttcatga taatgtcaat 4320
tcatctgatt gaactttttg aaggacatct ccttttctaa aagtttttgt tcacttcctg 4380
aaaatggttg ctttcccaaa tgtattctcc tgtattttcc ttgtgcttta tgttgaccag 4440
gatgactctg gccttccctt tttcaaatgg tgacttcatt accttttaac aaaagatcaa 4500
atagcctttg tatgcttgaa atcgaagcag aatttagggc tcaatttctc taggagatgt 4560
gtagcagtaa cataacacgt ggttaaagca acttttaagc actgccactg tcaaatttga 4620
tatgacgcta ttactattta gagtgtcaaa tagtatttct tttgactacc aatttttcaa 4680
tatctttttt ataccttttt tctttgactg tgacttatag tatattttat gtagtttcag 4740
aatatatatt ttagttttta aagaattgat gtctgaaagt acagccaatt agatggtttc 4800
acccttgtac ttcaaatctt ataagttgtt taggttaaag gaataatatg catgattaca 4860
ttgtttggat tgaaattcag aagatcaaat gtgcaggcaa aaagcttgcg gaagatataa 4920
ttctgtcatt ttgttgttta gattaacttc ccctgtcaaa tagctaaaac taattcatac 4980
tttctcagga tccttcttga ttggactact ctgatcttct tccatatatt gatatatcat 5040
attccttttg cttttaggtt aacggggaga tatataacca tagagaatta cgggaaaagc 5100
tgaagtccca ccagtttcga actggcagtg actgtgaagt tattgcacat cttgtaagta 5160
ctttcagaat atattcaaat aaatatacat gacttctcag tttctgtgcg aagaatgatt 5220
tcaaaatacc atgactcaaa tttctcatca aaagagaaaa aggtggattg tagtaagaga 5280
atgagatttt ttgtcataat ttctttctta tggtatttga actccacgat ctacctatcg 5340
cagtatgaag actttggaga agacttcgtt cacatgttgg atggaatgtt ctcctttgtg 5400
cttcttaaca cccgtgataa aagtttcatt gctgctcggg atgcaattgg cattacgccc 5460
ctttatatgg gatggggtct tgatggtatg taatttttcc atttcataat ttttggtata 5520
ttgccctttt aataaatagc ctgtgagtta tgtgaagttc ctttgttgtt tccttgtttt 5580
ttatttgcct gatgcatctt tgttgttttg cctactataa tgctggtttc atgctgttgt 5640
tacaaacaaa tgatgtataa tagtgaacag tacatgtttt cttgccgctg tagatcaaca 5700
atgtaccaag aacatctgtc ttaagctagt tcaacctaat cttcagttat ctatcattcc 5760
agcgaataac acatgtagag ttaaccggac attacctaag gccagctata caacattgtg 5820
aggtgtgact aaggaactct gaacactgtt tggagaaatg gaaagcaaca tatccgtgtt 5880
tgttttgaag tactataagg aggcagcaac cttgctggat ttatcacttg acagcaaatg 5940
tcaatggttg aggaacattt tttatggctg gaggagaaac agagaagaga aaaaataatt 6000
gattttaaaa aacagaggag aaaattttgt gaagtcgtat gttaatttca atggctggag 6060
gtgcaatatt tgacttggca actagcgtgt agaagtttct ggttggggtt gagcatggaa 6120
gacactgttg cttttacaac tgtcattaga gtcggaaaca tattgtattc ttgagggagc 6180
gttgcaatgg ataaaaaagt cctacctact acttgatatt tgggcattca gaactggttg 6240
tctaaagctg agaaggaaga aaaggattta aaatggtgag tattgtaggc tttcttttct 6300
gtccagggtg cctggactag ttccctgaat tgggatactg gtggaggatt tcctgaaatt 6360
gatcatggcc aaagagatat gtcaggggat aagaacattg aaggatcaaa ggaaggtgct 6420
gcgctaagtt ttgctagaaa tgaacagttc cactatggct tttgtcaaag atgggaattg 6480
aattggcccc aacttctggg catctccatc catcgtcagg tgaacaaaag taaggttaca 6540
atggtagaat ttttcattat ttcaaacaag taaggtgaag tcgtcagcaa tcattgcctg 6600
agaaggaacc aggaacaaaa tctagaagat gtctgctaaa gtgctaatag ataatctgcc 6660
aaatttagca cggtaaggct agtgcagatc tggttcaaat tttaaattgg tccaaaatgc 6720
tttaataaat gtcggacctc gttttggtcc cccaaaaggt tgctatataa agacttgaca 6780
aagaaacagg tgaaggagaa gcccaaacaa ttgaacaaac tacacctagc ctaaagaaag 6840
tggttgatag tttggtcgga ggaggaagag cgagcctgcg gagagagcct caaacataaa 6900
aaatgaccta gtcttgtaat gagtctggcc aaatcaagta agctctgact ttcaagagag 6960
atcaacagaa aacttcgtca gtctagataa atcaagaaaa ttatgataat gaaaaggagg 7020
ctggacattc tactgccgac caaccctggt tttggtgtca gtgagacaag tggaactaaa 7080
gaagaatatg gatcaaaaga gtttatccgg ttgacacctg ataaactcac tacatcagtt 7140
ttcagttcaa aaacatttca attcttgctg ataaagacag gcaaactgtc acctaccaat 7200
caaaggaggt tgtttcaatt gagttagagg tatcaaagga ggcggcagta tatatggagc 7260
aagttttgat gaagggagtt gcttctattg gagaccacca ggtactcaag tgcttcccgt 7320
agttcctagg cattttataa ttgatatgac agagaaatgt cttattgaca agaatagact 7380
aggactacag ggataatagt gttgggaagg tgaggacttc tttggttgct gtcaggaact 7440
tggaaacaaa gggagaaagg ggttggaatc aaaggttttg atttcatttt tgatttatga 7500
ggaaataatg ggtagtggat ggagtcgagg atgggaaaga ggtttaccaa ttgcaccatt 7560
gaatatgtaa ttagagacac tatcctctta tttgggaaca cgactagggt gagggaacgc 7620
tccgacaggg atgtggttta ttcgggtatg ttggcacgaa gcttttgtgt ctaggtttgg 7680
tgttggctta agtgcaacac cttaggaaga caaacaaact taggacagat ttgatagttg 7740
tcggaaattg cacagtgatg atattttctt cttccaaacg ttgggaatga ttgcacatgt 7800
agtaactaga gttatttctc atttaagctt ttatatgtaa gaatcaaata aaataattga 7860
tgaaatgcat gaaagcaact gtcttacgcc tgaatgcttt taaaaggcat gctttacaaa 7920
gtttttggta catctatctg acatgacaca ctaacattca ctcttatatg tgtacatatt 7980
taactaagtc caagcctaac tatctaactg gagatatcaa agttcttagc tctttaaagc 8040
tttcttaact ttaggaagat cctgcagtgt tgttttttta taaccgtggt atccaggcta 8100
gcttgcacac atctcgacta attccaccgg gtgcttgcta tctcccacca acacagatta 8160
cctggtaact catgggaaga aatcacctat tgttttgagt ctgctggcat ttgaagtttt 8220
ggactactaa cctcatggtt ctcatcccac ttcattgacc acttggccac atccttgggt 8280
gcctgcactg tagctgtaac gaagatagtt tttttttccg ggatactctt atattgtgat 8340
aatttttttg tattcgctgt taatggatga ggttatgtga ctctccggtg gtgagagtgt 8400
taatgaacat caagctcata tgtcatttca ttttctgatt gatcttgctc ttcactcttt 8460
acgagtttaa tgtgttagtc cttcccagtt ttttttaaga gcgagaagcg caaaaaagcg 8520
acaagggctc gcctcgcttc aaaagcgaag cgcaaagcga agcgcacact ttattaaagt 8580
gaagcgcaat tcttaaaaaa cataatgtaa accttgcaaa gacacaatat aaaattataa 8640
ataatcaaaa agttcaaatt gtcaaaatca aagctactaa attactagaa tcaacctctt 8700
attcttctac tcttcttgtt cttcttcaag attgtcaaaa tcttgaattc ctctattatc 8760
ttcatattgc tcgtcatctt cctcctcccc ctcctcttcc tcttcatgat cactttcatc 8820
ttcatcaatt agggataggg atcgacttgt agtagccaca ctttttccct tcctaatcga 8880
gcttgaactt gaagtattcc cccttaaacc ataaggattc tccccaactc cactagcaac 8940
cgcaacatca ccccaagtga aattagaatc gccttcaaat acttcttcat cttcacaatt 9000
ttcggggact ccggttagcc actcattagc ctcatcaata ttgtccaaaa gaattggatc 9060
aattagattg cggtggttgt agcgacgcct caatgctcta ttgtatttta tgaacactag 9120
attatggagg cgcttcaagg ttagtcgatt cctcttcttt gtatgaatct gcaaacaaga 9180
tgtgtcaact aattagtagg agttgggaca attgagatat aatttacttt atctcaaaac 9240
acgaaataat tctttttatt tctcacgtgt tcaaaaacgc tccagttcct ttcacatccg 9300
gatgagctac aagttaaact tagaactctg atggcgaaag tctgtaaatt cggagtctct 9360
acaccatatt ggtcccacca ctcaactata gaagtgaaaa aaaaaaattc aagagttaat 9420
aagtataaaa aatacattaa agttagagct ataaaatata gaagcacttg gtcacctggc 9480
gatttcgtct ttctttgttt aatagcaagt cggagcttaa aaagtccctc agctgccttg 9540
taaatagcaa gctgatctac tatcttttct tgcatgtctt catctggggt caacttgata 9600
acaacctcat ggaatcctgt ccacacttct ctagccaatg aattattctc atgctgatca 9660
taaaagagtg acgggttcag aataagtcca gctgcatgca aaggtctatg aagttgctca 9720
ctccatcttg catcaatgat ctgaaagacc tttgcatatt tctgctcatc agtgaatgat 9780
gcttgaatag cctccttggc cctatccata gcttcataga ggtagcccat tggtggtttt 9840
tgctccccat ccaccaaacg gagtacttta accaaagggc caccaatttt aagagcatga 9900
aggacattat tccaaaaaga ataagaaaga ataatgcgtg caacatcttt ccctgcactt 9960
tcctttgcaa atttactctt gctccattcc tctgaagtga acaactttct caaattggat 10020
ttttgcaagt ggatactatg taaagtcaag aaagcagtgg cgaaccttgt cttgcccggt 10080
ttcaccaaat ttttttgtcc ggtgaatctt ctcatcatat tcaataacaa gggccgctga 10140
gaaatataag aatgtaccct aacgccctgg ccaaaaactg tagagaaggg tttttccttg 10200
aaaatgtccc cgaagattaa gttgatacaa tgagccgcac atggagtcca atagacattc 10260
ttgtacgctc cttccaccat gccacccgct ttcacatttt cactcgcatt atcagtgacc 10320
acttgaacaa ctttgcttgg gccaatcttt tcaatggtgt tctgaaacaa ggtgaacatt 10380
ttgatgtggt cagtggatga gtcgctagca tcaatggact caagaaacaa acttcccttg 10440
ggagaattca ccaacacgtt aataatcatt ttcccagttc ttgccgtcca cttatccatc 10500
ataatggagc agccatactt gttccacgca actttatgtt cctccacaat tttattagtc 10560
tcctccactt ccttatttag ataaggacct ctgatttcat gataagtggg aggcttcatt 10620
ccaggtccgt attgaccaac ggcctcaata aagtctccaa aagtgtcagt atagttgaca 10680
caattgaagg ggagcccagc atcatagacc catcgtgcaa aagctctaac tgcacgatcc 10740
ctcaaaatgt ccttagcaat tttttgtaca tcttttccac cgctctttcc ttccggcttc 10800
tttgggaaat agcaatctat aggaccttta gtcctactcg tacccgtcga tccatggctt 10860
gaagagattg catctcttcc ccgttttgtt ggaagtgaca attcttcaat atcatcatca 10920
ccatcatcat caagattggt caccaatggt tgatgactca tttgattttt ttgctccttc 10980
ttcttctcaa caaaattctt tatttcatcc ctcacctccg gtggacattt cggacaactt 11040
gtgacgtttc tatcgccacc aataagatgg aatttaaagc gataaattcc acccgttgta 11100
atcttgttac aaaacttgca tacaatattt gtattcttct cattaactct atcgccatat 11160
cgccaagccg ggtctttatc tttcgatctt tgagacattt tgaaatccct attaagatat 11220
aagaaaatac ataatcaaat aaactgaaaa tacatataat atatataata attaatttta 11280
taaaccgaaa tacacattaa aaaaatcaaa taaactaaaa tataacatat ataattttct 11340
aaattgaaat acatataaaa ttaatcaaat aaactgaaaa cataacatat atataataat 11400
taattttata tactgaaaaa tgcattaaga aaatcaaata aactgaaaaa tataacatat 11460
ataaataata attaatttta tatactgaaa taaaaatcaa ataaactaaa atataacata 11520
tatatataat aattaatttt ctaaaatgaa atacatataa aattaatcaa ataaactgaa 11580
aatataacat atataaataa taattaatat tatatactga aatatacatt aaaaaatcaa 11640
ataaactgaa aaatataaca tatataaata ataattaatt ttatatgctg aaatatacat 11700
taaaaaaatc aaataaactg aaaaatataa catatataaa taataattaa ttttataaac 11760
taaaatacat attaaaagta ataaataaga agaataaagg ggggaaaaaa acaaaatggg 11820
ttagagataa aaggaagaag aagacgctga agtctgaaca gtgcaaactg gactgtcaga 11880
gtctcagaga taaacgaaga agaagaagga agaaagaagg aaaaagaaag aaaaaaaagc 11940
agaatctgac gggaccaaga agaagaaaga aagaagaaag aaaaaaaaga gaagaagaaa 12000
attacctgga ctggtcagca atgtcaatga agttgaaccc taggctttgg tcgacctcga 12060
tcgaagaaga gagaagaaga aatggccatt ttttgtcaat ttagggtctg ttttgtataa 12120
tagaaaaaca gacccaaagt tttttttaaa aaaaaaaggg cctgcgcttt tttaacaaaa 12180
gcgatcgctt cgtcgcttcc tccgttgaag cgtgcgcttc cctcggtcga gtcgcctcag 12240
gcagctagaa gcgacactga gtcgcgtcgc gtcgcttcgc gctttaagcg cgaagcgatc 12300
gcttttctaa acactggtcc ttccaagttc tgtccatctt tctggtgtgc ttaacccatg 12360
tatattggac cacactttgt attcatgctg attgcaacaa agcctgactg atcctgaact 12420
cctccttttt gccttgggta gaattatttt tgtgttttta gtgttacata tttcaattta 12480
aaggaaaaag ggagaagctt aaggtagaat cctgctcctt gacctttgga tccttacaca 12540
ttgaagtgga cagacatgtt tgctttgctt tgctttgatg agttgtttta gtagtcagtc 12600
taagcaactg ctgatgtatt ctgctttatt gtaagtgtct ataattttta tctttccaca 12660
atttgctttc agctctttct gcatataaga gaaaataaca gaagcttcta ttcgatcatg 12720
taatattgac tgcacaagtg catatttaaa caactttggc tatgcgtatg caagcaataa 12780
taaatattac tttttatgca gggtctgtat ggttttcttc agagatgaaa gccttaagtg 12840
atgactgtga acgatttgtt agtttccttc ccggtcatat ttattcaagc aaaaatggta 12900
tgcccatacg aagctcattt ctgagcttgc gcttcattta tgatcagcgt acttttccta 12960
tttatttcag atttacttta ggaccctgtt ggaaacagtc ttgaaccctt attgatggtg 13020
taattattat ctcttgatca accaaaggag taattacatg taattattac aaatttgcaa 13080
caaaatcttt taggataatc agtggagctt catgaatctt ttaggatagt tgggatgcta 13140
gagtgatgta cttttgataa tgtcggttaa cataaatcct ctgactttgt actgatgtac 13200
tggctgagca gattttatat tactttctaa tccctctaca aataaaaaaa ttgtgagact 13260
gcaatataac accatttttt taatgcattt acaatgttta aacttgattt agcaacgtag 13320
aacatgtcga ccttaaacag aggttctatt tgtcctagat aaaggaagat cagcataatg 13380
aaacattaca ggagaggagc tctattgagt ttttgttaat ctagagggag cccgtaaata 13440
gtacaccaat tcatctctta tatgacttct ttttcatttt ggaacgtttt aaaagtttga 13500
ctacattttt tgatctccgt acactaaact ttttgatgta ttaaactaac tatttccact 13560
actttaatat tttcttcctt aaacaactcc tgttagcatc ttaaactctg tgtgttcaat 13620
accctcatat aaaattatac gagtattttt tttatatttg cttatgtcct tggagtttgg 13680
gatggggaag ctcatcattt gaggcgaaag gtaaattctt ttgaatggac aaaaagtgaa 13740
cctcggtctt tggcatggag ggactattta agtcgccatc attaccaaag tcatcctttc 13800
aagtgtttgc tttcaagaac tattcatgct atgtctcaag tatattatgt gtgcaggagg 13860
actcagaaga tggtacaacc caccatggta ctcagaaacc attccttcta ctccatatga 13920
tcaccttgtc ttacggaaag cttttgagaa ggtcattcta ttttggaagt cgcttgtttc 13980
tgagttgtat ataatattta ggtgccactt tgttatattt taaacttcat caaaagcctt 14040
tggattcttt gaggacacat atccccttga ctgactggga gctgaatttc cttgatgtat 14100
agtgcttatt ttgattgtcc tctcctttgt aatactcttg cctataaata aaccttcctt 14160
atcaaaagaa aatgataaaa tgttaaaaat actttgtgat agatgttgac tatagacttt 14220
aaatgatatt ttatctcaat ctttatcaat tcttttttac cttcacatgc aaattatact 14280
acagtatgtg aaatatgcaa tagtttgtcc tgcaattttc atctttcatg tacctctctc 14340
tactatccgt gtattactac tgatttgatg cattaaatgc taattttatt cccaaccttt 14400
actttatgca ggctgtagtc aagcgactta tgacggatgt accatttggt gtgcttctct 14460
caggcggact agattcttca cttgttgctg cagtggccaa ccgctatttg gctgatactg 14520
aagctgcgcg acaatgggga tcacagttgc ataccttttg cgtaggcttg aaggtggtga 14580
tgctcctttt tgtaaacttg ctctgatttg ggaattgcta gctgtatcat cccaatttac 14640
atggaagttc tagaccatta ttgctgtttc ttttccatat atagggattt tctgaaaata 14700
gtaaaaagga aatggaattt ctctttatta aagataccga atgccttttt gggaaatgat 14760
cttacagttc tgaatcgaaa atttcctttt gacattgtga ttaattctga tatgaatttg 14820
gtccatcctg gctttccttc tcttgattgt agattttcct tctcttgtta caattatatg 14880
cctgtctttt tgtcttactc tctgagacag gcaatgtaac tccagttgca aggtttggta 14940
atattgactt gtttatttct gttatcacat cctcctgaaa aagtgctact agcttgtatt 15000
catgcgggaa tttgaaattt tcaagtgtat agctttacaa aatgatcaga actgatcaca 15060
gtcattgctc taaataaaat tttctaaaaa gcaggtattg ttgtgatata atgtgacact 15120
taaataaatt taaacctaca tcctttcgaa atctgtaata gctccactaa tatttcttgt 15180
gctaaactat tccaacacct cttatcgatg ccaattttga cagaccttag tgcacttaaa 15240
agtgtttagt gttctttatg cagtaggtat gttattgtcc aaaatgacag aaataaagtt 15300
agagatttgt tgaaagccac ttggtgtgat tgagcagttc ggagatgagt gatttatcca 15360
ttttatatac ttgtagtgtt ttgttttctt ttgcttttgt cattgtaatt aaggttgtct 15420
tttgtttgac attagggttc tcctgatctg aaagctgcca gagaggtggc agactacctt 15480
ggaacccgtc accatgagtt tcactttaca gtgcaggtaa ctttccacaa agacatccta 15540
ccatgtgacc aggaggtcac gggtttgagc tgtggaaaca accttttgca gaaatgcagc 15600
gtaaggttgc gtacaataga cccttgtggt ccggcccttc cccgaccccg cacatagcgg 15660
gagcttagtg caccgggctg cccatcctaa gctgattttc cttataggag tataaccgct 15720
taagtcaaat tggctatcta cttctttcta ttgctacatc aggaaggaat tgatgcacta 15780
gatgaagtca tttatcatgt tgaaacatat gatgtgacca ctatcagagc cagtacacca 15840
atgtttctca tgtctcggaa gataaagtct ttgggtgtga aaatggttct atctggtgaa 15900
ggttctgatg aaatttttgg cggttattta tatttccaca agggacccaa caaagaggag 15960
tttcaccaag aaacttgtag aaaggtggat ctcattgtca tttcttcagt cattcaggat 16020
tattggaagt aactgtattt ttactagaat acttctttga actatttcta gctctgatgg 16080
acgctttgat gttaatccag attaaagcac ttcatcttta tgattgcttg agggccaaca 16140
aatctacttc agcttggggt gttgaagctc gtgtaccttt cttggataaa gaatttatca 16200
atgttgcaat gaacattgat ccagagtgga aaatggtaac ctacagtgct tccgtctcat 16260
ttctccaccc cccacccccc caaaaaaaag aacagaaaag aattgaaaag agagattctt 16320
ttcttccttc aaggaagaag ggggtaacaa aagaatagct gcaagttatg cagaaaaaat 16380
agtcttccat gtcatgggag accagggttc aaatctgatt gttaaaaaaa acactaggtg 16440
aactcttcta atctgtttaa gccttggtgg acatagttaa ctagtatatg agctagttag 16500
agctgactcg aaaccaccat tattcaaaaa attaagaaag tcttcaatgt tgttccaccg 16560
tgaattcatg atgtgtcgac tttcatgtgt ttatctctgg atagatgaag agtttttttg 16620
gatcttgatt aattctgttt ggcccctttg aggttctgca cgctatttct agactcaaac 16680
ttgctgcgac ctatgttagg tctagagaat atctgcctaa taaaacaatg gaagttacat 16740
tcaagtattt tacctccata ttacaagtca gtgttagtta aatgggtctg atatatggga 16800
tagaaaatgg cttcataccc ataacctcgt taaaatgttg ctgatgtgct gatctcatct 16860
gtacatctag tataatccta attacaaatt aatgcatctc tttgcggtga ctatagtatg 16920
ctgaagaacc tgtcttgaaa cattgcagat cagacctgat ctcggaagaa tagaaaagtg 16980
ggttctacgc aatgcttttg acgatgatca gaatccttat ctgccaaagg tttgtttgtt 17040
ttcttgtatc aggtccaata atcataatct ttgtttattc atttgcataa aaggcagtag 17100
actcccaata cattgtgtac gcttatgcac aaatgtatga gatgtgtatt atatgtgtca 17160
catataaggg tattatatgt acacactcct cccaccttcc acatatatta tgtgtgtgtg 17220
tgtgtgtgta taatgaaatg atgtttgaaa agacattatg atcaagtagt tgatcctcat 17280
ctgctttctg tttttagcat atcttgtata ggcagaagga acagttcagt gatggagttg 17340
gctacagttg gattgatggc ttgaaggatc acgcaagcag tctggtgagc tctttagttg 17400
ttttctcttc cctcctcttc cattctttat cccttccttt tgtggttttg ttgaacacat 17460
ctcatatact tggtccaggt ttctgattct atgttagcga atgcaagttt tgtttacccg 17520
cataacacac ccacgacaaa ggaaggatac tattatagaa ctatttttga gcgatatttc 17580
cccaaggtgg gtcttacata gtcttctttt ccttgttatt catgttcagt ttatgatatt 17640
ttgtcccaaa caatgttttc tttttcttcc tttttctgtt aacaagatcg cttctatctt 17700
cgtatttgtt gataaagtta atcacattgt taactattat ttggagcaga atgctgcgag 17760
ggaaacagtt ccaggtggtc caagtgtggc atgcagcact gcaaaagcag tagaatggga 17820
cgcagcttgg tccaagaatc tagatccatc tggtcgagct gcactcggtg ttcatgcagc 17880
tgcttatgag gatgcatcag aggttaaaac cacagtcgcg acagatactg ctcagaaact 17940
tgacgttgat aaagctgcag tagctgtttg a 17971
<210> 12
<211> 581
<212> PRT
<213> Nicotiana tabacum
<400> 12
Met Cys Gly Ile Leu Ala Val Phe Gly Cys Ile Asp Asn Ser Gln Ala
1 5 10 15
Lys Arg Ser Arg Ile Ile Glu Leu Ser Arg Arg Leu Arg His Arg Gly
20 25 30
Pro Asp Trp Ser Gly Leu His Ser His Asp Asp Cys Tyr Leu Ala His
35 40 45
Gln Arg Leu Ala Ile Val Asp Pro Thr Ser Gly Asp Gln Pro Leu Tyr
50 55 60
Asn Glu Asp Lys Thr Ile Val Val Ala Val Asn Gly Glu Ile Tyr Asn
65 70 75 80
His Arg Asp Leu Arg Glu Lys Leu Lys Ser His Gln Phe Arg Thr Gly
85 90 95
Ser Asp Cys Glu Val Ile Ala His Leu Tyr Glu Asp Tyr Gly Glu Asp
100 105 110
Phe Val His Met Leu Asp Gly Met Phe Ser Phe Val Leu Leu Asp Thr
115 120 125
Arg Asp Lys Ser Phe Ile Ala Ala Arg Asp Ala Ile Gly Ile Thr Pro
130 135 140
Leu Tyr Met Gly Trp Gly Leu Asp Gly Ser Val Trp Phe Ser Ser Glu
145 150 155 160
Met Lys Ala Leu Ser Asp Asp Cys Glu Arg Phe Val Ser Phe Leu Pro
165 170 175
Gly His Ile Tyr Ser Ser Lys Asn Gly Gly Leu Arg Arg Trp Tyr Asn
180 185 190
Pro Pro Trp Tyr Ser Glu Thr Ile Pro Ser Thr Pro Tyr Asp His Leu
195 200 205
Val Leu Arg Lys Ala Phe Glu Lys Ala Val Val Lys Arg Leu Met Thr
210 215 220
Asp Val Pro Phe Gly Val Leu Leu Ser Gly Gly Leu Asp Ser Ser Leu
225 230 235 240
Val Ala Ala Val Ala Asn Arg Tyr Leu Ala Asp Thr Glu Ala Ala Arg
245 250 255
Gln Trp Gly Ser Gln Leu His Thr Phe Cys Val Gly Leu Lys Gly Ser
260 265 270
Pro Asp Leu Lys Ala Ala Arg Glu Val Ala Asp Tyr Leu Gly Thr Arg
275 280 285
His His Glu Phe His Phe Thr Val Gln Glu Gly Ile Asp Ala Leu Asp
290 295 300
Glu Val Ile Tyr His Val Glu Thr Tyr Asp Val Thr Thr Ile Arg Ala
305 310 315 320
Ser Thr Pro Met Phe Leu Met Ser Arg Lys Ile Lys Ser Leu Gly Val
325 330 335
Lys Met Val Leu Ser Gly Glu Gly Ser Asp Glu Ile Phe Gly Gly Tyr
340 345 350
Leu Tyr Phe His Lys Ala Pro Asn Lys Glu Glu Phe His Gln Glu Thr
355 360 365
Cys Arg Lys Ile Lys Ala Leu His Leu Tyr Asp Cys Leu Arg Ala Asn
370 375 380
Lys Ser Thr Ser Ala Trp Gly Val Glu Ala Arg Val Pro Phe Leu Asp
385 390 395 400
Lys Glu Phe Val Asn Val Ala Met Asn Met Asp Pro Glu Ser Lys Met
405 410 415
Ile Arg Pro Asp Leu Gly Arg Ile Glu Lys Trp Val Leu Arg Asn Ala
420 425 430
Phe Asp Asp Asp Gln Asn Pro Tyr Leu Pro Lys His Ile Leu Tyr Arg
435 440 445
Gln Lys Glu Gln Phe Ser Asp Gly Val Gly Tyr Ser Trp Ile Asp Gly
450 455 460
Leu Lys Asp His Ala Ser Arg Leu Val Ser Asp Ser Met Leu Ala Asn
465 470 475 480
Ala Ser Phe Val Tyr Pro His Asn Thr Pro Thr Thr Lys Glu Gly Tyr
485 490 495
Tyr Tyr Arg Thr Ile Phe Glu Arg Tyr Phe Pro Lys Asn Ala Ala Arg
500 505 510
Glu Thr Val Pro Gly Gly Pro Ser Val Ala Cys Ser Thr Ala Lys Ala
515 520 525
Val Glu Trp Asp Ala Ala Trp Ser Lys Asn Leu Asp Pro Ser Gly Arg
530 535 540
Ala Ala Leu Gly Val His Ala Ala Ala Tyr Glu Asp Ala Ser Glu Val
545 550 555 560
Lys Thr Thr Val Ala Thr Asp Thr Pro Gln Lys Leu Glu Val Asp Lys
565 570 575
Ala Ala Val Ala Val
580
<210> 13
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 13
tcacatatat tccctcataa cacac 25
<210> 14
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 14
aagaagcatc ccactctaca gctt 24
<210> 15
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 15
atatcttccc tcacaacact ccaact 26
<210> 16
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер
<400> 16
ctaccggaag gatcaaggtt gt 22
<210> 17
<211> 330
<212> ДНК
<213> Nicotiana tabacum
<400> 17
tcacatatat tccctcataa cacacccatt acaaaggaag catactacta taggatgatt 60
ttcgagcgct ttttcccaca gaattcagct gggctaaccg ttcctggagg agcaagtgtg 120
gcgtgtagca cagctaaagc tgtagagtgg gatgcttctt ggtcaaagaa ccttgatcct 180
tcaggcaggg ctgctattgg tgtacataac tcggcttatg agaatcatgt acctgctatg 240
gctaatggga atttgaccaa aaaaatcatt ggtcgtgtgc cttctatggt agaagttggt 300
gctgctcccg agctcacaat aaagagttag 330
<210> 18
<211> 364
<212> ДНК
<213> Nicotiana tabacum
<400> 18
gaccagattg gagtgggata tatcaacatg gtgattttta cttagcacat caacgtttag 60
caattatcga tcctacttct ggtgatcagc ctctgtttaa tcaagataag actattgttg 120
ttacagtcaa tggagaaatt tacaatcatg agaaacttcg taatcttatg cctaatcaca 180
agttcagaac cggaagtgat tgtgatgtta ttgcacatct ttatgaagaa tatggagaaa 240
attttgtgga catgttggat ggggtgttct cttttgtatt gttggatacg cgcgataaca 300
gctttcttgc tgctcgtgat gcgattggaa ttactcccct ctatattggt tggggacttg 360
atgg 364
<210> 19
<211> 1815
<212> ДНК
<213> Nicotiana tabacum
<400> 19
atgtgcggga tcttggctgt tttgggttgt tctgatgatt ctcaggccaa aagggttcgt 60
gttctcgagc tctctcgcag gtaggttgaa gcatcgtgga ccagattgga gtgggctgta 120
tcaacatggg gactgttact tggcacatca gcgtctagct attgttgatc ctgcttccgg 180
tgatcaacct ctgtttaacg aagataagac gattgttgtt acggtaggta aatggagaga 240
tctacaatca cgagcaactt cgtaagcaaa tgcctaatca taagttccgg actggcagtg 300
actgtgatgt cattgcacac ctagtagtat gaagaacatg gagaagattt tgtggacatg 360
ctggatggga tcttcgcttt tgtgttattg gatactcgag ataacagctt tcttgttgct 420
cgtgatgcca ttggaattac ttccctttat attggttggg gacttgatgg tagggtctgt 480
atgaatatca tctgagctta agggcttgaa tgatgactgc gaacattttg aagttttccc 540
accagggcac ttgtactcta gcaagaatgg cggctttagg aggtggtaca atcctccttg 600
gttctctgag gccattcctt ccactcgtta tgatccctta gttctcaggc gtgcctttga 660
aaatgtaggc tgttatcaaa aggttgatga ctgatgtccc ctttggtgtt ctcctctccg 720
ggggactcga ttcatccttg gttgcttcga ttactgctcg ctacttggct ggtacaaagg 780
ctgccaagca gtggggagca cagcttcatt ccttctgtgt tggccttgag gtagggctca 840
ccggatctca aggctgcaag agaagttgct gactacttgg gaaccgttca ccacgagttt 900
cacttcaccg ttcaggtagg atggaattga tgcaattgaa gatgttattt accatattga 960
gacatacgat gtaacgacaa tcagagcaag cactcctatg ttccttatgt cgcgtaagat 1020
taagtcacta ggagtgaaga tggtcatatc tggggaagga tctgatgaag tgtttggtgg 1080
ctacttgtac tttcacaagg ctcccaacaa ggaagagttc cacaaggaaa catgtcgcaa 1140
ggtagattaa agcgcttcac caatatgact gcttaagagc aaataagtca acatctgcat 1200
ggggtttaga agctagagtc cctttcctag ataaggagtt catcaatgtt gccatgagta 1260
ttgatccaga gtggaagttg agattaaacc agagcaaagg aggattgaaa agtgggctct 1320
aaggagggcc tttgatgatg aggagcatcc ttatctccca aaggtagcac atcctgtata 1380
ggcaaaaaga acaattcagt gatggcgtgg gcatagttgg atagatggac tcaaagcaca 1440
tgctgaacaa catgtaggtg accaatagga tgatgtttaa tgcttcacat atattccctc 1500
ataacacacc cattacaaag gaagcatact actataggat gattttcgag cgctttttcc 1560
cacaggtaga attcagctgg gctaaccgtt cctggaggag caagtgtggc gtgtagcaca 1620
gctaaagctg tagagtggga tgcttcttgg tcaaagaacc ttgatccttc aggcagggct 1680
gctattggtg tacataactc ggcttatgag aatcatgtac ctgctatggc taatgggaat 1740
ttgaccaaaa aaatcattgg tcgtgtgcct tctatggtag aagttggtgc tgctcccgag 1800
ctcacaataa agagt 1815
<210> 20
<211> 155
<212> PRT
<213> Nicotiana tabacum
<400> 20
Met Cys Gly Ile Leu Ala Val Leu Gly Cys Ser Asp Asp Ser Gln Ala
1 5 10 15
Lys Arg Val Arg Val Leu Glu Leu Ser Arg Arg Leu Lys His Arg Gly
20 25 30
Pro Asp Trp Ser Gly Leu Tyr Gln His Gly Asp Cys Tyr Leu Ala His
35 40 45
Gln Arg Leu Ala Ile Val Asp Pro Ala Ser Gly Asp Gln Pro Leu Phe
50 55 60
Asn Glu Asp Lys Thr Ile Val Val Thr Val Asn Gly Glu Ile Tyr Asn
65 70 75 80
His Glu Gln Leu Arg Lys Gln Met Pro Asn His Lys Phe Arg Thr Gly
85 90 95
Ser Asp Cys Asp Val Ile Ala His Leu Tyr Glu Glu His Gly Glu Asp
100 105 110
Phe Val Asp Met Leu Asp Gly Ile Phe Ala Phe Val Leu Leu Asp Thr
115 120 125
Arg Asp Asn Ser Phe Leu Val Ala Arg Asp Ala Ile Gly Ile Thr Ser
130 135 140
Leu Tyr Ile Gly Trp Gly Leu Asp Gly Ser Val
145 150 155
<210> 21
<211> 1818
<212> ДНК
<213> Nicotiana tabacum
<400> 21
atgtgcggga tcttggctgt tttgggttgt tctgatgatt ctcaggccaa aagggttcgt 60
gttctcgagc tctctcgcag gtaggttgaa gcatcgtgga ccagattgga gtgggctgta 120
tcaacatggg gactgttact tggcacatca gcgtctagct attgttgatc ctgcttccgg 180
tgatcaacct ctgtttaacg aagataagac gattgttgtt acggtaggta aatggagaga 240
tctacaatca cgagcaactt cgcaagcaaa tgcctgatca taagttccgg actggaagtg 300
actgtgatgt cattgcacac ctagtagtat gaagaacatg gagaagattt tgtggacatg 360
ctggatggga tcttcgcttt tgtgttactg gatactcgag ataacagctt tcttgttgct 420
cgtgatgcca ttggaattac ttccctttat attggttggg gacttgatgg tagggtctgt 480
atgaatatca tctgagctta agggcttgaa tgatgactgc gaacattttg aagttttccc 540
accaggacac ttgtactcta gcaagaatgg cggctttagg aggtggtaca atcctctttg 600
gttctctgag gctattcctt ccactcctta tgatccctta gttctcaggc gcgcctttga 660
aaatgtaggc tgttatcaaa aggttgatga ctgatgtccc ttttggtgtt ctgctctccg 720
ggggactcga ttcatccttg gttgcttcga ttactgcccg ctacttggct ggcacaaagg 780
ctgccaagca gtggggagca cagcttcatt ccttctgtgt tggccttgag gtagggatca 840
ccggatctca aggctgcaag agaagttgct gactacttgg gaaccgttca ccacgagttt 900
cacttcaccg ttcaggtagg atggaattga tgcaattgaa gatgttattt accatattga 960
gacatacgat gtaacgacaa tcagagcaag cactcctatg ttccttatgt cgcgtaagat 1020
taagtcacta ggagtgaaga tggttatatc tggggaaggc tctgatgaag tgtttggtgg 1080
ctacttgtac tttcacaagg ctcccaacaa ggaagagttc cacaaggaaa catgtcgcaa 1140
ggtagattaa agcacttcac caatatgact gtttaagagc aaataagtca acatctgcat 1200
ggggcttaga agctagagtg cctttcctag ataaggagtt catcaatgtt gccatgagta 1260
ttgatccaga gtggaagttg gtagattaaa ccagagcaaa ggaggattga gaagtgggct 1320
ctaaggaggg cctttgatga tgaggagcat ccctatctcc caaaggtagc acatcctata 1380
caggcagaaa gaacaattca gtgatggcgt aggctatagt tggatagatg gactcaaagc 1440
acatgctgaa caacatgtag gtgaccaata ggatgatgct taatgcttca catatattcc 1500
ctcataacac accgattaca aaggaagcat actattatag gatgattttc gagcgctttt 1560
tcccacaggt agaattcagc tgggctaacc gttcctggag gagcgagtgt ggcgtgtagc 1620
acagctaaag ctgtagagtg ggatgcttct tggtcaaaga accttgatcc ttcaggaagg 1680
gctgctattg gtgtacataa ctcagcttat gagaatcatg aacctgctat ggctaatggg 1740
aatttggcca caaaaatcat tggccgtgcg ccgtctatgg tagaagttgg tgctgctcat 1800
gagctcacaa taaggagt 1818
<210> 22
<211> 155
<212> PRT
<213> Nicotiana tabacum
<400> 22
Met Cys Gly Ile Leu Ala Val Leu Gly Cys Ser Asp Asp Ser Gln Ala
1 5 10 15
Lys Arg Val Arg Val Leu Glu Leu Ser Arg Arg Leu Lys His Arg Gly
20 25 30
Pro Asp Trp Ser Gly Leu Tyr Gln His Gly Asp Cys Tyr Leu Ala His
35 40 45
Gln Arg Leu Ala Ile Val Asp Pro Ala Ser Gly Asp Gln Pro Leu Phe
50 55 60
Asn Glu Asp Lys Thr Ile Val Val Thr Val Asn Gly Glu Ile Tyr Asn
65 70 75 80
His Glu Gln Leu Arg Lys Gln Met Pro Asp His Lys Phe Arg Thr Gly
85 90 95
Ser Asp Cys Asp Val Ile Ala His Leu Tyr Glu Glu His Gly Glu Asp
100 105 110
Phe Val Asp Met Leu Asp Gly Ile Phe Ala Phe Val Leu Leu Asp Thr
115 120 125
Arg Asp Asn Ser Phe Leu Val Ala Arg Asp Ala Ile Gly Ile Thr Ser
130 135 140
Leu Tyr Ile Gly Trp Gly Leu Asp Gly Ser Val
145 150 155
<210> 23
<211> 1818
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК стоп-мутанта ASN5-S: ASN5-S_Q66*
<220>
<221> мутация
<222> (199)..(201)
<400> 23
atgtgtggaa tcttggcttt gttgggttgt tcagatgatt ctcaggccaa aagggttcga 60
gttcttgagc tctctcgcag gcaggttgaa gcatcgtgga ccagattgga gtgggatata 120
tcaacatggt gatttttact tagcacatca acgtttagca attatcgatc ctgcttctgg 180
tgatcagcct ctgtttaatt aagataagac gattgttgtt acagtaggtc aatggagaga 240
tttacaatca tgagaaactt cgtaatctta tgcctaatca caagttcaga actggaagtg 300
attgtgatgt tattgcacat cttgtagtat gaagaatatg gagaaaattt tgtggacatg 360
ttggatgggg tgttctcttt tgtattgttg gatacgcgcg ataacagctt tcttgctgct 420
cgtgatgcaa ttggaattac tcccctatat attggttggg gacttgatgg taggctctgt 480
gtggatttca tctgagctaa aaggattaaa tggtgactgt gaacattttg aagttttccc 540
tcccggtcac ttgtactcga gcaagaatgg cgggtttagg agatggtaca atcctcaatg 600
gttctctgag gctattccat caaatcctta cgacccctta gttttgagac gtgccttcga 660
aaatgtaggc tgttatcaaa cgattgatga ccgatgtccc ctttggtgtt ctgctctccg 720
ggggacttga ttcgtctttg gttgcttctg tcactgctcg ctacttggct ggaacaaaag 780
ctgctaagca atggggagca cagcttcatt ccttctgtgt tggtctcgag gtagggctca 840
ccagatctca aggctgcaag agaagttgct gactatttgg gaaccgttca ccacgagttc 900
accttcacag ttcaggtagg atggaattga tgctattgaa gatgttattt accatatcga 960
gacatacgat gtaacaacga tcagagcaag cactcctatg ttccttatgt cgcgtaagat 1020
taaatcactg ggagttaaga tggtcatatc aggggaaggc tcagatgaac tgtttggcgg 1080
ctatttgtac ttccacaagg ctccgaacaa ggaagaattc catgtggaga catgtcacaa 1140
ggtagataaa agcgcttcac caatacgact gtttgagagc aaataaggca acatcagcat 1200
ggggcttaga agctagagta ccatttctgg ataaagagtt catcaacgtt gctatgagta 1260
tcgatcctga atggaagatg gtagattaaa cacgatcatg gtaggatcga gaagtgggtt 1320
cttaggaagg cttttgatga tgaggagcaa ccctatctcc caaaggtagc atattctgta 1380
ccggcagaaa gaacaattca gtgatggcgt aggctatagt tggatcgatg gactcaaagc 1440
acatgctgaa caacatgtag gtgactgata ggatgatgct taatgctgca catatcttcc 1500
ctcacaacac tccaactaca aaggaagcat actattacag gatgattttc gagaggttct 1560
tcccacaggt agaattcagc aaggctaact gttcctggag gaccgagtat agcttgcagc 1620
acagctaaag ctattgagtg ggatgcttcg tggtcgaaca accttgatcc ttccggtagg 1680
gctgcaatcg gtgtacataa ctcggcttat gacgatcatc tccccgatgt tggtaatggg 1740
aatttggaca caacgatcat cgataatgtg ccgaggatgg taggagtggg tgctgctgca 1800
gagctcacaa taaggagc 1818
<210> 24
<211> 65
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность стоп-мутанта ASN5-S: ASN5-S_Q66*
<400> 24
Met Cys Gly Ile Leu Ala Leu Leu Gly Cys Ser Asp Asp Ser Gln Ala
1 5 10 15
Lys Arg Val Arg Val Leu Glu Leu Ser Arg Arg Leu Lys His Arg Gly
20 25 30
Pro Asp Trp Ser Gly Ile Tyr Gln His Gly Asp Phe Tyr Leu Ala His
35 40 45
Gln Arg Leu Ala Ile Ile Asp Pro Ala Ser Gly Asp Gln Pro Leu Phe
50 55 60
Asn
65
<---
Реферат
Изобретение относится к области биохимии, в частности к мутантному или трансгенному растению табака со сниженными уровнями аспарагина и сниженными уровнями акриламида в аэрозоле, полученном после нагревании или сгорания растения табака, а также к полученному из него растительному материалу и пригодному для потребления продукту. Также раскрыт способ получения растительного материала со сниженными уровнями аспарагина и сниженными уровнями акриламида в аэрозоле, полученном из указанного растительного материала. Изобретение также относится к табачному изделию для курения, содержащему вышеуказаный растительный материал, и устройству, образующему аэрозоль. Изобретение позволяет эффективно получать растительный материал табака со сниженными уровнями аспарагина и сниженными уровнями акриламида в аэрозоле, полученном из указанного растительного материала. 7 н. и 8 з.п. ф-лы, 22 ил., 3 табл., 5 пр.
Комментарии