Способы и рационы для защиты от химической интоксикации и связанных с возрастом заболеваний - RU2674681C2
Код документа: RU2674681C2
Чертежи












































Описание
ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет предварительной заявки на патент США с порядковым номером 61/763797, поданной 12 февраля 2013 года, раскрытие которой включено в данный документ во всей ее полноте посредством ссылки.
ЗАЯВЛЕНИЕ ОТНОСИТЕЛЬНО ФИНАНСИРУЕМЫХ ИЗ ФЕДЕРАЛЬНОГО БЮДЖЕТА ИССЛЕДОВАНИЙ
"Настоящее изобретение было выполнено при правительственной поддержке на основании контракта № Р01 AG034906-01, выданного Национальными институтами здравоохранения. Правительство обладает определенными правами на настоящее изобретение.
ОБЛАСТЬ ТЕХНИКИ
По меньшей мере в одном аспекте настоящее изобретение относится к способу ослабления симптомов связанного с возрастом заболевания и химической интоксикации.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Ограничение калорийности (CR) без недостаточности питания стабильно демонстрировало увеличение продолжительности жизни у ряда животных моделей, включая дрожжи, С.elegans и мышей. Тем не менее, влияние CR на срок жизни у не являющихся человеком приматов остается под вопросом и в значительной степени находится под влиянием состава рациона. Полагают, что связанное с CR увеличение срока жизни у модельных организмов обусловлено его влиянием на GH, GHR, что в дальнейшем приводит к снижению уровней IGF-1 и инсулина, а также передачи сигнала. Влияние сигнального пути инсулин/IGF-1 на продолжительность жизни было открыто у С.elegans, когда было показано, что мутации в данном пути, который регулируется наличием нутриентов, вызывают двукратное увеличение срока жизни. В других исследованиях выявили, что мутации в ортологах генов, выполняющих функции в сигнальных путях, связанных с ростом, включая Tor-S6K и Ras-cAMP-PKA, способствуют старению у множества модельных организмов, что обеспечивало, таким образом, подтверждение консервативной регуляции старения генами сигнальных путей, реагирующих на поступление нутриентов и стимулирующих рост.
Недавно было показано, что у людей с дефектом рецептора гормона роста (GHRD) с существенно сниженными уровнями сывороточного IGF-1 и инсулина не наблюдается смертность от рака или диабета, и несмотря на большую частоту ожирения, суммарная смертность от болезней сердца и инсульта в данной группе аналогична таковой у их родственников. Об аналогичной устойчивости к раку также сообщали в исследовании, в котором наблюдали 230 пациентов с GHRD.
Ограничение белков или ограничение определенных аминокислот, таких как метионин и триптофан, может отчасти объяснить влияние ограничения калорийности на продолжительность жизни и риск возникновения заболеваний, поскольку ограничение белков снижает уровни IGF-1, может увеличивать продолжительность жизни у млекопитающих независимо от потребления калорий и, как было также показано, уменьшает частоту возникновения рака в моделях на грызунах.
Соответственно, существует потребность в применении диеты, которая может ослаблять симптомы связанного с возрастом заболевания как у субъектов, желающих изменить свой рацион на длительный срок, так и у тех, которые рассматривают лишь периодическое применение диеты, но в остальное время продолжают придерживаться своего нормального рациона.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение разрешает одну или несколько проблем предшествующего уровня техники путем обеспечения по меньшей мере в одном варианте осуществления способа ослабления симптома старения или связанных с возрастом симптомов. Данный способ включает стадию, на которой определяют средний суточный уровень потребления белков субъектом. В одном варианте средний суточный уровень потребления белков выражают в виде процента калорий, приходящихся на белки, которые субъект в среднем потребляет за день. С учетом потребления белков определяют относительные количества калорий, приходящихся на белки из источников животного и растительного происхождения. Субъекта периодически обеспечивают высокопитательным рационом с низким содержанием белков вместо его нормального рациона, если определяют, что средний суточный уровень потребления белков субъектом выше, чем предварительно установленный предельный уровень потребления белков, если субъект моложе предварительно установленного возраста.
В другом аспекте обеспечивают способ снижения у субъекта уровней глюкозы и/или IGF-1. Способ включает стадию периодического обеспечения субъекта низкокалорийным рационом и/или рационом с низким содержанием белков, содержащим менее приблизительно 10 процентов калорий, приходящихся на источники белка растительного происхождения. Уровни глюкозы и/или IGF-1 у субъекта контролируют для определения необходимости увеличения или уменьшения потребления белков.
В другом аспекте рацион с низким содержанием белков включает пищевую добавку, обеспечивающую избыточные уровни заменимых аминокислот, потребляемых в течение периода 5-7 дней вместе с очень низкими количествами белков или безбелковым рационом. В одном варианте рацион с низким содержанием белков чередуют с рационом с нормальным содержанием белков. В таких вариантах рацион с низким содержанием белков на основе источников растительного происхождения обеспечивают в течении 7 дней каждые 2 недели - 2 месяца с нормальным рационом в течение 1-7 недель между ними. Как правило, пищевая добавка обеспечивает в качестве источника азота одну или несколько из следующих аминокислот: аланина, аспарагиновой кислоты, цистеина, глутаминовой кислоты, глицина, гистидина, пролина, серина и тирозина, при фактическом исключении изолейцина, лейцина, лизина, метионина, фенилаланина, треонина, триптофана, валина и аргинина, так что изолейцин, лейцин, лизин, метионин, фенилаланин, треонин, триптофан, валин и аргинин вместе присутствуют в количестве, составляющем менее 5% от общей массы рациона субъекта. В дополнительном варианте изолейцин, лейцин, лизин, метионин, фенилаланин, треонин, триптофан, валин и аргинин в комбинации присутствуют в количестве, составляющем менее 3% от общей массы рациона субъекта.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
ФИГУРА 1. В таблице 9 показан общий обзор содержания калорий в рационе, имитирующем воздержание от пищи, адаптированном для людей-субъектов. Рацион, имитирующий воздержание от пищи (FMD), Prolon, индуцирует реакцию, подобную той, которая наблюдается при режиме воздержания от пищи, при максимальной питательности. Показано количество потребляемых калорий для каждого из 5 дней потребления рациона, а также установленное количество ккал на фунт массы тела. Уменьшение потребления калорий в течение 5 дневного режима питания (Δ5-день) показано либо 1) из расчета 2000 калорий на день потребления рациона, либо 2) из расчета на рационы с 2800,2400 и 2000 калорий для индивидуума с массой >200, 150-200 и <150 фунтов, соответственно.
ФИГУРА 2. В таблице 10 показано определенное содержание макронутриентов для каждого дня потребления рациона, скорректированное для человека-субъекта массой 180-200 фунтов. Содержание макронутриентов для каждого дня из 5 дней режима потребления FMD приведено из расчета на индивидуума со средней массой тела 180-200 фунтов. Потребление калорий в 1 день потребления рациона уменьшено в меньшей степени по сравнению со следующими днями (2-5) для того, чтобы предоставить организму возможность приспособиться к низкому потреблению калорий. Представлен % калорий, приходящихся на жиры, углеводы (в частности, на сахар) и белки, для каждого дня режима потребления Prolon.
ФИГУРА 3. В таблице 11 показано определенное содержание микронутриентов для каждого дня потребления рациона, скорректированное для человека-субъекта массой 180-200 фунтов, в одном варианте по настоящему изобретению. Содержание микронутриентов для каждого дня из 5 дней режима потребления FMD приведено из расчета на индивидуума со средней массой тела 180-200 фунтов. Процент от суточной нормы (% DV) рассчитан из расчета на рацион с 2000 калорий. * для ряда микронутриентов DV не определена; показанные значения приведены из расчета на, стандартное суточное потребление (RDI).
ФИГУРА 4. При использовании моделей пропорциональных рисков Кокса для показателей смертности по всем причинам и смертности от рака были выявлены статистически значимые (р<,05) взаимосвязи между возрастом и группами потребления белков. Основываясь на данных моделях, прогнозируемый остаток вероятной продолжительности жизни рассчитывали для каждой группы потребления белков с учетом возраста в начале исследования. Исходя из результатов, низкое потребление белков, по-видимому, обладает защитным эффектом в отношении смертности по всем причинам и смертности от рака до наступления возраста 66 лет, при этом с этого момента оно становится неблагоприятным. Не выявлено значимых взаимосвязей между сердечно-сосудистым заболеванием (CVD) и смертностью от диабета.
ФИГУРА 5. Уровни IGF-1 в сыворотке крови у респондентов в возрасте 50-65 лет и 66+ лет, сообщавших о низком, умеренном или высоком уровне потребления белков. Уровень IGF-1 у респондентов в возрасте 50-65 лет является существенно более низким среди потребляющих низкое количество белков по сравнению с потребляющими высокое количество белков (Р=0,004). В возрасте 66+ лет различие между высоким и низким потреблением белков становится малозначимым (Р=0,101). Когорта, для которой определяли уровни IGF-1, включает 2253 субъекта. Среди субъектов в возрасте 50-65 лет (n=1125) 89 относились к категории низкого потребления белков, 854 относились к категории умеренного потребления белков и 182 относились к категории высокого потребления белков. Среди субъектов в возрасте 66+ лет (n=1128) 80 относились к категории низкого потребления белков, 867 относились к категории умеренного потребления белков и 181 относился к категории высокого потребления белков. *Р<0,01.
ФИГУРА 6. (А) Частота возникновения опухолей у самцов мышей C57BL/6 в возрасте 18 недель, которым имплантировали 20000 клеток меланомы (В16) и которых кормили либо рационом с высоким содержанием белков (n=10), либо с низким содержанием белков (n=10). (В) Прогрессирование увеличения объема опухоли В16 у (18-нед.) самцов мышей C57BL/6, которых кормили либо рационом с высоким содержанием белков (n=10), либо с низким содержанием белков (n=10). (С) Уровень IGF-1 в день 16 у (18-нед.) самцов мышей C57BL/6, которых кормили либо рационом с высоким содержанием белков (n=5), либо с низким содержанием белков (n=5). (D) Уровень IGFBP-1 в день 16 у (18-нед.) самцов мышей C57BL/6, которых кормили либо рационом с высоким содержанием белков (n=10), либо с низким содержанием белков (n=10). (Е) Прогрессирование развития опухоли меланомы В16 у самок мышей GHRKO в возрасте 10 месяцев (n=5) в сравнении с соответствующими им по возрасту и полу однопометными контролями (контр.; n=7). (F) Частота возникновения опухолей у самок мышей BAFB/c в возрасте 12 недель, которым имплантировали 20000 клеток рака молочной железы (4Т1), которых кормили либо рационом с высоким содержанием белков (n=10), либо с низким содержанием белков (7%; n=10). (G) прогрессирование развития рака молочной железы 4Т1 у (12-нед.) самок мышей BAFB/c, которых кормили либо рационом с высоким содержанием белков (n=10), либо с низким содержанием белков (n=10). (Н) Уровень IGF-1 в день 16 у (12-нед.) самок мышей BAFB/c, которых кормили либо рационом с высоким содержанием белка казеина (n=5), либо с низким содержанием белка казеина (n=5). (I) Уровень IGFBP-1 в день 16 у (12-нед.) самок мышей BAFB/c, которых кормили либо рационом с высоким содержанием белка казеина (n=10), либо с низким содержанием белка казеина (n=10). (J) Уровень IGF-1 в день 16 у (12-нед.) самок мышей BAFB/c, которых кормили либо рационом с высоким содержанием белка казеина (n=5), либо с низким содержанием белка казеина (n=5). (К) Уровень IGFBP-1 в день 16 у (12-нед.) самок мышей BAFB/c, которых кормили либо рационом с высоким содержанием соевого белка (n=10), либо с низким содержанием соевого белка (n=10). (L) Выживаемость и (М) частота возникновения мутаций в ДНК у дрожжей, которые подвергали воздействию стандартной смеси аминокислот в концентрации 0,5х, 1х или 2х. (N) PDS и STRE активность у дрожжей в среде, которая содержит только Trp, Leu и His, в сравнении с теми, которые выращивали в присутствии всех АА. (О) Делеция в Ras2 защищает от нестабильности генома, индуцированной оксидативным стрессом, что определяют по частоте возникновения мутаций в ДНК (Can1) у дикого типа (DBY746) и ras2Δ-мутантов, которых длительно подвергали воздействию 1 мМ Н2О2. (Р) Модель изучения влияния аминокислот на старение и нестабильность генома у S. cerevisiae. Аминокислоты активируют пути Tor-Sch9 и Ras-cAMP-PKA, также активируемые глюкозой, и способствуют зависимой от возраста и оксидативного стресса нестабильности генома частично посредством понижения активности Gis1 и Msn2/4. *Р<0,05, **Р<0,01, ***Р<0,001, ****Р<0,0001.
ФИГУРА 7. Воздействие потребления белков на массу тела у молодых и старых мышей. (А) Молодых (в возрасте 18 недель) (n=10) и старых (в возрасте 24 месяцев) (n=6) мышей C57BL/6 кормили рационом с высоким содержанием белков (18%). (В) Молодых (в возрасте 18 недель) (n=10) и старых (в возрасте 24 месяцев) (n=6) мышей C57BL/6 кормили рационом с низким содержанием белков (4%).
ФИГУРА 8. Таблица 12. Взаимосвязь между смертностью и потреблением белков.
ФИГУРА 9. (А) 30-Дневное изменение массы тела у самцов мышей C57BL/6 в возрасте 18 недель, которых кормили изокалорийными рационами, различающимися по содержанию белков: либо высокое (18%), либо низкое (4%). (В) 30-Дневное потребление пищи в ккал/день у самцов мышей C57BL/6 в возрасте 18 недель, которых кормили изокалорийными рационами, различающимися по содержанию белков: либо высокое (18%), либо низкое (4%). (С) Уровень IGFBP-2 в день 16 у самцов (18-нед.) мышей C57BL/6, которых кормили либо рационом с высоким содержанием белков (n=10), либо с низким содержанием белков (n=10). (D) Уровень IGFBP-3 в день 16 у самцов (18-нед.) мышей C57BL/6, которых кормили либо рационом с высоким содержанием белков (n=10), либо с низким содержанием белков (n=10). (Е) 30-Дневное изменение массы тела у самок 12-недельных мышей BALB/c, которых кормили изокалорийными рационами, различающимися по содержанию белков: либо высокое (18%), либо низкое (4%). (F) 30-Дневное потребление пищи в ккал/день у самок мышей BALB/c в возрасте 12 недель, которых кормили изокалорийными рационами, различающимися по содержанию белков: либо высокое (18%), либо низкое (4%). (G) Уровень IGF-3 в день 16 у самок мышей BALB/c в возрасте 18 недель, которых кормили либо рационом с высоким содержанием белков (n=10), либо с низким содержанием белков (n=10). *Р<0,05, **Р<0,01, ***Р<0,001, ****Р<0,0001; ANOVA.
ФИГУРА 10. Прогрессирование увеличения объема опухоли меланомы В16 у самок мышей GHRKO в возрасте 10 месяцев (n=5) в сравнении с соответствующими им по возрасту и полу контролями дикого типа (Wt; n=13). **Р<0,01.
ФИГУРА 11. (А) Выживаемость дрожжей в динамике и (В) ослабление зависимой от возраста нестабильности генома показаны в виде частоты возникновения мутаций в гене САШ (определяли как количество Canr-мутантов/106 клеток) у дикого типа (DBY746) в сравнении с ras2Δ-мутантами. (С) Выживаемость в динамике дикого типа и ras2Δ-мутантов, которых длительно обрабатывали 1 мМ Н2О2. (D) Дефицит Ras2 защищает от индуцированной стрессом нестабильности генома (частота возникновения мутаций Canr). *Р<0,05, **Р<0,01, ***Р<0,001, ****Р<0,0001.
ФИГУРЫ 12а)-f). IGF-1 ослабляет взаимосвязь между потреблением белков и смертностью. Исходя из результатов, полученных при использовании моделей пропорциональных рисков Кокса для изучения взаимосвязи между потреблением белков и уровнем IGF-1 в отношении смертности, прогнозируемые соотношения рисков рассчитывали по IGF-1 как для групп с умеренным потреблением, так и с высоким потреблением белков по отношению к группе с низким потреблением белков. В возрастной группе 50-65 лет не выявлено значимых взаимосвязей между потреблением белков и уровнем IGF-1 в отношении смертности по всем причинам (3а) или смертности от CVD (3b). Однако взаимосвязь между уровнем IGF-1 и потреблением белков, высокое по сравнению с низким, была значимой (р=,026) в отношении смертности от рака (3с) для субъектов в возрасте 50-65 лет. Результаты показывают, что повышение уровня IGF-1 на каждые 10 нг/мл повышает риск смертности от рака на 9% для группы с высоким потреблением белков по сравнению с группой с низким потреблением белков (HRвысокоепотреблениебелков×IGF-1: 1-09; 95% CI: 1,01-1,17). Взаимосвязь между потреблением белков и уровнем IGF-1 была значимой для респондентов в возрасте 66+ лет только в отношении смертности от CVD. У тех, кто потреблял рацион с высоким или умеренным содержанием белков, был пониженный риск CVD, если уровень IGF-1 также был низким; однако, если уровень IGF-1 был повышен, то преимущество отсутствовало.
ФИГУРА 13. Таблица 13. Характеристики выборки.
ФИГУРА 14. Таблица 14. Взаимосвязь между потреблением белков и смертностью (N=6381).
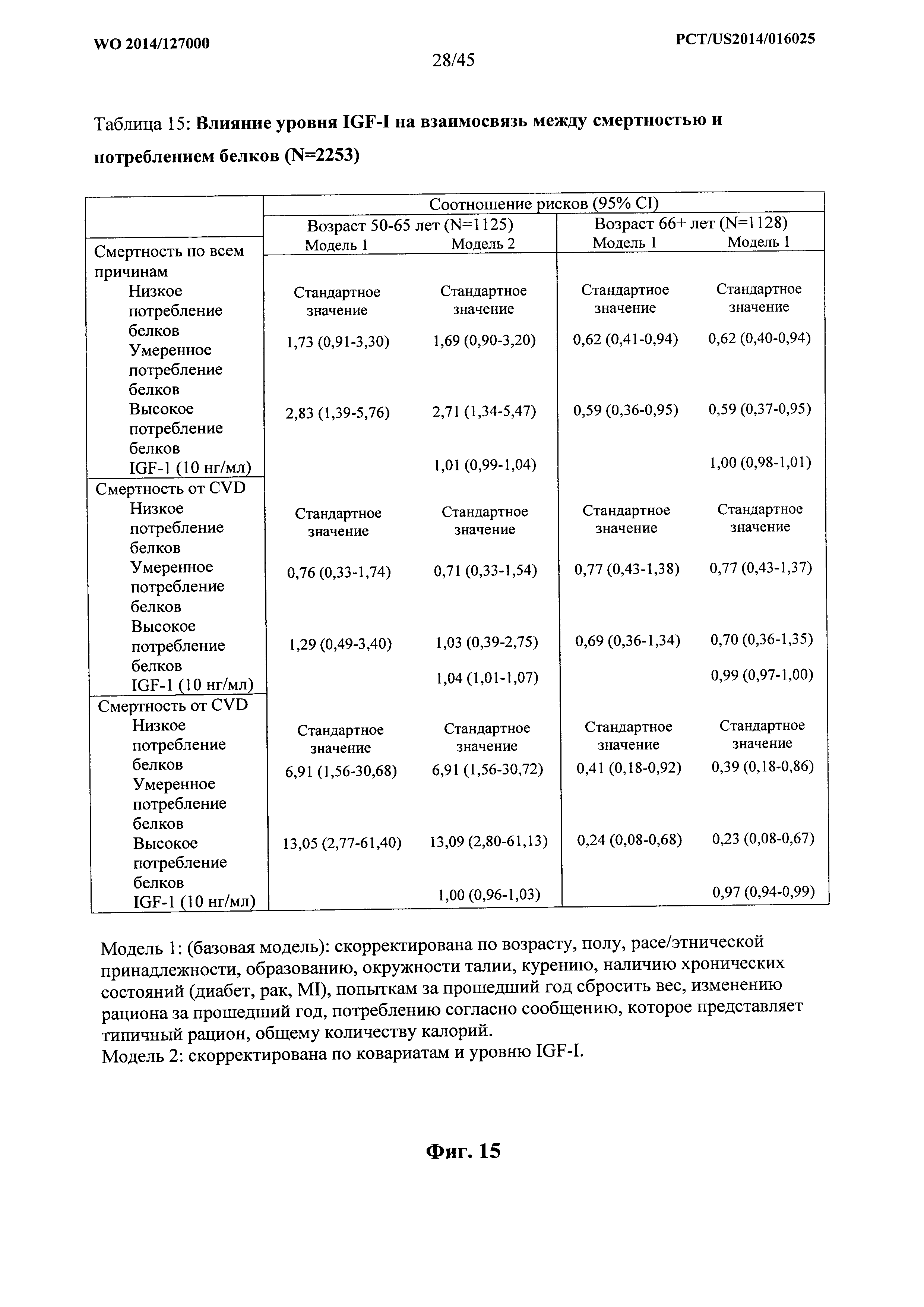
ФИГУРА 15. Таблица 15. Влияние уровня IGF-1 на взаимосвязь между потреблением белков и смертностью (N=2253).
ФИГУРА 16. Таблица 16. Соотношения рисков для взаимосвязи между потреблением белков и уровнем IGF-1 в отношении смертности.
ФИГУРА 17. Таблица 17. Влияние белков животного и растительного происхождения на взаимосвязь между потреблением белков и смертностью.
ФИГУРА 18. Таблица 18. Среднее значение HbA1c, распространенность диабета и среднее значение BMI, скорректированные по возрасту и потреблению белков.
ФИГУРА 19. Таблица 19. Взаимосвязи между смертностью от диабета и потреблением белков среди участников, у которых отсутствовал диабет в начале исследования.
ФИГУРЫ 20A-D. Люди-субъекты принимали участие в 3 циклах 5-дневного потребления низкокалорийного и высокопитательного рациона с низким содержанием белков, имитирующего воздержание от пищи (FMD, отмечено зеленым, смотри текст), с последующим потреблением нормального рациона в течение примерно 3 недель (отмечено коричневым) (А). Кровь отбирали перед началом 5-дневного потребления рациона и при его завершении (временные точки А и В), а также через 5-8 дней после завершения 3-го 5-дневного потребления FMD (временная точка С). 5-Дневное потребление рациона существенно снижало уровни глюкозы (В), IGF-1 (с) и IGFBP-1 (D) в крови. Глюкоза *, р<0,05, N=18; IGF-1, **, р<0,01, *р<0,05, N=16; IGFBP-1, **, р<0,01, N=17; при этом все статистические тесты были выполнены в виде парного t-теста, двустороннего по исходным значениям.
ФИГУРА 21. Обеспечиваемые макронутриентами калории в экспериментальных рационах в %. Стандартный корм AIN93G являлся референтным рационом, и его предоставляли всем мышам. Данный рацион являлся основой для всех экспериментальных рационов, модифицированных по составу макронутриентов (жиров, белков и углеводов). В рационе с низким содержанием углеводов LCHP количество приходящихся на углеводы калорий было уменьшено до 20% по сравнению с составом AIN93G (13% против 63,9%), но он содержал больше белков (45,2%) и жиров (41,8%). В рационах 20% Р-1 (соевое масло в качестве источника жиров) и 20% Р-2 (кокосовое масло в качестве источника жиров) количество калорий, приходящихся на источники белков, было уменьшено до 20% в сравнении с составом AIN93G; рацион 0% Р не содержал белков; все данные рационы являлись изокалорийными по отношению к стандартному корму AIN93G. Кетогенный рацион с высоким содержанием жиров 60% HF был создан для обеспечения 60% потребляемых калорий, приходящихся на источники жиров, при этом калории, поступающие из белков и углеводов, пропорционально уменьшили. Рацион 90% HF представлял собой кетогенный рацион, содержащий 90% жиров, обеспечивая при этом лишь минимальное количество углеводов (менее 1%) и половину количества белков (9%). Подробное описание состава рациона и содержание калорий приведены в таблице 14.
ФИГУРА 22. Ограничение калорийности снижает массу тела, уровни глюкозы и IGF-1. А) Самок мышей CD-I в возрасте 12-15 недель либо кормили ad lib (серые квадраты) стандартным кормом для грызунов AIN93G, содержали на рационах AIN93G с ограниченной на 40%, 60%, 80% и 90% калорийностью (треугольники), либо ограничивали прием ими пищи (STS, зеленые прямоугольники) до потери мышами 20% от их исходной массы (точечная линия). N=5 на экспериментальную группу. Все данные представлены в виде среднего значения ± SEM. В) Линейное приближение для зависимости степени жесткости режима CR от продолжительности (в днях) до достижения 80% массы тела. С) Уровни глюкозы в крови у мышей при достижении 80% массы тела. Красная линия представляет среднее значение; *р<0,05, *** р<0,001, ANOVA, множественное сравнение Тьюки. С) Уровни IGF-1 в сыворотке крови у мышей при достижении 80% массы тела. Красная линия представляет среднее значение; *** р<о,001, ANOVA, множественное сравнение Тьюки.
ФИГУРА 23. Влияния рационов с определенным составом макронутриентов на массу тела, потребление пищи, уровни глюкозы и IGF-1 в сыворотке крови. Пятерых самок мышей CD-I в возрасте 12-15 недель кормили ad lib либо стандартным кормом для грызунов AIN93G (черные круги), либо А) двумя разными рационами с низким содержанием белков (20% Р-1 и 20% Р-2), рационом с низким содержанием углеводов, но с высоким содержанием белков (LCHP), белководефицитным рационом (0% Р), либо В) рационом с высоким содержанием жиров (60% HF) и кетогенным рационом (90% HF). Подробный обзор макронутриентов представлен в таблице 1. С) Суточное потребление калорий ad lib для рационов AIN93G, 20% Р-1,20% Р-2, LCHP и 0% P. D) Суточное потребление калорий ad lib для рационов 60% HF и 90% HF; AIN93G показан в качестве стандарта. Все данные представлены в виде среднего значения ± SEM. Е) Уровни IGF-1 в сыворотке крови после 9 дневного кормления ad lib. Линии представляют среднее значение; *р<0,05, *** р<0,001, ANOVA, множественное сравнение Тьюки при сравнении с контрольным AIN93G.
ФИГУРА 24. А) Тест на стрессоустойчивость для рационов с ограниченной калорийностью и определенным составом макронутриентов. Мышей кормили ad lib (AIN93G), ограничивали прием ими пищи в течение 60 ч. (STS) или кормили рационами с ограниченной на 50% калорийностью с определенным составом макронутриентов (AIN93G, LCHP, 0% Р, 60% HF, 90% HF) в течение 3 дней (зеленый квадрат) до внутривенного введения доксорубицина (24 мг/кг, красная пунктирная линия). Выживаемость контролировали в течение 25 дней после введения, после чего оставшихся животных рассматривали как выживших. В) Уровни глюкозы в крови после 3 дней кормления ad lib и рационов CR, а также после 60 ч. STS. Линии представляют среднее значение; *р<0,05, *** р<0,001, ANOVA, множественное сравнение Тьюки. С помощью программного обеспечения для статистического анализа Prism (GraphPad Software) строили график с использованием данных выживаемости из парных объединенных экспериментов.
ФИГУРА 25. Прогрессирование развития опухоли глиомы GL26 и рака молочной железы 4Т1 in vivo. А) Прогрессирование развития подкожной опухоли глиомы мышей GL26 показано в виде общего объема опухоли в мм3. Как только опухоли можно было пальпировать под кожей на 10 день, начинали проводить измерение опухолей. Животных кормили ad lib либо AIN93G (N=5) в качестве контроля, либо рационом с низким содержанием белков 20% Р-1 (N=6). Все данные представлены в виде среднего значения ± SEM. В) Прогрессирование развития подкожного рака молочной железы 4Т1 мышей показано в виде общего объема опухоли в мм. Как только опухоли можно было пальпировать под кожей на 12 день, начинали проводить измерение опухолей. Контрольных животных (N=10) не обрабатывали, и опухоль быстро прогрессировала, достигая конечного объема 2000 мм к 54 дню после имплантации опухоли. Животных в группе цисплатина (CDDP) (N=9) инъецировали в дни 15, 33 и 44. Первую дозу CDDP доставляли в количестве 12 мг/кг путем внутривенной инъекции, две последующие инъекции производили в количестве 8 мг/кг, чтобы избежать химической интоксикации. Мышей в группе 50% ICR + CDDP (N=9) кормили в прерывистых режимах рационом AIN93G с уменьшением суточного потребления калорий до 50% в сравнении с контрольной группой в течение трех дней (ICR, зеленый квадрат) перед инъекцией цисплатина. Схема инъекций такая же, как для группы CDDP. Все данные представлены в виде среднего значения ± SEM; *** р<0,001, ANOVA, множественного сравнения Тьюки при сравнении с контролем.
ФИГУРА 26. А) Потребление пищи в ккал/день животными, которых кормили ad lib либо (серые квадраты) стандартным кормом для грызунов AIN93G, кормили рационами AIN93G с ограниченной на 40%, 60%, 80% и 90% калорийностью (треугольники), либо ограничивали прием ими пищи (STS, зеленые прямоугольники) до потери мышами 20% от их исходной массы тела. В) Уровни глюкозы в крови у мышей после 48 ч содержания на всех экспериментальных рационах. Линии представляют среднее значение; ** р<0,01, *** р<0,001, ANOVA, множественное сравнение Тьюки.
ФИГУРА 27. Уровни глюкозы в крови после 9 дней кормления ad lib указанными экспериментальными рационами. Линии представляют среднее значение.
ФИГУРА 28. А) Профиль массы тела у мышей, которых кормили ad lib (AIN93G) или кормили рационами с ограниченной на 50% калорийностью с определенными составами макронутриентов (60% HF, LCHP, 0% Р) в течение 3 дней (зеленый квадрат) до внутривенного введения доксорубицина (24 мг/кг, красная пунктирная линия). В) Профиль массы тела у мышей, которых кормили ad lib (AIN93G), кормили рационами с ограниченной на 50% калорийностью с определенными составами макронутриентов (AIN93G, 90% HF) в течение 3 дней (зеленый квадрат) или ограничивали прием ими пищи в течение 60 ч. (зеленый квадрат) до внутривенного введения доксорубицина (24 мг/кг, красная пунктирная линия).
ФИГУРА 29. Калории в %, обеспечиваемые макронутриентами классического кетогенного рациона и модифицированным рационом Аткинса.
ФИГУРА 30. Таблица 20. Обзор содержания макронутриентов и калорий в экспериментальных рационах.
ФИГУРА 31. Таблица 21. Подробный состав рационов с определенным составом макронутриентов.
ФИГУРА 32. Таблица 22. Схема адаптации к рационам с определенным составом макронутриентов.
ФИГУРА 33. Таблица 23. Состав рационов с ограниченной калорийностью.
ФИГУРА 34. Таблица 24. Состав рационов с ограниченной калорийностью и определенным составом макронутриентов;
ПОДРОБНОЕ ОПИСАНИЕ
Далее будет предоставлена подробная информация о предпочтительных композициях, вариантах осуществления и способах по настоящему изобретению. Необходимость масштабирования фигур отсутствует. Раскрытые варианты осуществления по настоящему изобретению являются лишь иллюстративными и могут быть осуществлены в разных и альтернативных формах. Таким образом, конкретные подробности, раскрытые в данном документе, не следует истолковывать как ограничивающие, а лишь как описательную основу для любого аспекта настоящего изобретения и/или как описательную основу для обучения специалиста в данной области применению различным образом настоящего изобретения.
За исключением примеров или случаев, где явно указано иное, все числовые величины в данном описании, обозначающие количества материала или условия реакции и/или применения, следует понимать как измененные словом "приблизительно" в описании наиболее широкого объема изобретения. Практическое применение в указанных числовых пределах является в общем предпочтительным. Кроме того, если явно не указано иное, процент, "части" и величины соотношений приведены по массе; описание группы или класса материалов, подходящих или предпочтительных для данной цели, в связи с настоящим изобретением предполагает, что смеси каких-либо двух или более представителей группы или класса являются в равной степени подходящими или предпочтительными; описание составляющих в химических терминах относится к составляющим при добавлении к какой-либо указанной в описании комбинации и не обязательно исключает химическое взаимодействие между компонентами смеси при смешивании; первое определение сокращения или другой аббревиатуры распространяется на все последующие случаи использования в данном документе той же аббревиатуры и применяется с необходимыми изменениями по отношению к грамматическим вариантам первоначально определенных аббревиатур; и, если явно не указано иное, количественное определение свойства выполняют по той же методике, которая упоминается ранее или позднее для того же самого свойства.
Настоящее изобретение не ограничивается конкретными вариантами осуществления и способами, которые описаны ниже, поскольку конкретные компоненты и/или условия могут, разумеется, варьировать. Кроме того, используемая в данном документе терминология, применяется только с целью описания конкретных вариантов осуществления настоящего изобретения и не предназначена для ограничения каким-либо образом.
Используемые в данном описании и прилагаемой формуле изобретения формы единственного числа включают ссылки на множественное число, если контекст четко не указывает иное. Например, ссылка на компонент в единственном числе подразумевает включение множества компонентов.
Термин "незаменимая аминокислота" относится к аминокислотам, которые не могут синтезироваться организмом. У людей незаменимые аминокислоты включают изолейцин, лейцин, лизин, метионин, фенилаланин, треонин, триптофан, валин. Кроме того, следующие аминокислоты также являются незаменимыми у людей при определенных условиях: гистидин, тирозин и селеноцистеин.
Термины "килокалория" (ккал) и "калория" означают пищевую калорию. Термин "калория" относится к так называемой малой калории.
Термин "субъект" относится к человеку животному, включая всех млекопитающих, таких как приматы (в частности, высшие приматы), овца, собака, грызуны (например, мышь или крыса), морская свинка, коза, свинья, кошка, кролик и корова.
В одном варианте осуществления настоящего изобретения обеспечивают способ ослабления симптома старения или связанных с возрастом симптомов. Например, с помощью способа согласно варианту осуществления настоящего изобретения можно предупреждать или лечить диабет или рак и отсрочить связанную с возрастом смертность и другие связанные с возрастом заболевания. В других вариантах данные способы применимы для снижения у субъекта уровней глюкозы и/или IGF-1. В других вариантах данные способы применимы для лечения (например, ослабления симптома) химической интоксикации у субъекта. Как правило, с помощью способа ослабляют один или несколько симптомов данных состояний. В частности, обеспечивают способ увеличения продолжительности жизни субъекта. В контексте настоящего изобретения увеличение продолжительности жизни означает повышение вероятности более долгой жизни субъекта. Например, если многих субъектов, имеющих такой же профиль (массу, возраст, уровни глюкозы, уровень инсулина и т.д., смотри ниже) что и данный субъект, подвергают воздействию способа по настоящему изобретению, средняя выживаемость увеличивается. Данный способ включает стадию, на которой определяют средний суточный уровень потребления белков субъектом. В одном варианте средний суточный уровень потребления белков выражают в виде процента калорий, приходящихся на белки, которые субъект в среднем потребляет за день. Количество потребляемых субъектом белков можно оценить путем опроса субъекта или заполнения в письменном виде опросника относительно суточного и недельного потребляемого субъектом количества белков, жиров и углеводов. С учетом потребления белков определяют относительные количества калорий, приходящихся на белки из источников животного и растительного происхождения.
Субъекта обеспечивают рационом с низким содержанием белков, если определяют, что средний суточный уровень потребления белков субъектом выше, чем предварительно установленный предельный уровень потребления белков, если субъект моложе предварительно установленного возраста. Как правило, предварительно установленный возраст составляет от 60 до 70 лет. Предварительно установленный возраст в порядке возрастающего предпочтения составляет 60 лет, 61 год, 62 года, 63 года, 64 года, 66 лет, 67 лет, 68 лет, 69 лет, 70 лет и 65 лет и моложе. Характерно, что рацион с низким содержанием белков обеспечивает процент калорий, приходящихся на белки, который меньше предварительно установленного предельного уровня потребления белков. Как правило, предварительно установленный предельный уровень потребления белков составляет 20% калорий, приходящихся на белки, от суммарного среднего количества калорий, потребляемых в день субъектом. В одном варианте предварительно установленный предельный уровень потребления белков составляет 15% калорий, приходящихся на белки, от суммарного среднего количества калорий, потребляемых в день субъектом. В одном варианте предварительно установленный предельный уровень потребления белков составляет 10% калорий, приходящихся на белки, от суммарного среднего количества калорий, потребляемых в день субъектом. В другом варианте предварительно установленный предельный уровень потребления белков составляет 5% калорий, приходящихся на белки, от суммарного среднего количества калорий, потребляемых субъектом в день. В некоторых вариантах рацион с низким содержанием белков обеспечивает в порядке возрастающего предпочтения более 40%, 50%, 60%, 70%, 80% и 90% калорий, приходящихся на белки из источников растительного происхождения, таких как соевые бобы. Преимущественно, рацион с низким содержанием белков обеспечивает приблизительно 100% калорий, приходящихся на белки из источников растительного происхождения.
Уровни IGF-1 и/или IGFBP1 у субъекта контролируют для того, чтобы определить, какими должны быть периодичность и тип рациона для субъекта (например, смотри количество белков ниже), и конкретнее, необходимо ли увеличение или уменьшение потребления белков (т.е., как правило, уровни IGF-1 снижаются, а уровни IGFBP1 повышаются после обеспечения субъекта одним или несколькими циклами потребления рациона с низким содержанием белков). В частности, рацион с низким содержанием белков снижает уровни IGF-1 по меньшей мере на 10 процентов и/или повышает уровни IGFBP1 по меньшей мере на 50% процентов. В другом варианте рацион с низким содержанием белков снижает уровни IGF-I по меньшей мере на 20 процентов и/или повышает уровни IGFBP1 по меньшей мере на 75% процентов. В другом варианте рацион с низким содержанием белков снижает уровни IGF-I по меньшей мере на 50 процентов и/или повышает уровни IGFBP1 по меньшей мере в 2 раза. Если определяют, что снижение уровней IGF-I и/или повышение уровней IGFBP1 является недостаточным, рацион с низким содержанием белков можно откорректировать таким образом, чтобы обеспечить еще более низкие количества калорий, приходящихся на источники белков.
В модификации варианта осуществления по настоящему изобретению, если возраст субъекта превышает предварительно установленный возраст, который предусматривает обеспечение субъекта рационом с высоким содержанием белков, субъекта обеспечивают рационом с высоким содержанием белков, при этом рацион с высоким содержанием белков характеризуется процентным содержанием калорий, приходящихся на белки, выше обеспечиваемого предварительно установленным предельным уровнем потребления белков.
В другом варианте субъекта обеспечивают рационом с низким содержанием белков в течение предварительно установленного количества дней. Например, субъекта обеспечивают рационом с низким содержанием белков в течение от 2 до 10 дней. В другом варианте субъекта обеспечивают рационом с низким содержанием белков в течение от 3 до 7 дней. Во многих случаях субъекта периодически обеспечивают рационом с низким содержанием белков. Периодичность, с которой субъекта обеспечивают рационом с низким содержанием белков, определяют по уровням инсулиновой резистентности, уровням глюкозы натощак, IGF-I, IGFBP1, ожирению, индексу массы тела, потере массы за предыдущие 10 лет, семейному анамнезу рака, семейному анамнезу диабета, семейному анамнезу ранней смертности у субъекта. Периодичность может быть от ежемесячной для субъектов с высокими уровнями IGF-I (например, выше 200 нг/мл), низкими уровнями IGFBP1 и/или инсулиновой резистентностью до одного раза в 3 месяца для субъектов с IGF-I от 120 до 200 нг/мл и отсутствием инсулиновой резистентности.
В одном варианте рацион с низким содержанием белков представляет собой рацион, имитирующий воздержание от пищи, который обеспечивает менее 10% калорий, приходящихся на белки, и/или при этом все белки имеют растительное происхождение, как изложено в международной заявке на патент США PCT/US13/66236; полное раскрытие данной заявки на патент включено в данный документ посредством ссылки. В частности, субъект принимает рацион с низким содержанием белков в течение первого периода времени. Иногда используемый в данном документе рацион с низким содержанием белков по данному варианту осуществления). В одном варианте рацион с низким содержанием белков обеспечивает от 4,5 до 7 килокалорий на фунт массы тела субъекта в первый день (день 1), а затем от 3 до 5 килокалорий на фунт массы тела субъекта в день со второго по пятый день (дни 2-5) потребления рациона с низким содержанием белков. Субъект принимает второй рацион в течение второго периода времени. В одном варианте второй рацион обеспечивает общее потребление калорий в пределах 20 процентов от нормального потребления субъектом калорий, в течение 25-26 дней (например, сразу же) после рациона с низким содержанием белков. Обычно наблюдается, что уровень IGF-I снижается, а уровень IGFBP1 повышается. В одном варианте способ по данному варианту осуществления повторяют от 1 до 5 раз. В другом варианте способ по данному варианту осуществления повторяют от 2 до 3 раз. В еще одном варианте способ по данному варианту осуществления повторяют в течение ряда лет или в течение всей жизни субъекта с периодичностью каждые 1-3 месяца в зависимости от уровней IGF-I и IGFBP1 у субъекта, а также потребления белков. Периодичность может быть от ежемесячной для субъектов с высоким потреблением белков (более 15% калорий, приходящихся на белки), и/или с высокими уровнями IGF-I (например, выше 200 нг/мл) и низкими уровнями IGFBP1, и/или с инсулиновой резистентностью до одного раза в 3 месяца для субъектов с потреблением белков, составляющим 10-15% калорий, с уровнями IGF-I от 120 до 200 нг/мл и отсутствием инсулиновой резистентности.
В другом варианте комбинация рациона с низким содержанием белков и второго рациона (например, нормального рациона субъекта и нормального количества потребляемых калорий) обеспечивает субъекта суммарным количеством калорий в пределах 10 процентов от нормального количества потребляемых субъектом калорий. В другом варианте комбинация рациона с низким содержанием белков и второго рациона обеспечивает субъекта суммарным количеством калорий в пределах 5 процентов от нормального количества потребляемых субъектом калорий. В еще одном варианте комбинация рациона с низким содержанием белков и второго рациона обеспечивает субъекта суммарным количеством калорий в пределах 1 процента от нормального количества потребляемых субъектом калорий.
В одном варианте рацион, имитирующий воздержание от пищи (FMD), предусматривает полное замещение рациона субъекта в течение 5 дней. На протяжении 5-дневного периода субъект потребляет большое количество воды. Здоровые субъекты с нормальной массой тела (индекс массы тела или BMI от 18,5 до 25) потребляют рацион раз в месяц (5 дней на данном рационе и 25-26 дней на их нормальном рационе) в течение первых 3 месяцев и каждые 3 месяца после того (5 дней каждые 3 месяца). Измеряют массу тела субъекта, и субъект должен восстановить по меньшей мере 95% массы, потерянной во время потребления данного рациона, до начала следующего цикла. Субъектам с BMI менее 18,5 не следует принимать FMD без рекомендации и наблюдения врача. Такой же режим (раз в месяц в течение 3 месяцев, а затем один раз в 3 месяца) можно адаптировать для лечения или для поддержки лечения всех состояний, представленных в данной заявке на патент.
Руководства по потреблению для FMD включают данные пищевой ценности относительно калорий, макронутриентов и микронутриентов. Калории потребляют в зависимости от массы тела потребителя. Суммарное число потребляемых калорий составляет 4,5-7 калорий на фунт (или 10-16 калорий на килограмм) в день 1 и 3-5 калорий на фунт (или 7-11 калорий на килограмм) во 2-5 дни. На фигурах 1-3 приведены перечни нутриентов для дней с первого по пятый. Помимо макроэлементов рацион должен содержать менее 30 г сахара в день 1 и менее 20 г сахара в дни 2-5. Рацион должен содержать менее 28 г белков в день 1 и менее 18 г белков в дни 2-5, преимущественно или исключительно из источников растительного происхождения. Рацион должен содержать от 20 до 30 граммов мононенасыщенных жиров в день 1 и 10-15 граммов мононенасыщенных жиров в дни 2-5. Рацион должен содержать от 6 до 10 граммов полиненасыщенных жиров в день 1 и 3-5 граммов полиненасыщенных жиров в дни 2-5. Рацион должен содержать от менее 12 г насыщенных жиров в день 1 и менее 6 граммов насыщенных жиров в дни 2-5. Как правило, жиры во все дни получают из комбинации следующих: миндаля, орехов макадамии, орехов пекан, кокосового ореха, кокосового масла, оливкового масла и льняного семени. В одном варианте рацион FMD включает более 50% от рекомендованной суточной нормы пищевых волокон во все дни. В дополнительном варианте количество пищевых волокон составляет более 15 граммов в день во все пять дней. Рацион должен содержать 12-25 граммов глицерина в день для дней 2-5. В одном варианте глицерин обеспечивают в количестве 0,1 грамма на фунт массы тела/день.
В одном варианте FMD включает следующие микронутриенты (по меньшей мере 95% из источников неживотного происхождения): более 5000 ME в день (дни 1-5) витамина А; 60-240 мг в день (дни 1 -5) витамина С; 400-800 мг в день (дни 1 -5) кальция; 7,2-14,4 мг в день (дни 1-5) железа; 200-400 мг в день (дни 1-5) магния; 1-2 мг в день (дни 1-5) меди; 1-2 мг в день (дни 1-5) марганца; 3,5-7 мкг в день (дни 1-5) селена; 2-4 мг в день (дни 1-5) витамина В1; 2-4 мг в день (дни 1-5) витамина В2; 20-30 мг в день (дни 1-5) витамина В3;1-1,5 мгв день (дни 1 -5) витамина В5; 2-4 мг в день (дни 1-5) витамина В6; 240-480 мкг в день (дни 1-5) витамина В9; 600-1000 МЕД в день (дни 1-5) витамина D; 14-30 мг в день (дни 1-5) витамина Е; более 80 мкг в день (дни 1-5) витамина К; 16-25 мкг витамина В12 обеспечивают на протяжении всего 5-дневного периода; 600 мг докозагексаеновой кислоты (DHA, полученной из водорослей) обеспечивают на протяжении всего 5-дневного периода. Рацион FMD обеспечивает высокий уровень содержания микронутриентов (т.е. более 50 процентов по массе), полученных преимущественно из природных источников: капусты кормовой, орехов кешью, перца желтого стручкового, лука, лимонного сока, дрожжей, куркумы, грибов, моркови, оливкового масла, свекольного сока, шпината, томата, капусты листовой, крапивы, тимьяна, соли, перца, витамина В12 (цианокобаламина), разновидностей свеклы, тыквы мускатной, капусты листовой, томата, орегано, томатного сока, апельсинового сока, сельдерея корневого, салата ромэн, шпината, тмина, кожуры апельсина, лимонной кислоты, мускатного ореха, гвоздики и их комбинаций. В таблице 1 представлен пример дополнения микронутриентами, которые могут быть обеспечены в рационе FMD.


В другом варианте осуществления обеспечивают комплект рациона для реализуемой схемы рациона, изложенной выше. Комплект рациона включает первый набор порций для рациона с низким содержанием белков, принимаемых субъектом в течение первого периода времени, при этом рацион с низким содержанием белков обеспечивает от 4,5 до 7 килокалорий на фунт массы тела субъекта в первый день и от 3 до 5 килокалорий на фунт массы тела субъекта в день со второго по пятый день потребления рациона с низким содержанием белков. Комплект рациона включает порции, обеспечивающие менее 30 г сахара в первый день; менее 20 г сахара со второго по пятый дни; менее 28 г белков в первый день; менее 18 г белков в день со второго по пятый дни; от 20 до 30 граммов мононенасыщенных жиров в первый день; от 10 до 15 граммов мононенасыщенных жиров со второго по пятый дни; от 6 до 10 граммов полиненасыщенных жиров в первый день; от 3 до 5 граммов полиненасыщенных жиров со второго по пятый дни; менее 12 г насыщенных жиров в первый день; менее 6 граммов насыщенных жиров со второго по пятый дни и от 12 до 25 граммов глицерина в день со второго по пятый дни. В одном варианте комплект рациона дополнительно включает порции, достаточные для обеспечения приведенными выше микронутриентами. В дополнительном варианте комплект рациона обеспечен инструкциями, предоставляющими подробное описание изложенных выше способов.
В одном варианте изложенных выше вариантов осуществления продовольствие для 5-дневного рациона включает: супы/бульоны, безалкогольные напитки, ореховые батончики и пищевые добавки. Рацион принимают следующим образом: 1) в первый день обеспечивают рацион питательностью 1000-1200 ккал с высоким содержанием микронутриентов; 2) в течение следующих 4 дней обеспечивают суточный рацион питательностью 650-800 ккал плюс напиток, который содержит заменитель глюкозы в качестве источника углерода (глицерин или аналог), обеспечивающий от 60 до 120 ккал. Заменитель источника углерода не препятствует влиянию воздержания от пищи на активацию стволовых клеток.
В другом варианте изложенных выше вариантов осуществления 6-дневная схема рациона с низким содержанием белков включает: супы/бульоны, безалкогольные напитки, ореховые батончики и пищевые добавки. Рацион принимают следующим образом: 1) в первый день обеспечивают рацион питательностью 1000-1200 ккал с высоким содержанием микронутриентов; 2) в течение следующих 3 дней обеспечивают суточный рацион питательностью менее 200 ккал плюс напиток, который содержит заменитель глюкозы в качестве источника углерода (глицерин или аналог), обеспечивающий от 60 до 120 ккал. Данный заменитель источника углерода, в том числе глицерин, не препятствует влиянию воздержания от пищи на активацию стволовых клеток; 3) в 5-й день субъект потребляет нормальный рацион; и 4) в 6-й день дополнительно к нормальному рациону обеспечивают добавочные восполняющие пищевые продукты, состоящие из источника с высоким содержанием жиров питательностью 300 ккал и питательной смеси микронутриентов, в 6-й день обеспечивают восполняющие пищевые продукты, состоящие из источника с высоким содержанием жиров питательностью 300 ккал и питательной смеси микронутриентов.
В еще одном варианте схема рациона включает: продовольствие для 6-дневного рациона с низким содержанием белков, включающее: супы/бульоны, безалкогольные напитки, ореховые батончики и пищевые добавки. 1) В первые день обеспечивают рацион питательностью 1000-1200 ккал с высоким содержанием микронутриентов; 2) в течение следующих 3 дней суточный рацион питательностью от 600 до 800 ккал, который содержит менее 10 граммов белка и менее 200 ккал, приходящихся на сахара; 3) в 5-й день субъект получает нормальный рацион; и 4) в 6-й день дополнительно к нормальному рациону обеспечивают добавочные восполняющие пищевые продукты, состоящие из источника с высоким содержанием жиров питательностью 300 ккал и питательной смеси микронутриентов, в 6-й день обеспечивают добавочные восполняющие пищевые продукты, состоящие из источника с высоким содержанием жиров питательностью 300 ккал и питательной смеси микронутриентов.
Особенно предпочтительная схема рациона и комплекты пищевых рационов приведены в публикации WIPO № WO 2011/050302, а схемы питания - в данном документе. Публикация WIPO № WO 2011/050302 включена в данный документ посредством ссылки во всей своей полноте. В частности, субъектов обеспечивают рационом с низким содержанием белков в течение первого периода времени, вторым рационом в течение второго периода времени и необязательно третьим рационом в течение третьего периода времени. Рацион с низким содержанием белков обеспечивает субъекта не более чем 50% от нормального количества потребляемых субъектом калорий, при этом по меньшей мере 50% килокалорий приходится на жиры, предпочтительно мононенасыщенные жиры. Нормальное количество потребляемых субъектом калорий представляет собой число ккал, которое расходует субъект для поддержания своей массы. Нормальное количество потребляемых субъектом калорий можно оценить, проведя опрос субъекта или исходя из массы тела субъекта. Нормальное количество потребляемых субъектом калорий ориентировочно составляет в среднем 2600 ккал/день для мужчин и 1850 ккал/день для женщин. В определенных случаях рацион с низким содержанием белков обеспечивает субъекта 700-1200 ккал/день. В особенно применимом варианте рацион с низким содержанием белков обеспечивает мужчину-субъекта со средней массой тела приблизительно 1100 ккал/день, а женщину-субъекта со средней массой тела - 900 ккал/день. Как правило, первый предварительно установленный период времени составляет от приблизительно 1 до 5 дней. В конкретных примерах первый предварительно установленный период времени составляет 1 день. Для объективной оценки уровня жиров в рационе с низким содержанием белков Управление по санитарному надзору за качеством пищевых продуктов и медикаментов рекомендует следующее распределение питательных веществ для типичного суточного рациона питательностью 2000 килокалорий: 65 граммов жиров (приблизительно 585 килокалорий), 50 граммов белков (приблизительно 200 килокалорий), 300 граммов общих углеводов (приблизительно 1200 килокалорий). Таким образом, в одном варианте рациона с низким содержанием белков большая часть калорий, приходящихся на углеводы и белки, устранена.
Несмотря на то, что рацион с низким содержанием белков, по сути, содержит любой источник жиров, особенно применимыми являются источники с высоким содержанием ненасыщенных жиров, в том числе источники мононенасыщенных и полиненасыщенных жиров (например, омега-3/6 незаменимых жирных кислот). Подходящие примеры пищевых источников мононенасыщенных жиров включают без ограничения арахисовое масло, маслины, орехи (например, миндаль, пекан, фисташки, кешью), авокадо, семена (например, кунжута), масла (например, оливковое, кунжутное, арахисовое, каноловое) и т.д. Подходящие примеры пищевых источников полиненасыщенных жиров включают без ограничения грецкие орехи, семена (например, тыквы, подсолнуха), льняное семя, рыбу (например, лосось, тунец, скумбрия), масла (например, сафлоровое, соевое, кукурузное). Рацион с низким содержанием белков включает компонент, выбранный из группы, состоящей из растительных экстрактов, минеральных веществ, омега-3/6 незаменимых жирных кислот и их комбинаций. В одном варианте такой растительный экстракт обеспечивает эквивалент 5 рекомендованных суточных порций овощей. Подходящие источники растительных экстрактов включают без ограничения капусту китайскую, капусту кормовую, латук, спаржу, морковь, тыкву мускатную, люцерну, горох огородный, томат, капусту, капусту цветную, разновидности свеклы. Подходящие источники омега-3/6 незаменимых жирных кислот включают рыбу, такую как лосось, тунец, скумбрия, луфарь, меч-рыба и им подобные. В дополнительном варианте рацион с низким содержанием белков включает источники жиров, такие что по меньшей мере 25 процентов калорий, приходящихся на жиры, обеспечиваются короткоцепочечными жирными кислотами, содержащими от 2 до 7 атомов углерода, и/или среднецепочечными насыщенными жирными кислотами, содержащими от 8 до 12 атомов углерода. Конкретные примеры жирных кислот включают лауриновую и/или миристиновую кислоту, а источники жиров включают оливковое масло, масло из косточек и/или кокосовое масло. В другом варианте рацион с низким содержанием белков включает калории, приходящиеся на жиры, в количестве от приблизительно 0 до 22 процентов от суммарного количества калорий, содержащегося в рационе.
В одном варианте субъекта затем обеспечивают вторым рационом в течение второго периода времени. Второй рацион обеспечивает субъекта не более чем 900 ккал/день. В определенных случаях второй рацион обеспечивает субъекта не более чем 200 ккал/день. Как правило, второй предварительно установленный период времени составляет от приблизительно 2 до 7 дней. В отдельных конкретных случаях второй предварительно установленный период времени составляет 3 дня. В еще одном варианте второй рацион включает компонент, выбранный из группы, состоящей из растительных экстрактов, минеральных веществ, омега-3/6 незаменимых жирных кислот и их комбинаций. В одном варианте такой растительный экстракт обеспечивает эквивалент 5 рекомендованных суточных порций овощей. Подходящие источники растительных экстрактов включают без ограничения капусту китайскую, капусту кормовую, латук, спаржу, морковь, тыкву мускатную, люцерну, горох огородный, томат, капусту, капусту цветную, виды свеклы. Подходящие источники омега-3/6 незаменимых жирных кислот включают рыбий жир из лосося, тунца, скумбрии, луфаря, меч-рыбы и им подобных.
Эффективность схем питания в данном документе контролируют путем измерения у субъекта ряда параметров. Например, желательно, чтобы концентрация IGF-I в сыворотке крови субъекта снижалась на 25-90% к завершению второго периода потребления рациона в зависимости от исходного уровня IGF-I и потребления белка и от уровней, оптимальных для защитного действия в отношении смертности, описанных в прилагаемых публикациях. Также желательно, чтобы концентрация глюкозы в крови субъекта снижалась на 25-75% к завершению второго периода потребления рациона.
В другой модификации варианта осуществления настоящего изобретения рацион с низким содержанием белков включает пищевую добавку с определенными аминокислотами, содержащую соответствующие аминокислоты. Как правило, пищевая добавка, обеспечивает избыточные уровни заменимых аминокислот для потребления в течение периода 5-7 дней вместе с очень низкими количествами белков или безбелковым рационом. В одном варианте рацион с низким содержанием белков чередуют с рационом с нормальным содержанием белков. В таких вариантах рацион с низким содержанием белков обеспечивают в течении 7 дней каждые 2 недели - 2 месяца с нормальным рационом в течение от 1 до 7 недель между ними. В одном варианте использование пищевой добавки с определенными аминокислотами фактически исключает из рациона следующие аминокислоты: изолейцин, лейцин, лизин, метионин, фенилаланин, треонин, триптофан, валин и аргинин. В данном контексте "фактически исключает" означает, что общее количество исключенных аминокислот составляет в порядке возрастания предпочтения менее 5 процентов по массе, 3 процентов по массе, 1 процента по массе и 0,5 процента по массе. Взамен, рацион с определенными аминокислотами в качестве источника азота обеспечивает одну или несколько из следующих аминокислот: аланина, аспарагиновой кислоты, цистеина, глутаминовой кислоты, глицина, гистидина, пролина, серина и тирозина. В таблицах 2-4 представлены характеристики рациона с определенными аминокислотами для мыши, который также ограничен по содержанию белков, как указано ниже. Типичный рацион для мыши обеспечивает приблизительно 19 ккал в день. Для других млекопитающих, таких как люди, рацион с ограниченным содержанием белков (PR) рассчитан таким образом, чтобы обеспечить необходимое количество калорий. Например, типичное количество потребляемых калорий для взрослых в Соединенных Штатах составляет приблизительно 2200 калорий в день. В таблице 5 представлены количества килокалорий в день из каждого источника для людей-субъектов, тогда как в таблице 6 представлены количества в граммах в день из каждого источника для людей.





В одном варианте килограмм рациона для мыши с определенными аминокислотами включает от приблизительно 2 г до 20 г аланина, от 10 г до 30 г аспарагиновой кислоты, от 2 г до 20 г цистеина, от 40 г до 80 г глутаминовой кислоты, от 2 г до 20 г глицина, от 2 г до 20 г гистидина, от 15 г до 50 г пролина, от 5 г до 30 г серина и от 5 до 30 г тирозина. В отношении людей-субъектов данные диапазоны умножают на коэффициент (т.е. приблизительно 0,572) для получения структуры состава рациона на день для людей-субъектов. Например, суточные количества указанных аминокислот для людей (рацион, обеспечивающий 2200 калорий/день) в рационе с определенными аминокислотами составляют от приблизительно 2 до 12 г аланина, от 5 г до 30 г аспарагиновой кислоты, от 1 г до 7 г цистеина, от 18 г до 73 г глутаминовой кислоты, от 2 г до 9 г глицина, от 2 г до 10 г гистидина, от 9 г до 37 г пролина, от 5 г до 21 г серина и от 5 до 21 г тирозина. В другом варианте рацион с определенными аминокислотами включает от приблизительно 160 до приблизительно 240 г указанных аминокислот на килограмм рациона. Таким образом, для людей рацион с определенными аминокислотами обеспечивает от приблизительно 80 до 160 г указанных аминокислот в день, при этом используется коэффициент (0,572) для пересчета значения на килограмм рациона в значение, характерное для рациона человека, составляющее приблизительно 2200 калорий/день. В другом варианте рацион с определенными аминокислотами включает по меньшей мере 6 аминокислот, выбранных из группы, состоящей из аланина, аспарагиновой кислоты, цистеина, глутаминовой кислоты, глицина, гистидина, пролина, серина и тирозина, в изложенных выше количествах. В еще одном варианте рацион с определенными аминокислотами обеспечивает количества аминокислот в граммах на кг массы тела человека в день, изложенные в таблице 7. В частности, рацион с определенными аминокислотами обеспечивает следующие количества в граммах на кг массы тела человека в день: 0,06 г аланина, 0,14 г аспарагиновой кислоты, 0,04 г цистеина, 0,45 г глутаминовой кислоты, 0,05 г глицина, 0,06 г гистидина, 0,23 г пролина, 0,13 серина и 0,13 г тирозина. В другом варианте количество каждой из данных аминокислот находится в пределах диапазона плюс или минус 30 процентов от указанной величины.


В другом варианте способ включает стадию приема субъектом рациона с ограниченным содержанием белков (PR) в течение первого периода времени. В одном варианте рацион с низким содержанием белков включает пищевую добавку с определенными аминокислотами. В одном варианте первый период времени составляет от приблизительно 5 дней до 14 дней, при этом 7 дней являются типичными. Более того, рацион с низким содержанием белков обеспечивает субъекта 70-100 процентами от нормального количества потребляемых субъектом калорий. Рацион с низким содержанием белков в качестве источника азота включает фактически только аминокислоты. Например, в рационе с ограниченным содержанием белков менее 10 процентов калорий приходится на белки. В другом варианте в рационе с ограниченным содержанием белков менее 5 процентов калорий приходится на белки. В другом варианте в рационе с ограниченным содержанием белков ноль процентов калорий приходится на белки. В частности, в рационе с ограниченным содержанием белков фактически исключены следующие аминокислоты: изолейцин, лейцин, лизин, метионин, фенилаланин, треонин, триптофан, валин и аргинин. В данном контексте "фактически исключает" означает, что общее количество исключенных аминокислот составляет в порядке возрастания предпочтения менее 5 процентов по весу, 3 процентов по весу, 1 процента по весу и 0,5 процента по весу. Взамен, рацион с ограниченным содержанием белков в качестве источника азота обеспечивает одну или несколько из следующих аминокислот: аланина, аспарагиновой кислоты, цистеина, глутаминовой кислоты, глицина, гистидина, пролина, серина и тирозина. В таблицах 2-4 приведены характеристики рациона с ограниченным содержанием белков, включающего пищевую добавку, для исследований на мышах, которые приведены ниже. Типичный рацион для мыши обеспечивает приблизительно 19 ккал в день. Для других млекопитающих, таких как люди, рацион с низким содержанием белков рассчитан таким образом, чтобы обеспечить необходимое количество калорий. Например, типичное количество потребляемых калорий для взрослых в Соединенных Штата составляет приблизительно 2200 килокалорий в день. В таблице 5 представлены количества килокалорий в день из каждого источника для людей-субъектов, тогда как в таблице 6 представлены количества в граммах в день из каждого источника для людей.
В одном варианте количества аминокислот в килограмме рациона с низким содержанием белков для мышей представлены в таблице 8. В одном варианте килограмм рациона с низким содержанием белков для мышей включает от приблизительно 2 г до 20 г аланина, от 10 г до 30 г аспарагиновой кислоты, от 2 г до 20 г цистеина, от 40 г до 80 г глутаминовой кислоты, от 2 г до 20 г глицина, от 2 г до 20 г гистидина, от 15 г до 50 г пролина, от 5 г до 30 г серина и от 5 до 30 г тирозина. В отношении людей-субъектов данные диапазоны умножают на коэффициент (т.е. приблизительно 0,572) для получения суточной потребности людей-субъектов в данных аминокислотах в день. Например, для людей суточные количества указанных аминокислот (рацион, обеспечивающий 2200 калорий/день) в рационе с низким содержанием белков составляют от приблизительно 2 до 12 г аланина, от 5 г до 30 г аспарагиновой кислоты, от 1 г до 7 г цистеина, от 18 г до 73 г глутаминовой кислоты, от 2 г до 9 г глицина, от 2 г до 10 г гистидина, от 9 г до 37 г пролина, от 5 г до 21 г серина и от 5 до 21 г тирозина. В другом варианте рацион с ограниченным содержанием белков включает от приблизительно 160 до приблизительно 240 г указанных аминокислот на килограмм рациона. Таким образом, для людей рацион с низким содержанием белков обеспечивает от приблизительно 80 до 160 г указанных аминокислот в день, при этом используется коэффициент (0,572) для пересчета значения на килограмм рациона в значение, характерное для рациона человека, составляющее приблизительно 2200 калорий/день. В другом варианте рацион с ограниченным содержанием белков включает по меньшей мере 6 аминокислот, выбранных из группы, состоящей из аланина, аспарагиновой кислоты, цистеина, глутаминовой кислоты, глицина, гистидина, пролина, серина и тирозина, в изложенных выше количествах. В таблице 8 представлен пример содержания аминокислот в рационе с ограниченным содержанием белков для рациона для мышей В таблице 8 также представлен коэффициент, выражающий отношение количества указанных аминокислот в рационе с ограниченным содержанием белков к таковому в контроле (нормальном рационе). Данные отношения в одинаковой степени применимы по отношению к другим млекопитающим, таким как люди-субъекты. В еще одном варианте рацион с низким содержанием белков обеспечивает количества аминокислот в граммах на кг массы тела человека в день, изложенные в таблице 8. В частности, рацион РК обеспечивает следующие количества в граммах на кг массы тела человека в день: 0,06 г аланина, 0,14 г аспарагиновой кислоты, 0,04 г цистеина, 0,45 г глутаминовой кислоты, 0,05 г глицина, 0,06 г гистидина, 0,23 г пролина, 0,13 серина и 0,13 г тирозина. В другом варианте количество каждой из данных аминокислот находится в пределах диапазона плюс или минус 30 процентов от указанной величины.
В другом варианте обеспечивают способ снижения у субъекта уровней глюкозы и/или IGF-1. Способ включает стадию обеспечения субъекта рационом с низким содержанием белков, содержащим менее приблизительно 10 процентов калорий, приходящихся на источники белков. Уровни глюкозы и/или IGF-1 у субъекта контролируют для определения необходимости увеличения или уменьшения потребления белков. В одном варианте рацион с низким содержанием белков включает от 0 до 10 процентов калорий, приходящихся на источники белков. В дополнительном варианте рацион с низким содержанием белков включает от 0 до 5 процентов калорий, приходящихся на источники белков. В другом варианте рацион с низким содержанием белков, как правило, содержит приблизительно 0 процентов калорий, приходящихся на источники белков. В другом варианте рацион с низким содержанием белков также представляет собой низкокалорийный рацион, который включает источники жиров, такие что по меньшей мере 50 процентов калорий, приходящихся на жиры, обеспечиваются длинноцепочечными ненасыщенными жирными кислотами, приведенными выше, содержащими от 13 до 28 атомов углерода. Типичные источники жиров включают растительное масло, такое как соевое масло. В дополнительном варианте рацион с низким содержанием белков включает источники жиров, такие что по меньшей мере 25 процентов калорий, приходящихся на жиры, обеспечиваются короткоцепочечными жирными кислотами, содержащими от 2 до 7 атомов углерода, и/или среднецепочечными насыщенными жирными кислотами, содержащими от 8 до 12 атомов углерода. Конкретные примеры жирных кислот включают лауриновую и/или миристиновую кислоту, а источники жиров включают оливковое масло, масло из косточек и/или кокосовое масло. В другом варианте рацион с низким содержанием белков включает калории, приходящиеся на жиры, в количестве от приблизительно 0 до 22 процентов от суммарного количества калорий, содержащегося в рационе.
В другом варианте осуществления предлагается способ ослабления симптома химической интоксикации у субъекта. Способ включает стадию обеспечения рационом с низким содержанием белков в течение первого периода времени, при этом рацион с низким содержанием белков включает менее 10 процентов калорий, приходящихся на белки. Субъекта обеспечивают рационом с ограниченной калорийностью в течение второго периода времени, при этом рацион с ограниченной калорийностью содержит от 0 до 50% калорий от их количества в рационе с низким содержанием белков. В одном варианте рацион с ограниченной калорийностью включает от 0 до 10% калорий, приходящихся на источники белков. В одном варианте субъекта подвергают лечению химиотерапевтическими средствами. Примеры химотерапевтических средств включают без ограничения доксорубицин, циклофосфамид, цисплатин, 5-фторурацил и их комбинации.
Следующие примеры иллюстрируют различные варианты осуществления настоящего изобретения. Специалистам в данной области будут очевидны многие варианты в пределах сущности и объема формулы настоящего изобретения.
Эксперименты с низким потреблением белка
Объединяли эпидемиологическое исследование в США в отношении 6381 мужчины и женщины, достигших возраста 50 лет и выше, из NHANES, единственное общенациональное репрезентативное исследование пищевого рациона в США с исследованиями на мышах и клетках для понимания связи между уровнем потребления и источником белков и аминокислот, старением, заболеваниями и смертностью.
Результаты
Популяция людей
Исследуемая популяция включала 6381 взрослого, достигшего возраста 50 лет и выше, из NHANES III, общенационального репрезентативного одномоментного срезового углубленного исследования. Средний возраст в исследуемой выборке составлял 65 лет, и она являлась репрезентативной для населения США по этнической принадлежности, образованию и показателям здоровья (таблица 13).
В среднем субъекты потребляли 1823 калории, из которых основная часть приходилась на углеводы (51%), затем на жиры (33%) и белки (16%) - при этом 11% белков животного происхождения. Процент калорий, приходящихся на белки, использовали для разделения субъектов на группу с высоким потреблениям белков (20% калорий или более, приходящихся на белки), группу с умеренным потреблением белков (10-19% калорий, приходящихся на белки) и группу с низким потреблением белков (менее 10% калорий, приходящихся на белки).
Последующее наблюдение в отношении показателей смертности было доступно для всех участников NHANES посредством связи с Национальным реестром смертности вплоть до 2006 г. (22). В нем были предоставлены время и причина смерти. Период последующего наблюдения в отношении смертности охватывал всего 83308 человеко-лет старше 18 лет при 40% общей смертности, 19% смертности от сердечно-сосудистых заболеваний (CVD), 10% смертности от рака и приблизительно 1% смертности, вызванной диабетом.
Взаимосвязь между потреблением белков и смертностью
Используя модели пропорциональных рисков Кокса, выявили, что существует прямая взаимосвязь между высоким и умеренным потреблением белков и показателями смертности, связанной с диабетом, но она отсутствует для показателей смертности по всем причинам, смертности от CVD или от рака при оценке всех субъектов в возрасте старше 50 лет. Результаты показали, что участники групп как с высоким потреблением белков, так и с умеренным потреблением белков имели более высокие степени риска смертности от диабета по сравнению с участниками группы с низким потреблением белков. Хотя в совокупности данные результаты и указывают на то, что потребление белков от умеренного до высокого способствует смертности от диабета, требуются более масштабные исследования для дополнительной проверки этой вероятности. Альтернативное объяснение повышенной смертности от диабета в группе с высоким потреблением белков состоит в том, что после установления диагноза диабета, индивидуумы могу переходить на потребление рациона, предусматривающего более высокое содержание белков, низкое содержание жиров и низкое содержание углеводов. Для того, чтобы это проверить, у участников, среди которых в начале исследования диабет не был распространен, оценивали взаимосвязь между потреблением белков и смертностью от диабета (таблица 19).
Среди субъектов без диабета в начале исследования у субъектов в группе с высоким потреблением белков имело место 73-кратное повышение риска (HR: 73,52; 95% CI: 4,47-1209,7), тогда как у категории с умеренным потреблением белков имело место 23-кратное повышение риска смертности от диабета (HR: 22,93; 95% CI: 1,31-400,7). Подчеркиваем, что наши соотношения рисков и доверительных интервалов могут быть завышены из-за нашего объема выборки и чрезвычайно низкой частоты смертности от диабета в группе с низким потреблением белков. Всего среди лиц без диабета в начале исследования зарегистрировали только 21 случай смерти от диабета -из которых только 1 был в группе с низким потреблением белков. Однако несмотря на малый объем выборки, наши результаты тем не менее демонстрируют значимые взаимосвязи между повышенным потреблением белков и смертностью, связанной с диабетом.
С помощью моделей пропорциональных рисков Кокса проводили повторное исследование в отношении взаимосвязи между потреблением белков и возрастом, чтобы определить, отличается ли взаимосвязь между потреблением белков и смертностью у лиц среднего возраста и пожилого возраста. Были выявлены значимые взаимосвязи как в отношении смертности по всем причинам, так и смертности от рака, демонстрируя, что низкое потребление белков являлось предпочтительным в зрелом возрасте; однако с возрастом эти преимущества уменьшались (фиг. 4). Исходя из данных результатов, данную выборку стратифицировали на две возрастные группы - в возрасте 50-65 лет (n=3039) и в возрасте 66+ лет (n=3342) и повторно исследовали взаимосвязь между потреблением белков и смертностью от конкретных причин. Среди достигших возраста 50-65 лет повышенные уровни потребления белков были связанны со значительным повышением рисков смертности по всем причинам и смертности от рака (таблица 12). В данном возрастном диапазоне у субъектов в группе с высоким потреблением белков имело место 74% повышение относительного риска смертности по всем причинам (HR: 1,74; 95% CI: 1,02-2,97) и в 4 раза большая вероятность умереть от рака (HR: 4,33; 95% CI: 1,96-9,56) по сравнению с субъектами из группы с низким потреблением белков. Ни одна из данных взаимосвязей не зависела в значительной степени от контроля процентов калорий, приходящихся на общее количество жиров, или процента калорий, приходящихся на общее количество углеводов. Однако при контроле процента калорий, приходящихся на белки животного происхождения, взаимосвязь между общим количеством потребляемых белков и смертностью по всем причинам и смертностью от рака устранялась или значительно уменьшалась, указывая, соответственно, на то, что белки животного происхождения опосредуют значительную часть данных зависимостей. При осуществлении контроля в отношении влияния белков растительного происхождения изменение взаимосвязи между потреблением белков и смертностью отсутствовало, указывая на то, что потребление высоких уровней белков животного происхождения способствует смертности, и что белки растительного происхождения не обладают защитным эффектом (таблица 17).
По сравнению с субъектами, потребляющими рацион с низким содержанием белков, у субъектов, потреблявших умеренные уровни белков, также имело место 3-кратное повышение уровня смертности от рака (HR: 3,06; 95% CI: 1,49-6,25), что не было обусловлено либо процентом калорий, приходящихся на жиры, либо процентом калорий, приходящихся на углеводы, но незначительно уменьшалось при контроле процента калорий, приходящихся на белки животного происхождения (HR: 2,71; 95% CI: 1,24-5,91). Хотя величина эффекта не была столь весомой, как для субъектов в группе с высоким потреблением белков. Вместе взятые, данные результаты указывают на то, что у респондентов, достигших возраста 50-65 лет, потребляющих высокие уровни белков животного происхождения, наблюдается значительное увеличение рисков в отношении общей смертности и смертности от рака, в то же время, риски могут быть несколько снижены, если белок не поступает из источника животного происхождения. Подобные результаты были получены при анализе популяции в возрасте 45-65 лет (данные не показаны).
В противоположность результатам, приведенным выше, среди респондентов в возрасте 66 лет и выше в начале исследования повышенные уровни потребления белков были взаимосвязаны с противоположным эффектом в отношении общей смертности и смертности от рака, но с подобным эффектом в отношении смертности от диабета (таблица 12). По сравнению с субъектами, потреблявшими рацион с низким содержанием белков, у субъектов, потреблявших высокие количества белков, также имело место 28% снижение смертности по всем причинам (HR: 0,72; 95% CI: 0,55-0,94), тогда как у субъектов, потреблявших умеренные количества белков, наблюдалось 21% снижение смертности по всем причинам (HR: 0,78; 95% СТ. 0,62-0,99). Кроме того, на это не влиял процент калорий, приходящихся на жиры, углеводы или белки животного происхождения. У субъектов с высоким уровнем потребления белков также имело место 60% снижение смертности от рака (HR: 0,40; 95% CI: 0,23-0,71) по сравнению с теми, кто потреблял рацион с низким содержанием белков, на что также не влияло осуществление контроля потребления других нутриентов или источника белков.
Влияние IGF-1 на взаимосвязь между потреблением белков и смертностью
Имела место прямая взаимосвязь между скорректированными средними значениями уровней IGF-1 и потреблением белков для обеих возрастных групп (фиг. 5). Поскольку данные по IGF-1 были доступны только для произвольно выбранной подвыборки (n=2253), повторно исследовали зависящие от возраста взаимосвязи между потреблением белков и смертностью от конкретных причин в данной выборке и обнаружили, что они аналогичны наблюдавшимся в полной выборке; хотя с несколько большими значениями величины эффекта (таблица 15). Далее анализировали, выступает ли IGF-1 модификатором или промежуточной переменной во взаимосвязи между потреблением белков и смертностью. Выявили, что хотя IGF-1 не лежал в основе взаимосвязи между потреблением белков и смертностью (таблица 15), он являлся важным модификатором в данной взаимосвязи - на что указывают статистические значимые взаимосвязи между уровнем белков и IGF-1 (таблица 16).
На основе данных моделей рассчитывали предполагаемые соотношения рисков в зависимости от IGF-1 и белка в группе (фиг. 12). Результаты показали, что при сравнении группы высокого потребления белков и низкого потребления белков повышение IGF-1 на каждые 10 нг/мл повышает риск смертности от рака среди субъектов в возрасте 50-65 лет на дополнительные 9% (HRвысокоепотреблениебелков×IGF-1: 1,09; 95% CI: 1,01-1,17). Альтернативно, среди более пожилых субъектов (66+ лет) при сравнении с субъектами из группы с низким потреблением белка у субъектов, потреблявших рационы с высоким или умеренным количеством белков, имелся пониженный риск смертности от CVD, если IGF-1 был также низким; однако не было выявлено каких-либо преимуществ при повышении IGF-1.
Потребление белков, IGF-1 и раку мышей
Для проверки причин и дальнейшего исследования механизма изучали влияние потребления белков в диапазоне (4-18%), аналогичном таковому для субъектов в исследовании NHANES, на уровни циркулирующего в крови IGF-1, частоту возникновения рака и прогрессирование развития опухоли у грызунов. Самцов мышей C57BL/6 в возрасте 18 недель непрерывно в течение 39 дней кормили экспериментальными изокалорийными рационами, разработанными для обеспечения либо высокого (18%), либо низкого (7%) количества калорий, получаемых из белков, не применяя CR или не вызывая недостаточность питания (фиг. 9А, В).
Для того, чтобы понять, как различные уровни белков и IGF-1 у мышей могут воздействовать на способность новообразовавшейся опухоли продолжать существование и расти после одной недели потребления их соответствующих рационов, и первой, и второй группам подкожно имплантировали 20000 сингенных клеток меланомы мышей (В 16). Измерения опухолей начинали через 15 дней после имплантации при 22 днях потребления их соответствующих рационов, в этот момент частота возникновения заболевания составила 100% в группе с рационом с высоким содержанием белков и только 80% в группе с рационом с низким содержанием белков (фиг. 6А). В день 25 частота возникновения заболевания возрастала до 90% в группе с рационом с низким содержанием белков и оставалась на том же уровне до конца эксперимента (фиг. 6А). С 22 дня до конца эксперимента размер опухолей был существенно меньше в группе, потреблявшей меньшее количество белков, указывая на гораздо более медленное прогрессирование развития опухоли. В день 39 наблюдаемое среднее значение размера опухолей в группе с высоким потреблением белков было на 78% больше (в день 36 Р=0,0001; в день 39 Р<0,0001) (фиг. 6 В). Образцы крови отбирали и анализировали в день 16 для определения влияния потребления белков на IGF-1 и белок, ингибирующий IGF-1, IGFBP-1. Сывороточный IGF-1 был на 35% ниже (Р=0,0004) в группе с низким потреблением белков (4%) по сравнению с животными, которых кормили рационом с высоким содержанием белков (18%) (фиг. 6С). И наоборот, в группе с низким потреблением белков по сравнению с группой с высоким потреблением белков сывороточный IGFBP-1 был на 136% выше (Р=0,003) (фиг. 6D).
Для дополнительной проверки гипотезы, что ось GHR-IGF-1 способствует прогрессированию рака, мышам GHRKO с дефектом по GHR/IGF-1 и соответствующим им по возрасту и полу однопометным контролям (самцам мышей C57BL/6 в возрасте 18 недель) подкожно имплантировали меланому (В 16). Опухоли начали измерять через 10 дней после имплантации и продолжали до 18 дня. Данные показывают, что прогрессирование развития опухоли в значительной степени ингибировалось у мышей GHRKO по сравнению с прогрессированием у контрольной группы (фиг. 6Е; Р<0,01).
Также использовали мышиную модель рака молочной железы для проверки взаимосвязи между уровнями белков, частотой возникновения опухолей и их прогрессированием. Самкам мышей BAFB/c в возрасте 12 недель устанавливали тот же режим питания, который описан для мышей C57BL/6, за исключением того, что мыши должны были перейти с 4% на 7% ккал белковый рацион в течение первой недели для предупреждения потери массы (фиг. 9Е, F). После недели кормления данными рационами мышам подкожно имплантировали 20000 сингенных метастатических клеток рака молочной железы мышей (4Т1) и через 15 дней производили оценку в отношении опухолей. В 18 день после имплантации (25 день на рационе) частота возникновения опухолей составляла 100% в группе с высоким потреблением белков, но лишь 70% в группе с низким потреблением белков. К 39 дню частота возникновения заболевания в группе с низким потреблением белков возрастала до 80%, какой она оставалась до конца эксперимента (фиг. 6F). Данные прогрессировать развития опухоли также показывают, что группы на рационах с низким содержанием белков характеризовались меньшим средним значением размера опухолей. В группе с низким потреблением белков наблюдаемое среднее значение размера опухолей было на 45% меньше по сравнению с группой с высоким потреблением белков к 53 дню в конце эксперимента (Р=0,0038) (фиг. 6G). Также как и для мышей C57BL/6, измеряли IGF-1 на 16 день режима питания с ограничением белков. В группах с низким потреблением белков уровни IGF-1 снижались на 30% по сравнению с таковыми в группе с высоким потреблением белков (Р<0,0001) (фиг. 6Н). Кроме того, низкий уровень потребления белков также вызывал повышение уровня IGFBP1 на 84% (Р=0,001) (фиг. 61), аналогично тому, что наблюдали у мышей с генетическим фоном C57BL/6 (фиг. 6D). Аналогичным образом, когда уровень потребления соевых белков снижался с высоких уровней до низких уровней, наблюдали 30% снижение уровня IGF-1 (p<0,0001) (фиг. 6J) и 140% повышение уровня IGFBP-1 (p<0,0001) (фиг. 6К). Несмотря на наличие тенденции в отношении влияния замещения белков животного происхождения таким же уровнем белков растительного происхождения на IGF-I и IGFBP1, различия не были значимыми. Эти данные указывают на то, что более низкое потребление белков может играть роль в снижении частоты возникновения и/или прогрессирования рака отчасти путем снижения уровня IGF-1 и повышения уровня ингибитора IGF-1, IGFBP1. Дополнительные исследования с различными типами белков животного происхождения в сравнении с белками растительного происхождения необходимы для определения их влияния на IGF-I и IGFBP1.
Исследования на клетках
Для того, чтобы понять, существует ли принципиальная связь между уровнем аминокислот и сроком жизни, оценивали влияние присутствия определенных концентраций аминокислот на рост дрожжей и оценивали развитие при помощи анализов выживаемости и частоты возникновения мутаций. Штамм дикого типа DBY746 S. cerevisiae выращивали в присутствии половинной (0,5Х), стандартной (IX) и двойной концентрации (2Х) аминокислот, при этом концентрация всех других нутриентов оставалась постоянной. Выживаемость определяли в 1, 3, 5 и 8 дни. В 1 и 3 дни различий в выживаемости не наблюдали. В 5 день 2 наивысшие концентрации аминокислот показали тенденцию к повышенной смертности, результатом чего было 10-кратное снижение выживания клеток к 8 дню (фиг. 6L).
Для того, чтобы оценить взаимосвязь между аминокислотами, старением и зависимого от возраста повреждения ДНК, использовали старение S. cerevisiae для измерения частоты возникновения спонтанных мутаций. Частота возникновения мутаций была в 3 и 4 раза выше у 5-дневной культуры, но не у молодых клеток, которых подвергали воздействию аминокислот в концентрациях IX и 2Х, соответственно, по сравнении с клетками, которые подвергали воздействию аминокислот в концентрации 0,5Х (фиг. 6М). Данные результаты указывают на то, что даже у одноклеточных организмов аминокислоты способствуют клеточному старению и зависимой от возраста нестабильность генома.
Для дальнейшего определения путей, вовлечение которых способствует зависимой от возраста нестабильности генома, измеряли активацию экспрессии генов, реагирующих на стресс, которые регулируются путями Ras-PKA-Msn2-4 Tor-Sch9-Gisl, в присутствии или при отсутствии аминокислот. У клеток, выращенных в контрольной среде, содержащей только Trp, Leu и His (незаменимы для роста данного штамма), присутствие всех аминокислот в среде снижало активацию экспрессии генов, отвечающих за стрессоустойчивость, указывая на то, что добавление аминокислот было достаточным для подавления клеточной защиты (фиг. 6N).
Путь Tor-Sch9 увеличивает продолжительность жизни, но способствует возникновению мутаций в ДНК. Для того, чтобы определить, осуществляется ли регуляция зависимой от возраста нестабильности генома также при активации сигнального пути Ras-cAMP-PKA, исследовали ras2-дефектные мутанты. Подтвердили, что ras2Δ-мутанты являются долгоживущими (фиг. 11А), но и также показали, что инактивация сигнального пути Ras ослабляет зависимую от возраста и оксидативного стресса нестабильность генома (фиг. 11В, 11С, 6O,11D).
Вместе данные результаты указывают на механизм, где аминокислоты способны воздействовать на частоту возникновения мутаций и, таким образом, нестабильность геном по меньшей мере отчасти путем активации путей Tor-Sch9 и Ras/PKA и снижения стрессоустойчивости (фиг. 6Р).
Низкое потребление белков и поддержание массы у старых мышей
На основании наблюдаемых противоположных эффектов рациона с низким содержанием белков у субъектов в возрасте 50-65 лет в сравнении с субъектами в возрасте 65 лет и старше и значительного падения уровней BMI и IGF-1 после 65-летнего возраста предположили, что у более пожилых субъектов на рационе с низким содержанием белков может возникнуть истощение и неспособность к абсорбции или переработке достаточного уровня аминокислот. Для проверки данного возможного варианта у мышей молодых мышей (в возрасте 18 недель) и старых мышей (в возрасте 24 месяцев) кормили изокалорийными рационами, содержащими либо 18%, либо 4% белков животного происхождения. Рацион с очень низким содержанием белков специально выбрали с тем, чтобы выявить любую чувствительность к ограничению белков у старого организма. В то время как старые мыши, которых содержали на рационе с высоким содержанием белков в течение 30, прибавляли в массе, старые мыши, но не молодые, на рационе с низким содержанием белков к 15 дню потеряли 10% своей массы (фиг. 4А, В) в соответствии с теорией о влиянии старения, заключающемся в обращении благоприятных эффектов ограничения белков в отношении смертности в отрицательные эффекты.
Обсуждение
В данном документе при использовании крупного общенационального репрезентативного исследования питания населения в Соединенных Штатах в наших результатах показано, что среди тех, кто достиг возраста 45 и выше, уровень потребления белков связан с повышенным риском смертности от диабета, но не связан с различиями в показателях смертности по всем причинам, смертности от рака или от CVD. Тем не менее, обнаружили возрастную зависимость в отношении взаимосвязи между потреблением белков и смертностью, при этом субъекты в возрасте 50-65 лет получают преимущества от низкого потребления белков, а субъекты в возрасте 66+получают вред от рациона с низким содержанием белков - по меньшей мере в отношении общей смертности и рака. Это может объяснить, почему указанная в данном документе четкая взаимосвязь между потреблением белков, IGF-1 и смертностью не была описана ранее. Кроме того, среди 2253 субъектов, у которых измеряли уровни IGF-1, риски смертности по всем причинам и смертности от рака у субъектов с высоким потреблением белков при сравнении с группой с низким потреблением белков повышались, и в еще большей степени у тех, у кого также имелись высокие уровни IGF-1. Это соответствует предыдущим исследованиям, в которых исследовали взаимосвязь уровней IGF-1 с различными типами рака.
Причем имелись доказательства того, что тип потребляемого белка может быть важным. Наши результаты показали, что доля белков из источников животного происхождения в значительной мере обуславливала взаимосвязь между общим количеством потребляемого белка и смертностью по всем причинам, и смертностью от рака. Данные результаты соответствуют недавно полученным результатам исследований в отношении взаимосвязи между потреблением красного мяса и смерти по всем причинам, от CVD и от рака. В предыдущих исследованиях в США было выявлено, что низкоуглеводный рацион связан с повышением общей смертности, и когда такому рациону сопутствует повышенное потребление продуктов животного происхождения, риск общей смертности, а также смертности от CVD и от рака еще больше повышается. Однако наше исследование указывает на то, что влияние белков животного происхождения в отношении IGF-I, старения, диабета и рака может являться главным провоцирующим фактором смертности у людей в возрасте 45-65 лет на протяжении 18 лет после исследования, в котором устанавливали уровень потребления белка. К тому времени когорта, возраст которой на момент опроса составлял 65 лет, достигла бы возраста 83 лет, подчеркивая, что высокое потребление белков может способствовать смертности у субъектов старше 65 лет.
Наши результаты, полученные в экспериментах с дрожжами и мышами, могут также объяснить по меньшей мере часть принципиальной связи между потреблением белков, раком и общей смертностью путем нахождения связи между аминокислотами, стрессоустойчивостью, повреждением ДНК и частотой возникновения рака/прогрессированием рака. У мышей изменения, вызванные пониженными уровнями потребления белков, оказывали влияние, которое было достаточно сильным, чтобы предотвратить образование 10-30% опухолей, даже если в подкожном участке уже присутствовало 20000 опухолевых клеток. Более того, прогрессирование как меланомы, так и рака молочной железы в значительной степени подавлялось потреблением рациона с низким содержанием белков, указывая на то, что рационы с низким содержанием белков могут применяться как для предупреждения, так и для лечения рака.
Несмотря на то, что потребление белков связано с повышенной смертностью у взрослых среднего возраста в начале исследования, имелись доказательства того, рацион с низким содержанием белков может быть вредным для лиц пожилого возраста. Как высокое, так и умеренное потребление белков у лиц преклонного возраста было связано со значительными улучшениями в сравнении с группой с низким потреблением, подтверждая, что потребление белков, на которые приходится по меньшей мере 10% потребляемых калорий, может быть необходимым после наступления возраста 65 лет или возможно 75 лет для снижения зависимой от возраста потери массы и возможно предупреждения избыточного снижения уровня IGF-1 и других важных факторов. В предыдущих исследованиях было отмечено, повышенное потребление белков и повышение в результате уровня IGF-1 может оказаться благоприятным у лиц пожилого возраста. В действительности, неожиданный переход от защитного к неблагоприятному влиянию рациона с низким содержанием белков совпадает со временем, когда, как известно, масса стабилизируется, а затем снижается. С учетом предыдущих продольных исследований, масса проявляет тенденцию к увеличению вплоть до возраста 50-60 лет, в этот момент она стабилизируется перед непрерывным снижением в среднем на 0,5% в год у лиц, старше 65 лет. Полагаем, что это может зависеть от потери массы и болезненности рассматриваемых субъектов, при этом болезненные субъекты, которые потеряли значительную часть своей массы в процентах и имеют низкий уровень BMI, более восприимчивы к недостаточности белков. Также возможно, что другие факторы, такие как воспаление или генетические факторы, могут усиливать чувствительность к ограничению белков у пожилых субъектов, что соответствует нашим исследованиям на мышах.
Хотя в других исследованиях отмечали связанное с возрастом уменьшение всасывания нутриентов у грызунов, которое связано с изменениями рН микросреды, нарушением адаптационной реакции в кишечнике при старении, а также с изменениями в морфологии кишечника, до сих пор не установлена четкая взаимосвязь между морфологическими изменениями и изменениями абсорбции при старении. У людей в ряде исследований было показано, что кинетика расщепления и всасывания пищевых белков in vivo у здоровых пожилых людей не нарушена, однако в этих исследованиях также сообщали о повышении экстракции АА органами, что может привести к снижению доступности для периферических тканей, и предполагается, что в случае низкого потребления белков или повышенной потребности в белках ограниченная системная доступность пищевых АА может способствовать уменьшению синтеза мышечного белка. Более того, у людей другие факторы, такие как дефектная зубочелюстная система, прием лекарственных препаратов и психосоциальные проблемы, также играют существенную роль в показателях недостаточности питания.
Ранее было показано, что уровень IGF-1 снижается в пожилом возрасте, повышая в некоторой степени риск болезненности и смертности. Таким образом, наши результаты исследований могут объяснить противоречия в отношении IGF-I и смертности, указывая на то, что минимальные уровни белков и возможно IGF-1 являются важными в пожилом возрасте или что низкие уровни циркулирующего в крови IGF-1 отражают состояние связанной с недостаточностью питания болезненности и/или смертности. В действительности известно, что воспаление и другие нарушения уменьшают уровни IGF-1, повышая возможность того, что группа с низким потреблением белков и низким уровнем IGF-1 может содержать значительное количество как индивидуумов с недостаточностью питания, так и болезненных индивидуумов, имеющих основные заболевания или у которых они развиваются.
Необходимо признать, что в нашем исследовании существуют некоторые ограничения. Прежде всего, использование одного 24-часового воспроизведения режима питания с последующей 18-летней оценкой смертности может привести к неправильной группировке по режиму питания, если 24-часовой период не представлял собой обычный день. Однако 93% из нашей выборки сообщали, что 24-часовой период представлял обычный день. В нашем анализе также включили данную переменную в качестве контроля. Кроме того, было показано, что 24-часовое воспроизведение режима питания является достоверным подходом для определения "обычного рациона" субъектов. Хотя необходимо признать, что недостаток данных продольного анализа о потреблении пищи является потенциальным ограничением нашего исследования, исследование однородности пищевого рациона в течение более шести лет среди пожилых людей выявило незначительное изменение пищевых привычек с течением времени. Другое исследование, в котором изучали пищевые привычки в течение более двадцати лет, показало, что несмотря на то, что по мере старения людей потребление энергии, приходящейся на белки, жиры и углеводы, снижалось, снижения были равными среди трех типов.
Другое ограничение нашего исследования состоит в том, распределение респондентов на группы потребления белков с последующей стратификацией выборки для анализа в результате давало относительно малые объемы выборки, в особенности для анализов, включающих показатель смертности от диабета среди лиц без диабета в начале исследования или участников в подвыборке IGF-1. Вследствие этого наши соотношения рисков и 95% доверительные интервалы могут быть значительно больше тех, которые бы наблюдали при большем объеме выборки. Тем не менее, можно ожидать, что выборка малого размера будет уменьшать статистическую мощность нашего исследования и сделает более трудным выявление взаимосвязи. Следовательно, наша способность выявлять значимость указывает на то, что взаимосвязи между потреблением белков и смертностью весьма устойчивы. Кроме того, нижние пределы 95% доверительных интервалов из наших анализов смертности были значительно выше 1,0, означая, что повышенный риск, вероятно, является большим. И наконец, учитывая данные ограничения, наше исследование было усилено путем использования в нем надежных данных по смертности от конкретных причин, а также включения в него крупной общенациональной репрезентативной выборки - элемента, зачастую отсутствующего в предыдущих публикациях.
В целом, проведенные исследования на людях и животных показывают, что рацион с низким содержанием белков в среднем возрасте можно применять для предупреждения рака и общей смертности посредством процесса, который может включать по меньшей мере частичное регулирование уровней циркулирующего в крови IGF-1 и возможно уровней инсулина. В соответствии с другими эпидемиологическими исследованиями и исследованиями на животных наши результаты указывают на то, что рацион, в котором нутриенты растительного происхождения составляют большую часть потребляемой пищи, вероятно, обеспечивает максимальную пользу для здоровья. Однако, предполагается, что в возрасте до 65 лет и возможно 75 лет, в зависимости от состояния здоровья, опубликованное Советом по продовольствию и питанию Института медицины значение от 0,7 до 0,8 грамма белков/кг массы тела/день, рассматриваемое в настоящее время как минимальное требование, может оказывать защитный эффект по сравнению со значением 1-1,3 грамма белков/кг массы тела/день, потребляемым взрослыми в возрасте 19-70 лет. Также предполагается, что в пожилом возрасте может быть важным избегать низкого потребления белков и постепенно переходить на потребление по возможности преимущественно растительных белков в количестве от умеренного до высокого для обеспечения поддержания нормальной массы тела и защиты от болезненности.
Процедуры экспериментов
Потребление нутриентов для данных, полученных для людей
Данные о потреблении нутриентов базируются на отчетах о потреблении пищи и питья в течение 24-часового периода. Данные собирали посредством автоматизированной, основанной на применении персонального компьютера кодирующей системы с информацией по более чем восьмидесяти нутриентов. Существует ряд преимуществ использования данного способа для сбора данных о режиме питания. Учитывая, что время, проходящее между потреблением и воспроизведением этого факта, является коротким, участники, как правило, в состоянии вспомнить больше информации. Также, в отличие от методик создания отчетов, 24-часовое воспроизведение режима питания основано на сборе данных после потребления, что снижает возможность оценки для изменения пищевого поведения. Кроме того, было показано, что 24-часовое воспроизведение питания позволяет более убедительно оценить суммарное потребление энергии и белков по сравнению с широко используемыми опросниками частоты потребления различных пищевых продуктов и, как также было показано, является более адекватной техникой измерения суммарного потребления энергии и нутриентов, чем как Опросник с блоковой структурой частоты потребления различных пищевых продуктов, так и Опросник истории питания Национального института онкологии. И наконец, было выявлено, что данный подход точно оценивает потребление энергии, белков, жиров и углеводов независимо от индекса массы тела.
Последующее эпидемиологическое наблюдение в отношении смертности
Данные по смертности были доступны из Национального реестра смертности. Информацию по 113 потенциальным основным причинам смерти (UCOD-113) использовали для определения смертности по всем причинам, смертности от сердечно-сосудистого заболевания (CVD), смертности от рака и смертности от диабета.
Статистический анализ данных, полученных для людей
Модели пропорциональных рисков Кокса использовали для оценки связи между потреблением калорий, приходящихся на белки, с последующей смертностью по всем причинам, от CVD, от рака и от диабета - при этом три последних анализировали с помощью структур конкурирующих рисков. Затем проверяли взаимосвязь между возрастом и потреблением белков в связи со смертностью. На основе данных результатов субъектов распределили на две возрастные группы (50-65 лет и 66+ лет), которые использовали в остальных анализах. Модели пропорциональных рисков со стратификацией по возрасту использовали для оценки взаимосвязи процента калорий, приходящихся на белки, со смертностью в пределах двух возрастных групп, и проверки того, влиял ли на данную зависимость процент калорий, приходящихся на жиры, процент калорий, приходящихся на углеводы или на белки животного происхождения. Модели рисков повторно оценивали для подвыборки IGF-1 для определения, изменяло ли включение IGF-1 взаимосвязь между потреблением белков и смертностью. И наконец, модели пропорциональных рисков использовали для оценки взаимосвязи потребления белков и уровнем IGF-1 и использовали для подсчета прогнозированных соотношений рисков для каждой группы потребления белков при различных уровнях IGF-1, чтобы определить наличие дифференцированного влияния потребления белков на смертность в зависимости от уровней IGF-1. Все анализы проводили с использованием размеров выборки, учитывающих план выборки, и проверяя в отношении расы/этнической принадлежности, образования, пола, статуса наличия заболеваний, курения, изменений в режиме питания и суммарного потребления калорий.
Материалы и способы для экспериментов с дрожжами и мышами
При использовании моделей пропорциональных рисков Кокса не выявили взаимосвязь между потреблением белков и либо смертностью по всем причинам, от CVD, либо смертностью от рака (таблица 14). Однако высокое и умеренное потребление белков было прямо взаимосвязано со смертностью, связанной с диабетом. Одно из объяснений состоит в том, что диабет может быть более распространен в этих группах, возможно из-за перехода на более высокое потребление белков, более низкое потребление жиров и углеводов после установления диагноза диабета.
В заключение, было выявлено, что высокое потребление белков в сравнении с низким связано с более чем десятикратным повышением риска смертности от диабета для субъектов в возрасте 66 лет и выше. Однако причиной этого может быть значительно большая встречаемость субъектов с диабетом в анамнезе в группе с высоким потреблением белков и малое число субъектов, умерших от диабета, в группе с низким потреблением белков, что подчеркивает необходимость в дополнительных исследованиях для определения роли потребления белков в заболеваемости сахарным диабетом и смертности (HR: 10,64; 95% CI: 1,85-61,31).
Дополнительные материалы и способы
IGF-I в данных, полученных для людей
Половину субъектов в NHANES III случайным образом отбирали для участия в утреннем обследовании с последующим рекомендованным девятичасовым воздержанием от пищи. Из данной подвыборки 2253 субъекта, включенных в наше исследование, следовали предписаниям, и у них количественно оценивали данные по уровню IGF-I в сыворотке крови натощак. Уровень IGF-I определяли с помощью продукции Diagnostic Systems Laboratories Inc. с использованием стандартного лабораторного протокола и представляли в нг/мл.
Потенциальные искажающие факторы в данных, полученных для людей
Возраст, расу/этническую принадлежность, образование, пол, статус наличия заболеваний, курение, изменения в режиме питания и суммарное потребление калорий включали в анализы в качестве потенциальных искажающих факторов. Возраст приводили в годах, и совокупность данных из NHANES кодировали для защиты конфиденциальной информации респондентов, при этом возрасту 90 лет присваивали наивысшее значение кода. Фиктивные переменные вводили для распределения субъектов по трем категориям в зависимости от расы/этнической принадлежности: белые, не являющиеся латиноамериканцами, чернокожие, не являющиеся латиноамериканцами, и латиноамериканцы, при этом образование указывали по годам школьного обучения. Фиктивные переменные вводили для самоотчетов о статусе курильщика - "никогда", "в прошлом" и "в настоящее время". Субъектов просили сообщить об их анамнезе в виде вопросов, сформулированных как "Информировал ли вас доктор когда-либо о том, что у вас...", и использованных для введения трех фиктивных переменных в отношении наличия рака, инфаркта миокарда и диабета в анамнезе. Недавние изменения в пищевом рационе оценивали с помощью ответов на три вопроса: 1) "Пытались ли вы за прошедшие 12 месяцев сбросить вес?"; 2) "Изменяли ли вы за прошедшие 12 месяцев потребляемую пищу по медицинской причине или по состоянию здоровья?"; и 3) (после 24-часового воспроизведения режима питания) "Проведите сравнение пищи, которую вы употребили вчера, с вашей обычной". Окружность талии, являющей предпочтительной для BMI в качестве индикатора ожирения, измеряли с точностью до 0,1 см, начиная с правой стороны тела, по гребню подвздошной кости.
Модели рака на мышах
Все эксперименты на животных проводили в соответствии с процедурами, утвержденными Институциональным комитетом по уходу и использованию лабораторных животных USC. Для создания мышиной модели подкожного рака самцам мышей C57BL/6 в возрасте 18 недель, а также мышей GHRKO в возрасте 10 месяцев, соответствующим им по возрасту контрольным однопометным мышам и однопометным мышам дикого типа вводили клетки меланомы В16, а самкам мышей BALB/c в возрасте 12 недель - клетки рака молочной железы 4Т1. Перед введением клетки в логарифмической фазе роста собирали и суспендировали в бессывороточной среде Игла, модифицированной Дульбекко, с высоким содержанием глюкозы (DMEM) при 2×105 клеток или 2×106 и 100 мкл (2×104 клеток на мышь C57BL/6 или BALB/c; 2×105 клеток на мышь GHRKO) вводили подкожно в задненижнюю область. Перед подкожным введением опухоли всех мышей брили. Частоту возникновения опухолей определяли путем пальпирования участка введения, и размер опухоли определяли с использованием цифрового штангенциркуля, начиная с 10-15 дня после имплантации. Эксперименты с C57BL/6 и BALB/c завершали в различные моменты времени, исходя из утвержденных USC IACUC гуманных критериев конечных точек в отношении размера опухолей и образования язв. Мыши GHRKO (генетическая основа C57BL/6) были любезно предоставлены J.J. Kopchick (Университет Огайо, Афины).
Ограничение белков у мышей
Стандартный корм AIN-93G использовали в качестве референтного рациона с высоким содержанием белков на основе казеина (18% ккал приходится на белки, и рацион с низким содержанием белков 1,0 использовали в качестве рациона с низким содержанием белков на основе казеина (4% ккал приходится на белки) (Harlan Laboratories, Висконсин). Рационы являлись изокалорийными, и изменения в ккал, приходящиеся на жиры и углеводы, происходили пропорционально изменениям в ккал, приходящихся на белки. Определения суточного потребления начинали за 1 неделю перед проведением эксперимента, чтобы установить количество исходного потребления. Всех животных кормили ежедневно на протяжении эксперимента и их обеспечивали кормом в норме, превышающей на 50% их исходное потребление, для того, чтобы обеспечить не ограниченное по калорийности питание ad lib. Перед имплантацией опухоли мышей BALB/c распределяли в одну из 2 групп с различным количеством ккал, приходящихся на белки, и предварительно кормили в течение 1 недели. Данных мышей продолжали кормить на протяжении курса эксперимента аналогично описанному выше. Для определения влияния низкого потребления белков на старых мышей мышей C57BL/6 в возрасте 24 месяцев помещали в группу либо с 18%, либо с 4% ккал, приходящихся на белки, и непрерывно кормили рационом, как описано выше. Ежедневно определяли массу тела и уровень потребления. Животные имели постоянный доступ к воде.
Определения уровней mIGF-I и mIGFBP-1 в сыворотке крови у мышей
Для еженедельного получения сыворотки крови мышей анестезировали 3% изофлураном в ингаляционной форме, осторожно согревали для расширения вен и отбирали кровь из хвостовой вены. Анализы уровней mIGF-I и mIGFBP-1 в сыворотке крови проводили как описано ранее (Hwang et al., 2008), используя собственный анализ ELISA с применением рекомбинантного мышиного белка IGF-I или IGFBP-1 и поликлональных антител от R&D systems (Миннеаполис, Миннесота).
Статистический анализ данных, полученных для мышей
Сравнение групп по IGF-I проводили с использованием t-теста Стьюдента, сравнение групп по IGFBP-1 проводили с использованием t-теста Стьюдента и ANOVA, а сравнения групп по увеличению объема опухоли проводили с использованием двухфакторного дисперсионного анализа (ANOVA) при помощи GraphPad Prism v.6. Все статистические анализы были двухсторонними, а Р-значения <0,05 считали значимыми.
Определение выживаемости дрожжей и частоты возникновения мутаций
Клетки широко используемого штамма дрожжей DBY746 (МАТα leu2-3,112 his3-Δl trpl-289, ura3-52 GAL+) делали прототрофными путем трансформации соответствующей плазмидой, вносили в 1 мл полной синтетической среды (SDC) и выращивали на протяжении ночи при 30 градусах Цельсия на круговой качалке при 200 об./мин.
Данную стартовую культуру далее разделяли (1:100) и вносили в свежие синтетические среды SDC, содержащие стандартные аминокислоты в концентрации 0,5Х, 1X или 2Х (Hu et al., 2013) при отношении объема флакона к объему среды 5:1 и помещали обратно в термостат в те же самые условия. Аликвоты каждой культуры отбирали каждые два дня и соответствующие разведения высевали на чашки с обогащенной YPD. Колониеобразующие единицы (C.F.U.) подсчитывали после двухдневного роста. Оценивали процент выживших, рассматривая КОЕ в день 3 как 100% выживших. Все эксперименты проводили в трех повторах, и показано стандартное отклонение. Для подсчета частоты возникновения мутаций отбирали 107 в каждую временную точку выживания, промывали водой и высевали на полную синтетическую среду (SDC) с дефицитом аргинина и дополненную 60 □liq-1 канаванина (Can). После двух - трех дней выращивания при 30 градусах Цельсия определяли устойчивые к Сап колонии и выражали в виде числа устойчивых к Сап клонов из 106 жизнеспособных КОЕ.
Условия роста для эксперимента с Ras2
Хронологический жизненный цикл дрожжей изучали в среде SDC с истекшим сроком годности путем определения колониеобразующих единиц (КОЕ) каждые 48 ч. Число КОЕ в день 1 рассматривали как показатель исходной выживаемости (100%) и использовали для определения зависимой от возраста смертности.
Определения частоты возникновения мутаций в эксперименте с Ras2 с использованием Can1
Частоту возникновения спонтанных мутаций оценивали путем определения частоты возникновения мутаций в гене CAN1 (YEL063). Вкратце, выросшие за ночь инокуляты разводили жидкой средой SDC и инкубировали при 30°С. Жизнеспособность клеток определяли каждые 2 дня, начиная с 1 дня, путем высевания соответствующих разведений на чашки со средой с дрожжевым экстрактом, пептоном и декстрозой (YPD) и подсчета КОЕ. Для идентификации устойчивых к канаванину мутантов (Canr) в жидкой культуре соответствующее число клеток (начальное количество 2×107 клеток) отбирали путем центрифугирования, однократно промывали стерильной водой и высевали на селективную среду (SDC-Arg, дополненную 60 мкг/мл 1-канаванина сульфата). Мутантные колонии подсчитывали через 3-4 дня. Частоту возникновения мутаций выражали в виде отношения Canr к общему числу жизнеспособных клеток.
Исследования с низким потреблением белков у людей
Люди-субъекты принимали участие в 3 циклах 5-дневного потребления низкокалорийного и высокопитательного рациона с низким содержанием белков, имитирующего воздержание от пищи (FMD, отмечено зеленым, смотри текст), с последующим потреблением нормального рациона в течение примерно 3 недель (отмечено коричневым) (а). Кровь отбирали перед началом 5-дневного потребления рациона и при его завершении (временные точки А и В), а также через 5-8 дней после завершения 3-го 5-дневного потребления FMD (временная точка С). 5-Дневное потребление рациона существенно снижало уровни глюкозы (В), IGF-1 (с) и IGFBP-1 (D) в крови. Глюкоза *, р<0,05, N=18; IGF-1, **, р<0,01, *р<0,05, N=16; IGFBP-1, **, р<0,01, N=17; при этом все статистические тесты были выполнены в виде парного t-теста, двустороннего по исходным значениям. Результаты данного исследования представлены на фигуре 20.
Материалы и способы для экспериментов изучения химической интоксикации
2.1. Мыши. Все протоколы экспериментов с животными для мышей были утверждены Институциональным комитетом по уходу и использованию лабораторных животных (IACUC) Университета Южной Калифорнии. Самок мышей CD-I, BalB/С или C57BL/6N (Charles River) в возрасте 12-15 недель содержали в свободной от патогенной флоры среде в ходе всех экспериментов.
2.2. Рационы с определенным составом макронутриентов. Стандартный корм AIN93G (Harlan) использовали в качестве референтного рациона и предоставляли всем мышам, если не указано иное. Основой для рационов, модифицированных по составу макронутриентов (жиров, белков и углеводов), являлся AIN93G (фиг. 21 и таблица 20). В рационах 20% Р-1 (соевое масло в качестве источника жиров) и 20% Р-2 (кокосовое масло в качестве источника жиров) количество калорий, приходящихся на источники белков, было уменьшено до 20% в сравнении с составом AIN93G; рацион 0% Р не содержал белков; все данные рационы являлись изокалорийными по отношению к стандартному корму AIN93G. В рационе с низким содержанием углеводов LCHP количество приходящихся на углеводы калорий было уменьшено до 20% по сравнению с составом AIN93G (13% против 63,9%), но он содержал больше белков (45,2%) и жиров (41,8%). Кетогенный рацион с высоким содержанием жиров 60% HF был создан для обеспечения 60% потребляемых калорий, приходящихся на источники жиров, при этом калории, поступающие из белков и углеводов, пропорционально уменьшили. Рацион 90% HF представлял собой кетогенный рацион, содержащий 90% жиров, обеспечивая при этом лишь минимальное количество углеводов (менее 1%) и половину количества белков (9%). Подробный состав рациона и содержание калорий изложены в таблице S2. Перед тем, как приступить к экспериментам, мышей кормили контрольным рационом AIN93G, и исходя из их исходной массы тела, распределяли по экспериментальным группам (N=5/группа). Мыши привыкали к тестируемым рационам за неделю до экспериментов (схема адаптации показана в таблице 22). Все рационы предоставляли ad lib, если не указано иное.
2.3. Ограничение калорийности (CR) и кратковременное голодание (STS). Для ограничения калорийности с использованием рациона AJN93G стандартный корм размалывали в порошок и перемешивали с гидрогелем (Clear Н2О) в необходимых количествах с получение калорийной плотности 60%, 50%, 40%, 20%, 10% от AIN93G (таблица 23). Модифицированные по содержанию макронутриентов рационы с ограничением калорийности готовили аналогичным образом (таблица 24). Чтобы избежать недостаточности питания, все рационы дополняли витаминами, минеральными веществами, волокнами и незаменимыми жирными кислотами, соответствующими таковым в AIN93G. Исходное потребление пищи (3,7 г или 14 ккал/день) определяли путем кормления AIN93G перед экспериментом (данные не показаны). При режиме краткосрочного голодания (STS) мыши не имели доступа к пище в течение до 60 часов.
Во время всех экспериментов с CR и STS мышей содержали по одной в клетке в стандартных клетках типа "коробка для обуви", в которых ежедневно осуществляли чистку, чтобы не допустить копрофагии или поедания остатков корма. На протяжении всего времени животные имели доступ к воде и им предоставляли гидрогель для обеспечения достаточного восполнения жидкости. Массу тела каждого отдельного животного измеряли стандартно во время режимов с CR или STS.
2.4. Отбор крови для определения уровней глюкозы и IGF-1. Мышей анестезировали 2% изофлураном в ингаляционной форме и кровь отбирали с помощью пункции левого желудочка сердца. Кровь отбирали в пробирки, покрытые К2-EDTA, для получения сыворотки крови (BD). Уровень глюкозы в крови определяли с помощью системы Precision Xtra bloodglucose monitoring system (Abbott Laboratories). IGF-1 определяли с использованием специального набора для проведения ELISA у мышей (R&D Systems).
2.5. Устойчивость к высокодозной химиотерапии. Самок мышей CD-1 в возрасте 12-15 недель весом 25-32 г подвергали голоданию в течение до 60 ч. (STS) или кормили рационами с 50% CR, модифицированными по содержанию макронутриентов, в течение 3 дней с последующим внутривенным введением 24 мг/кг доксорубицина (DXR, Bedford Laboratories). Во всех экспериментах мышам давали стандартный корм AIN93G после инъекции химиотерапевтического препарата и проводили ежедневный контроль. Животных, проявляющих признаки тяжелого стресса и/или ухудшения состояния здоровья, признавали агонизирующими и подвергали эвтаназии.
2.6. Модель подкожной опухоли. Клетки модели рака молочной железы 4Т1 мышей и клетки глиомы GL26 поддерживали в DMEM (Invitrogen), дополненной 10% фетальной бычьей сывороткой (LBS), при 37°С с 5% СО2. Клетки в логарифмической фазе роста промывали и суспендировали в PBS при 2×106 клеток/мл и подкожно вводили (п/к 2×105 клеток/мышь в 100 мкл PBS) мышам в задненижнюю область. Используя штангельциркуль, измеряли размер опухоли. Для имитации многоциклового лечения, применяемого у людей, мышей 3 раза подвергали обработке путем внутривенного (в/в в латеральную хвостовую вену) введения цисплатина (Teva Parenteral Medicines Inc.) в дозе 12, 8 и 8 мг/кг массы тела, соответственно, три раза в дни 15, 33 и 44 после инокуляции опухоли. Осуществляли ежедневный контроль мышей, и животных, проявляющих признаки тяжелого стресса, ухудшения состояния здоровья или избыточную массу опухоли (2000 мм3) признавали агонизирующими и подвергали эвтаназии.
2.7. Статистический анализ. Сравнения между группами по показателям измерений глюкозы и IGF-1 осуществляли при помощи ANOVA с последующим множественным сравнением Тьюки, используя GraphPad Prism v. 5. Все статистические анализы были двухсторонними, а Р-значения <0,05 считали значимыми.
3. Результаты
3.1. Влияния ограничения калорийности на уровни глюкозы и IGF-1
Кратковременное голодание (STS) снижает уровни глюкозы и IGF-1 в сыворотке крови, повышает защиту клеток от высокодозной химиотерапии и повышает чувствительность злокачественных клеток к химиотерапевтическим лекарственным средствам. Влияния STS на уровень глюкозы и IGF-1 в основном достигаются, как только животные теряют примерно 20% массы тела. Таким образом, 20% потерю массы использовали в качестве критерия для сравнения уровней глюкозы и IGF-1 при потреблении рационов с ограниченной калорийностью с таковыми, полученным при режиме 60 ч. STS.
Порог 20% потери массы достигался в течение 4 дней при 90% CR, 6 дней при 80% CR, 9 дней при 60% CR или 13 дней при 40% CR (фиг. 22А и фиг. 26А). Время для достижения 20% потери массы в значительной степени зависит от жесткости ограничения калорийности (линейное приближение с r2=0,9976; фиг. 22 В). Через 48 часов снижение уровней глюкозы в крови коррелировало с жесткостью ограничения калорийности (линейное приближение с r2=0,7931; дополнительная фиг. 26 В). Режим 60 ч. воздержания от пищи (STS) снижал уровни глюкозы в крови на 70% по сравнению с таковыми у мышей, которых кормили ad lib (фиг. 22С, Р<0,001). 4-Дневный режим с 90% CR снижал уровни глюкозы в крови на примерно 40%, что существенно меньше, чем при STS (Р<0,05). Кроме того, наблюдали тенденцию в отношении влияния CR на снижение уровня глюкозы в крови в зависимости от CR-кормления: уровни глюкозы при 13-дневном кормлении с 40% CR были существенно (Р<0,05) ниже, чем в группе с 4-дневным 90% CR. Однако, в группе без ограничения калорийности полученные в результате уровни глюкозы в крови были более низкими, чем в группе 60 ч. воздержания от пищи; и требовалось 9 или более дней с CR для получения эффектов снижения уровня глюкозы в таком диапазоне, как в группе воздержания от пищи (фиг. 22С). У мышей во всех экспериментальных группах с CR, независимо от жесткости ограничения, достигались подобные уровни IGF-1 в сыворотке, как только достигался предел 20% потери массы, (Р<0,001), и имелись более низкие уровни IGF-1, чем у мышей в контрольной группе с кормлением ad lib (фиг. 22D).
3.2. Влияния рационов с определенным составом макронутриентов на уровни глюкозы и IGF-1
Ряд рационов с определенным составом макронутриентов (фиг. 21 и таблица 20) создавали на основе корма для грызунов AIN93G для того, чтобы определить, может ли ограничение определенных составляющих рациона имитировать влияния STS или краткосрочного CR на уровни глюкозы в крови и/или IGF-1 в сыворотке крови. В рационах с низким содержанием белков 20% Р-1 (соевое масло в качестве источника жиров) и 20% Р-2 (кокосовое масло в качестве источника жиров) количество калорий, приходящихся на источники белков, было уменьшено до 20% по сравнению с исходным составом AIN93G, тогда как количество углеводов и жиров было увеличено для поддержания изокалорийности рационов по отношению к AIN93G. Рацион 0% Р не содержал белков; количество углеводов, а также жиров было пропорционально увеличено для поддержания изокалорийности рациона по отношению к стандартному корму. В рационе LCHP количество калорий, приходящихся на источники углеводов, было уменьшено до 20% по сравнению с исходным составом AIN93G (13% против 63,9%) при обеспечении большего количества белков и жиров. Кетогенный рацион с высоким содержанием жиров 60% HF был создан для обеспечения 60% потребляемых калорий, приходящихся на источники жиров, при этом калории, поступающие из белков и углеводов, пропорционально уменьшили. Рацион 90% HF представлял собой кетогенный рацион, содержащий 90% калорий, приходящихся на жиры, обеспечивая при этом лишь минимальное количество (менее 1%) углеводов, и включающий 9% калорий, приходящихся на белки. Из-за более высокого содержания жиров рационы LCHP, 60% HF и 90% HF характеризуются высокой калорийной плотностью по сравнению со стандартным кормом AIN93G. Подробное описание состава рациона и содержание калорий приведены в таблице 21.
Самок мышей CD-I кормили ad lib экспериментальными рационами в течение девяти последовательных дней для установления профилей массы тела (фиг. 23А, В) и для контроля потребления калорий (фиг. 23С, D). Значительного отказа от пищи не наблюдали, но отметили, что мыши, которых кормили рационом с отсутствием белков (0% Р), через 6 дней уменьшили потребление пищи (фиг. 23С). Уменьшенное потребление калорий вызывало потерю массы у животных в данной экспериментальной группе (фиг. 23А). Мыши в группах на кетогенном рационе с высоким содержание жиров (60% HF и 90% HF) потребляли больше калорий в течение 9 дней кормления, чем мыши, которых кормили стандартным кормом AIN93G (фиг. 23D), и мыши, которых кормили ad lib кетогенным рационом 90% HF, быстро прибавляли в весе через 4-5 дней (фиг. 23 В). Мыши CD-I в экспериментальных группах, которых кормили рационами 20% Р и LCHP, не продемонстрировали различий в потреблении калорий или массе тела по сравнению с мышами, которых кормили контрольным рационом AIN93G (фиг. 23А, С).
Уровни глюкозы в крови в день 2, день 5 и день 9 у мышей на модифицированных по содержанию макронутриентов рационах не отличились от таковых на рационе со стандартным кормом (фиг. 27 и данные не показаны). В противоположность этому, уровни IGF-1 в сыворотке крови существенно повышались (Р<0,05) у мышей на кетогенном рационе 60% HF в течение 9 дней, но не у мышей, которых кормили кетогенным рационом 90% HF (фиг. 23 Е). Примечательно, что не только состав макронутриентов (например, содержание белков), но также источник жирных кислот по-разному модулировали уровни циркулирующего в крови IGF-1: рацион с низким содержанием белков 20% Р-1 (содержащий соевое масло в качестве единственного источника жиров) не снижал уровни IGF-1, а рацион с низким содержанием белков 20% Р-2 (коксовое масло в качестве единственного источника жиров) существенно (Р<0,05) снижал уровни IGF-1, при этом в данных рационах не было отличий, кроме источника жиров. Наиболее заметное влияние на уровень IGF-1 в сыворотке крови наблюдалось у мышей, которых кормили белководефицитным рационом 0% Р в течение 9 дней. Уровень циркулирующего в крови IGF-1 снижался примерно на 30% от такового у мышей на стандартном корме (фиг. 23Е). Белководефицитный рацион 0% Р был единственным рационом, который снижал уровни IGF-1 в сыворотке крови по сравнению с 60-ч. краткосрочным голоданием.
3.3. Краткосрочное ограничение калорийности и воздержание от пищи улучшают стрессоустойчивость
У мышей пониженные уровни IGF-1 в сыворотке крови и глюкозы в крови стимулируют способность преодолевать токсичность, индуцируемую высокодозным применением химиотерапевтических средств. Поскольку краткосрочное ограничение калорийности, а не рационы с определенным составом макронутриентов (кроме полного исключения белков), снижало уровни IGF-1 и глюкозы, использовали комбинаторный подход для проверки того, могут ли рационы с нехваткой определенных макронутриентов, которыми кормят при 50% ограничении калорийности от уровня регулярного суточного потребления калорий, обуславливать усиление защиты от химической интоксикации. Рационы 20% Р не включали в эксперименты по стрессоустойчивости из-за того факта, что рацион 0% Р показал гораздо более выраженные влияния на уровни IGF-1 в сыворотке крови.
Стрессоустойчивость проверяли у мышей CD-I, которых кормили либо ad lib стандартным кормом AIN93G, либо рационами с определенным составом макронутриентов с 50% ограничением калорийности от уровня нормального потребления калорий в течение трех дней перед обработкой доксорубицином (DXR, 24 мг/кг, в/в) (фиг. 24А). В группах с ограниченной на 50% калорийностью через 3 дня мыши теряли 12-15% от их исходной массы тела, тогда как в группе с STS мыши теряли 20% массы через 60 ч. После обработки с помощью DXR корм AIN93G предоставляли ad lib всем животным, и мыши повторно набирали массу до начала потери массы, индуцированной химической интоксикацией (фиг. 28А, В). Потеря массы продолжалась во всех экспериментальных группах до 8 дня после инъекции, после чего многие животные медленно восстанавливались. Мыши, которых кормили рационами 0% Р и LCHP с ограниченной калорийностью никогда полностью не восстанавливали свою исходную массу (фиг. 28А). Животные начали умирать в результате химической интоксикации через 9-18 дней после инъекции (фиг. 24А) в соответствии с опубликованным данными по дням начала и максимальной миелосупрессии после обработки с помощью DXR (http://dailymed.nlm.nih.gov). Мышей считали выжившими, если они были живы через 25 дней после введения DXR. Мыши, которых ad lib кормили рационом AIN93G за 3 дня до введения DXR, показали наихудший результат лишь с 16% выживаемостью к 25 дню (фиг. 24А). В отличие от мышей, которых кормили ad lib, подавляющее большинство (89%) мышей в режиме воздержания от пищи (60 часов) выжило после высокодозной химиотерапии. У контрольных мышей, обработанных DXR, наблюдались признаки интоксикации, в том числе сниженная подвижность, взлохмаченный шерстный покров и сгорбленная спина, тогда как мыши в группе STS не демонстрировали видимых признаков стресса или боли после обработки (данные не показаны). Трехдневное кормление комбинацией 50% CR с модифицированным содержанием макронутриентов до введения DXR улучшало у мышей стрессоустойчивость, результатом чего была 45-55% выживаемость (фиг. 24А). Не было никаких признаков того, что содержание жиров или углеводов оказывало влияние на результаты, поскольку со всеми рационами достигалась подобная степень защиты. Данные указывают на то, что краткосрочное CR, а не содержание жиров или углеводов в рационе, придает частичную защиту от химического воздействия, которая не настолько эффективна, как обеспечиваемая воздержанием от пищи. Мыши, которых кормили рационом LCHP с 50% CR демонстрировали худшие показатели, чем все остальные группы, которых кормили с CR, предположительно из-за влияния высокого содержания белков в данном рационе на уровень IGF-1.
Показатели уровней глюкозы в крови выявили, что трехдневное кормление модифицированными рационами с ограниченной калорийностью было недостаточным для существенного снижения уровней глюкозы, за исключением кетогенного рациона 90% HF с 50% CR (фиг. 24В). Снижение уровней глюкозы в группе кетогенного рациона, как оказалось, не усиливало стрессоустойчивость. У мышей в группе с STS уровни глюкозы в крови были существенно пониженными, в отличие от всех остальных экспериментальных групп (фиг. 24В).
3.4. Рацион с низким содержанием белков, очевидно, не замедляет прогрессирование развития глиомы GL26
Было показано, что рационы с низким содержанием белков снижают риск возникновения рака, тогда как высококалорийные рационы и рационы с высоким содержанием белков связанны с ожирением и активируют гормональные, метаболические и воспалительные изменения, которые модулируют канцерогенез. Для исследования воздействий рациона с низким содержанием белков в модели глиомы мышей переводили со стандартного корма (18,8% калорий приходится на белки, таблица 1) на рацион с низким содержанием белков (20% Р-1,3,9% калорий приходится на белки) через 10 дней после имплантации клеток GL26, когда опухоль можно было пальпировать (фиг. 25A). У мышей, которых кормили рационом с низким содержанием белков, наблюдалось прогрессирование развития опухоли, что не отличалось от такового у мышей, которых кормили ad lib рационом AIN93G (фиг. 25А). Данные результаты указывают на то, что прогрессирование развития опухоли невозможно замедлить путем ограничения белков, если опухоль уже образовалась.
3.5. Краткосрочное периодическое ограничение калорийности не повышает эффективность химиотерапии против рака молочной железы
Эффективность STS в усилении лечения различных форм рака состоит из двух эффектов: она защищает от индуцированной химиотерапией интоксикации нормальные клетки/ткани и повышает чувствительность злокачественных клеток к химиотерапевтическим средствам. Тем не менее, воздержание от пищи даже в течение короткого промежутка времени (например, 4 дней) может быть сложной задачей для большинства людей и, таким образом, "более мягкий" подход с ограничением калорийности может быть более осуществимым решением проблемы. Для проверки того, может ли краткосрочное периодическое потребление режима с 50% CR (ICR) привести в результате к благоприятным эффектам, подобно утвержденные протоколам воздержания от пищи, клетки рака молочной железы 4Т1 мышей подкожно имплантировали самкам мышей BalB/С и контролировали прогрессирование развития опухоли. Через двенадцать дней после имплантации опухолей измеряли объем опухолей и мышей распределяли либо в необработанную контрольную группу (AIN93G), группу, обработанную цисплатином (CDDP), или группу, которую периодически кормили с 50% CR (ICR) в течение трех дней перед обработкой цисплатином. Опухоль в необработанной контрольной группе быстро прогрессировала и к завершению эксперимента достигала объема 2000 мм3 через 54 дня после имплантации опухоли (фиг. 25В, черные круги). Три цикла обработки цисплатином замедляли прогрессирование развития опухоли; при этом объем опухолей у данных мышей составлял примерно половину размера таковых у необработанных мышей (фиг. 25В, голубые квадраты). В отличие от STS периодический режим кормления мышей в течение трех дней рационом AIN93G с ограниченной на 50% калорийностью перед введением цисплатина в результате не привел к повышению чувствительности опухоли и не усиливал действие химиотерапии (фиг. 25В, оранжевые треугольники). Объемы опухолей в данной экспериментальной группе существенно не отличались от объемов опухолей у мышей, которых только обрабатывали цисплатином.
4. Обсуждение
Ранее было показано, что значительное снижение уровней глюкозы и IGF-1 в крови в некоторой степени обуславливает благоприятные эффекты 2-3-дневного воздержания от пищи в животных моделях рака. У мышей 60-ч. краткосрочное воздержание от пищи уменьшает массу тела на 20% или более, IGF-1 в сыворотке крови на величину до 75% и уровень глюкозы на величину до 70%. В данных условиях животные становятся высокоустойчивыми к стрессу, что соответствует результатам, полученным с дрожжами, и у множества опухолей повышается чувствительность к химиотерапии и лучевой терапии. Если 20% потерю массы использовали в качестве ожидаемого результата, режимы с различной степенью CR приводили в результате к постоянно ускоряющейся потере массы, а также к снижению уровня IGF-1 и глюкозы. Однако, отмечали, что намного более короткий режим STS оказывал более выраженные влияния на уровень глюкозы, чем большинство рационов с CR, даже если рационы с CR потребляли в течение 9-13 дней, что приводило к потере массы, эквивалентной 20%. Менее выраженные влияния рационов с ограниченной калорийностью по сравнению с краткосрочным голоданием можно объяснить различной физиологической реакцией, которая является особой в условиях, при которых нутриенты полностью отсутствуют (Lee and Longo, 2011). Например, снижение уровня глюкозы в крови, вызванное краткосрочным воздержанием от пищи, в данном исследовании составляло 70% и возникало в течение 60 ч. по сравнению с 40% снижением уровня глюкозы, вызванным рационом с 90% CR через 96 ч.
При лишении пищи млекопитающие обычно проходят через три метаболические стадии: 1) постабсорбционная фаза, длящаяся в течение 10 или более часов после приема пищи, которая включает использование гликогена в качестве основного запаса источников энергии, 2) зависимое от аминокислот образование глюкозы путем глюконеогенеза, когда запас гликогена в печени истощается, и 3) фаза, в которой остаточная глюкоза преимущественно потребляется головным мозгом, тогда как глицерин и жирные кислоты высвобождаются из жировой ткани и становятся основным источником энергии. Образующиеся из жиров кетоновые тела становятся основными источниками углерода в течение нескольких дней воздержания от пищи. В организме данные изменения запускают клеточный ответ, включающий угнетающую регуляцию путей, вовлеченных в пролиферацию, рост клеток и пониженное образование активных форм кислорода с одновременным повышением стабильности генома и стрессоустойчивости клеток. Глюкоза представляет собой основной источник энергии для пролиферирующих клеток, таких как злокачественные клетки, и повышенные уровни глюкозы в крови были связаны с повышенным риском возникновения рака. Многие раковые клетки характеризуются повышенной интенсивностью поглощения глюкозы и зависят от гликолиза с последующим ферментативным расщеплением молочной кислоты даже в присутствии кислорода вместо гликолиза с последующим окислением пирувата, данный феномен известен как эффект Варбурга (Oudard et al., 1997; Warburg, 1956). В нормальных клетках снижение уровня глюкозы крови, а также IGF-1, по всей вероятности, способствует дифференциальной регуляции активации транскрипционных факторов стрессоустойчивости, которые регулируются чувствительными к поступлению нутриентов сигнальными путями и ходом клеточного цикла по принципу отрицательной обратной связи. В раковых клетках, наоборот, низкий уровень глюкозы представляет собой конкретную и основную проблему; в особенности, если также присутствуют химиотерапевтические препараты.
В соответствии с частичными эффектами в отношении уровня глюкозы в крови и IGF-1 результаты данного раскрытия указывают на то, что 50% CR в течение 72 часов, а также рационы, с ограниченным содержанием углеводов или белков, оказывают только частичные эффекты в отношении стрессоустойчивости. Комбинация краткосрочного периодического режима с 50% CR и обработки цисплатином, как было обнаружено, не вызывало усиления эффективности химиотерапии в отличие от комбинации STS и химиотерапии. Настоящее раскрытие свидетельствует о том, что три дня с 50% ICR не снижали существенно уровни глюкозы в крови и, таким образом, не могут обеспечить существенного уменьшения количества источников углерода, метаболизируемых клетками рака молочной железы мышей в течение данного промежутка времени. Ни один из этих рационов с 50% ограничением пищи и рационов с определенным составом макронутриентов, которыми кормили в течение 3 дней, за исключением кетогенного рациона 90% HF, не понижал уровни глюкозы в крови, что, как было показано, стимулирует защиту организма-хозяина и повышает чувствительность опухоли. Примечательно, что 50% снижение калорий, потребляемых с кетогенным рационом, приводит к 30% снижению уровней глюкозы в крови после трех дней кормления, эффект, который предположительно имеет место из-за очень низкого содержания углеводов (менее 1%) в данном рационе. Однако, эксперименты в отношении стрессоустойчивости в данном раскрытии указывают на то, что данное снижение не улучшает выживаемость. Кроме того, ни одна из мышей, получавших рационы с CR, не получала защиту, эквивалентную той, которая была вызвана 60 ч. воздержания от пищи (STS), в представленных в данном документе экспериментах. Дополнительные исследования с более длительными режимами кормления и экспериментальными группами большего объема будут необходимы для понимания того, могут ли конкретные рационы быть достаточными для получения эффектов DSR и DSS, которые близки к тем, которые вызваны циклами воздержания от пищи. В будущих исследованиях также можно будет оценить влияния различных рационов с определенными макронутриентами и с CR на продуцирование ROS, прогрессирование развития опухоли и стрессоустойчивость.
Белки рациона и суммарное содержание аминокислот, по всей видимости, влияют на продолжительность жизни и естественное старение. Ограничение потребления белков принимает участие в некоторых физиологических эффектах CR, включая снижение интенсивности метаболизма, снижение оксидативного повреждения, повышенную устойчивость печени к токсинам и онкогенным повреждающим факторам, уменьшение образования предраковых очагов и опухолей. Более того, ограничение как CR, так и белков снижает уровни IGF-1 в сыворотке крови, что может быть одним из факторов, способствующих увеличению продолжительности жизни, поскольку IGF-1-подобные сигнальные пути регулируют срок жизни у различных модельных организмов, таких как С. elegans, D. melanogaster и мыши. Было показано, что у животных путь IGF-1 воздействует как на срок жизни, так и на чувствительность к оксидативному стрессу, что соответствует большей устойчивости к оксидативному стрессу у мышей, дефектных по рецептору IGF-1. Белок forkhead box 01 (FOXO1), нисходящая мишень сигнального пути IGF-1/AKT, может проникать в ядро при отсутствии/снижении передачи сигнала сигнального пути IGF-1/AKT, где он может модулировать экспрессию широкого спектра генов, вовлеченных в устойчивость к оксидативному стрессу, продолжительность жизни и метаболизм, и, таким образом, он представляет собой ключевой механизм, вовлеченный в защиту от связанного с возрастом стресса и развития заболевания. Ранее было выдвинуто предположение, что результатом снижения уровня IGF-1 является улучшенная стрессоустойчивость в отношении высокодозной химиотерапии, а также повышение чувствительности опухоли. IGF-I оказывает сильный онкогенный эффект в отношении множества раковых клеток путем увеличения их скорости пролиферации и ингибирования апоптоза. Исследования у мышей с дефектами нижележащих эффекторов сигнального пути IGF-R, в том числе ингибирование mTOR посредством рапамицина и S6K1, демонстрирует центральную роль внутриклеточных митогенных путей ниже IGF-I в регулировании срока жизни и стрессоустойчивости с одновременным уменьшением роста опухолей. Кроме того, у людей с дефектом рецептора гормона роста имеются значительно более низкие уровни циркулирующего в крови IGF-1, а также у них наблюдается резко пониженная частота возникновения рака и диабета, которые более распространены среди родственников такого же возраста с интактным рецептором гормона роста.
Мыши из группы, которую кормили рационом с ограниченной калорийностью с низким содержанием углеводов (LCHP), характеризовались наихудшим показателем выживаемости среди всех групп с CR по сравнению с таким у мышей в контрольной группе. Тот факт, что мыши в данной группе потребляли аналогичные или более высокие количества калорий, получаемых из жиров, (20,9% в 50% CR LCHP в сравнении с 17,2% ad lib AIN93G), и что более важно, калорий, получаемых из белков, (22,6% в 50% CR LCHP в сравнении с 18,8% ad lib AIN93G) в течение трех дней кормления, может объяснить отсутствие защиты. Следует отметить, что результаты, представленные в эксперименте индуцирования стрессоустойчивости, базируются на относительно коротких (72 ч.) периодах кормления, таким образом, нельзя исключить, что более продолжительный режим CR либо с измененным количеством калорий и/или с ограничениями макронутриентов может в результате привести улучшенной стрессоустойчивости.
Кетогенные рационы широко используют для лечения рефрактерной эпилепсии у детей, но их также исследовали в отношении лечения рака. Для определения того, каким образом данный подход будет сопоставляться с нашими экспериментами в отношении стрессоустойчивости и потенциального повышения чувствительности опухолей, разработали два кетогенных рациона: собственный рацион 90% HF (% соотношение калорий по жирам:углеводам:белкам составляет 90%:1%:9%; фиг. 21) практически идентичен (разница ± 0,5%) классическому кетогенному рациону с соотношением жиры:углеводы:белки составляет 90%:1,4%:8,6%, соответственно (фиг. 29). Рацион с высоким содержанием жиров 60% HF (% соотношение калорий по жирам:углеводам:белкам составляет 60%: 31%: 9%) содержит жиры в соотношениях, подобных соотношению жиров в модифицированном рационе Аткинса (% соотношение калорий по жирам:углеводам:белкам составляет 60%:5%:35%; фиг. 29), но содержание белков было уменьшено, поскольку в предыдущих работах было установлено, что белки, а не углеводы, регулируют у человека уровни IGF-1. Описанные в данном документе результаты демонстрируют, что ни уровни глюкозы, ни IGF-1 существенно не снижались после кормления как первым, так и вторым кетогенным рационом в течение 9 последовательных дней.
Для оценки эффектов насыщенных жирных кислот в сравнении с ненасыщенными жирными кислотами, а также среднецепочечных жирных кислот в сравнении с длинноцепочечными жирными кислотами при лечении рака, разрабатывали два рациона, которые были изокалорийными относительно контрольного рациона с соевым маслом или кокосовым маслом в качестве источника жиров, но имели низкое содержание белков. Длинноцепочечные ненасыщенные жирные кислоты обнаружены в большинстве обычно используемых пищевых жиров и растительных масел, таких как соевое масло, тогда как коротко- и среднецепочечные насыщенные жирные кислоты (например, лауриновая и миристиновая кислота) обнаружены в относительном избытке в пальмоядровом масле и кокосовом масле. Среднецепочечные триглицериды (МСТ) могут легко гидролизоваться в желудочно-кишечном тракте и могут транспортироваться посредством системы воротной вены к гепатоцитам, тогда как большинство длинноцепочечных жирных кислот транспортируются в виде хиломикронов в лимфатической системе и упаковываются в триглицериды в печени. МСТ могут легко поступать в цикл Р-окисления в митохондриях, тогда как LCT зависят от переносчиков, таких как карнитин, чтобы проникать в матрикс митохондрий гепатоцитов. Данные исследований у людей также указывают, что потребление МСТ или рационов с повышенным отношением ненасыщенных жирных кислот к насыщенным жирным кислотам связано со снижением уровня глюкозы в крови, улучшением липидограммы и уменьшением ожирения. В исследовании биохимических и антропометрических профилей у женщин с абдоминальным ожирением добавка в рацион кокосового масла способствует уменьшению абдоминального ожирения.
Благоприятные эффекты продолжительного CR известны уже более века. Проблемы, связанные с реализацией CR в любом клиническом применении, состоят в том, что долговременное CR задерживает, но не останавливает прогрессирование многих злокачественных заболеваний и связано с длительным состоянием потери массы, что может быть неблагоприятным для раковых пациентов с кахексией или пациентов с риском возникновения кахексии, но также может вызывать долговременное уменьшение количества жиров и других запасов, что может повышать болезненность, особенно у пожилых пациентов. В действительности, продолжительное CR может замедлять заживление ран и иммунные функции, что может представлять собой дополнительную проблему для большинства пациентов, получающих химиотерапию или подвергающихся хирургическому лечению. Более того, 75% снижения уровня IGF-1 в сыворотке крови, вызванного 2-5-дневным воздержанием от пищи, у мышей и людей невозможно достигнуть при более умеренном CR, которое не снижает уровни IGF-1 у людей, если только потребление белков также не ограничено. Даже при сочетании с ограничением белков длительное CR вызывает лишь 30% снижение уровня IGF-1 у людей. Из-за постоянных эффектов в отношении уровней глюкозы и IGF-1 и последующих эффектов в отношении защиты нормальных клеток и повышения чувствительности раковых клеток без длительного дефицита массы тела, периодические циклы воздержания от пищи, очевидно, обладают высоким потенциалом для защиты пациентов, подвергаемых лечению различными химиотерапевтическими препаратами, при усилении их эффективности при лечении многих опухолей.
Хотя иллюстративные варианты осуществления описаны выше, это не означает, что данные варианты осуществления описывают все возможные формы настоящего изобретения. Скорее, слова, используемые в описании, являются описательными, а не ограничивающими словами, и следует понимать, что различные изменения могут быть произведены без отступления от сути и объема настоящего изобретения. Кроме того, признаки различных реализуемых вариантов осуществления могут быть объединены с получением дополнительных вариантов осуществления настоящего изобретения.
Реферат
Изобретение относится к медицине, а именно к эндокринологии, и может быть использовано для снижения уровней глюкозы у субъекта. Способ включает обеспечение субъекта рационом с низким содержанием белков, содержащим менее 10% калорий, приходящихся на источники белков, в течение первого периода времени, составляющего от 5 до 14 дней. Затем вводят субъекту второй рацион в течение второго периода времени после рациона с низким содержанием белков, где указанный второй рацион обеспечивает общее потребление калорий в пределах 20% от нормального потребления субъектом калорий, в течение 25-26 дней. Если определяют, что снижение уровней инсулиноподобного фактора роста 1 (IGF-I) является недостаточным, рацион с низким содержанием белков корректируют таким образом, чтобы обеспечить еще более низкие количества калорий, приходящихся на источники белков. Использование изобретения позволяет снижать уровень глюкозы и IGF-I за счет снижения содержания белка в рационе. 20 з.п. ф-лы, 34 ил., 7 табл.
Формула
Документы, цитированные в отчёте о поиске
Пищевые системы и способы увеличения долголетия
Комментарии