Белок, обладающий активностью в отношении стимуляции элонгации цепей жирных кислот, кодирующий его ген и их применение - RU2556375C1
Код документа: RU2556375C1
Чертежи


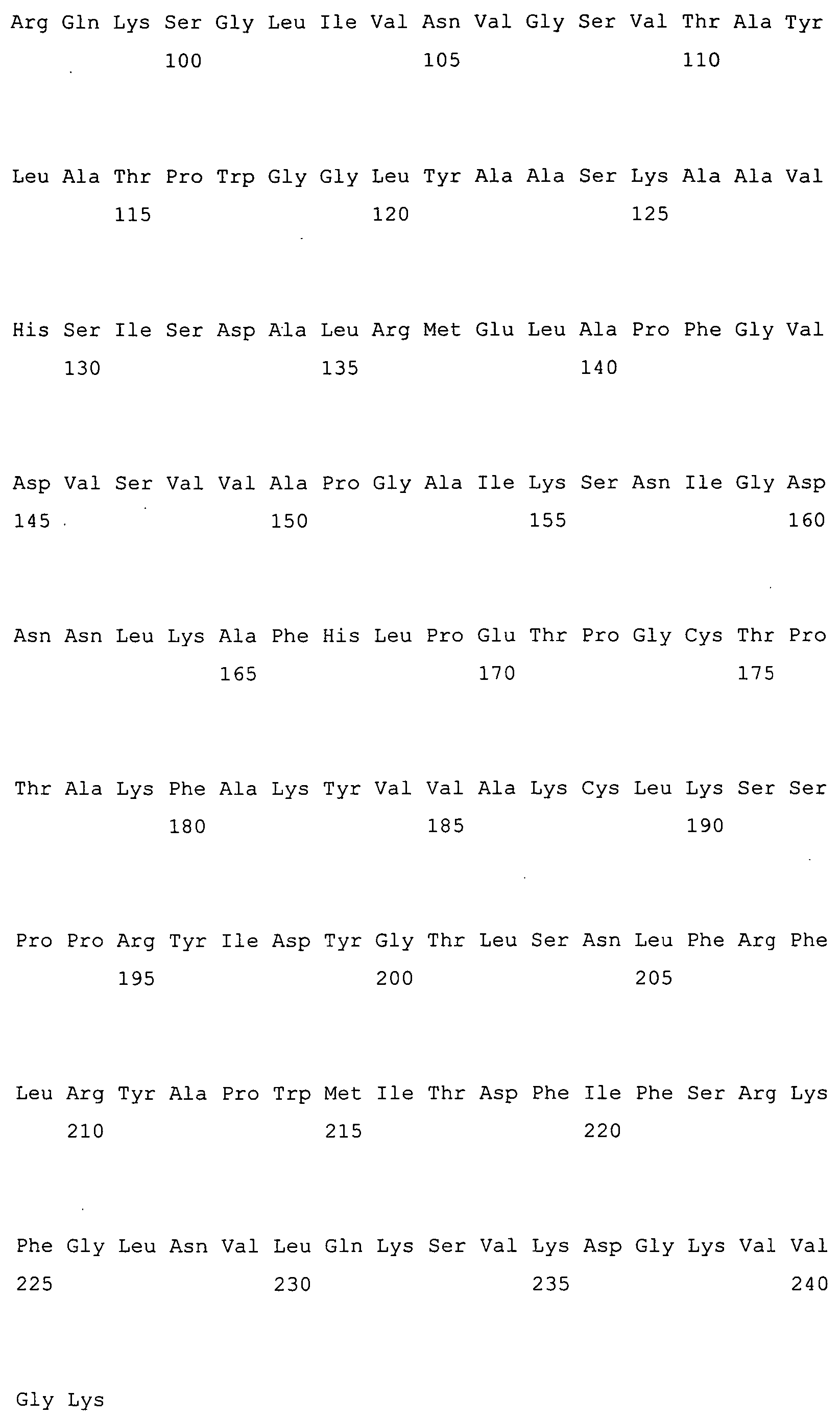





Описание
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0001] Настоящее изобретение относится к новому белку, обладающему активностью в отношении стимуляции элонгации цепей жирных кислот, кодирующему его полинуклеотиду и их применению.
УРОВЕНЬ ТЕХНИКИ
[0002] В дрожжах и других микроорганизмах реакция элонгации цепей жирных кислот осуществляется посредством четырех стадий, т.е. (i) реакции конденсации жирной кислоты ацил-СоА и малонил-СоА, (ii) реакции восстановления продукта конденсации 3-оксоацил-CoA, (iii) реакции дегидратации 3-гидроксиацил-CoA и (iv) реакции восстановления транс-2-еноил-CoA, посредством чего количество атомов углерода увеличивается на два за цикл этих стадий, удлиняя длину цепи (Непатентный документ 1).
Известно, что реакции в вышеуказанных стадиях (i)-(iv) катализируются ферментами (i) 3-кетоацил-CoA-синтазой, (ii) β-кетоацилредуктазой, (iii) 3-гидроксиацил-CoA-дегидрогеназой и (iv) еноил-CoA-редуктазой, соответственно (Непатентный документ 1).
Известно, что среди этих ферментов, 3-кетоацил-CoA-синтаза, которая является ответственной за реакцию конденсации, имеет специфичность в отношении жирных кислот, служащих в качестве субстратов, и ферменты, имеющие различные специфичности, клонировали из различных организмов.
В частности, в дрожжах, которые являются наиболее хорошо изученными грибами, для всех четырех стадий реакции элонгации цепей жирных кислот был идентифицирован фермент, ответственный за реакцию каждой стадии, и кодирующий его ген.
Например, в дрожжах имеются два гена, каждый из которых, как известно, кодирует фермент, ответственный за активность β-кетоацилредуктазы, и, кроме того, известно, что одновременная делеция обоих генов является летальной (Непатентный документ 2). Кроме того, известно также, что ген AYR1 имеет 1-ацил-дигидроксиацетонфосфат-редуктазную активность (Непатентный документ 3).
[0003] С другой стороны, PHS1 (эфирный) известен как ген, кодирующий фермент, ответственный за 3-гидроксиацил-CoA дегидрогеназную активность, тогда как TSC13 (эфирный) известен как ген, кодирующий фермент, ответственный за еноил-CoA-редуктазную активность.
В противоположность этому, известно, что липид-продуцирующий гриб, Mortierella alpina (M. alpina)), имеет гены 3-кетоацил-CoA-синтазы (так называемой элонгазы) (MALCE1 (ELO3), MALCE2, GLELO, MAELO), ответственные за первую реакцию, участвующую в элонгации цепей жирных кислот (Патентный документ 1), хотя гены для ферментов, других, чем 3-кетоацил-СоА-синтаза, еще не были идентифицированы.
Документы известного уровня техники
Патентные Документы
[0004] Патентный Документ 1: WO2010/147138
Непатентные Документы
[0005] Непатентный Документ 1: Kihara A., et al., (2008) J. Biol. Chem. 283, 11199-11209
Непатентный Документ 2: Han, G. et al., (2002) J. Biol. Chem. 277, 35440-35449
Непатентный Документ 3: Athenstaedt, K., and Daum, G. (2000) J. Biol. Chem. 275, 235-240
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0006] В этих условиях, имеется потребность в получении нового белка, участвующего в реакции элонгации цепей жирных кислот в клетках M. alpina, или гена, кодирующего этот белок.
[0007] В результате обширных и интенсивных усилий авторам изобретения удалось клонировать ген, кодирующий MaADR1, гомолог белка AYR1, который является 3-гидроксиацил-CoA-дегидрогеназой дрожжей, и тем самым было создано настоящее изобретение. Конкретно, настоящее изобретение относится к полинуклеотиду, белку, экспрессирующему вектору, трансформанту, способу получения композиции липида или жирной кислоты и пищевому продукту или тому подобное, полученным таким способом, как показано ниже.
[0008] Более подробно, настоящее изобретение включает следующие объекты.
[1] Полинуклеотид по любому из пунктов, выбранных из группы, состоящей из (a)-(e), показанных ниже:
(a) полинуклеотид, содержащий нуклеотидную последовательность SEQ ID NO: 1 или 4;
(b) полинуклеотид, кодирующий белок, который состоит из аминокислотной последовательности SEQ ID NO: 2;
(c) полинуклеотид, кодирующий белок, который состоит из аминокислотной последовательности с делецией, заменой, инсерцией и/или добавлением от 1 до 100 аминокислот в аминокислотной последовательности SEQ ID NO: 2, и который обладает активностью в отношении стимуляции элонгации цепей жирных кислот;
(d) полинуклеотид, кодирующий белок с аминокислотной последовательностью, которая на 60% или более идентична аминокислотной последовательности SEQ ID NO: 2, и который обладает активностью в отношении стимуляции элонгации цепей жирных кислот; и
(e) полинуклеотид, который гибридизируется в строгих условиях с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности SEQ ID NO: 1 или 4, и который кодирует белок, обладающий активностью в отношении стимуляции элонгации цепей жирных кислот.
[2] Полинуклеотид по [1] выше, который является любым из (f) или (g), показанных ниже:
(f) полинуклеотид, кодирующий белок, который состоит из аминокислотной последовательности с делецией, заменой, инсерцией и/или добавлением от 1 до 10 аминокислот в аминокислотной последовательности SEQ ID NO: 2, и который обладает активностью в отношении стимуляции элонгации цепей жирных кислот; и
(g) полинуклеотид, кодирующий белок с аминокислотной последовательностью, которая на 75% или более идентична аминокислотной последовательности SEQ ID NO: 2, и который обладает активностью в отношении стимуляции элонгации цепей жирных кислот.
[3] Полинуклеотид по [1] выше, который содержит нуклеотидную последовательность SEQ ID NO: 1 или 4.
[4] Полинуклеотид по [1] выше, который кодирует белок, состоящий из аминокислотной последовательности SEQ ID NO: 2.
[5] Полинуклеотид по любому из [1]-[4] выше, который является ДНК.
[6] Белок, кодируемый полинуклеотидом по любому из [1]-[5] выше.
[7] Вектор, содержащий полинуклеотид по любому из [1]-[5] выше.
[8] Трансформант, не являющийся человеческим, трансформированный полинуклеотидом по любому из [1]-[5] выше.
[9] Трансформант, не являющийся человеческим, трансформированный вектором по [7] выше.
[10] Трансформант по [8] или [9] выше, где этот трансформант является липид-продуцирующим грибом.
[11] Трансформант по [10] выше, где этим липид-продуцирующим грибом является Mortierella alpina.
[12] Способ получения композиции липида или жирной кислоты, который предусматривает сбор липидной композиции или композиции жирной кислоты из культивируемого продукта трансформанта по любому из [8]-[11] выше.
[13] Способ по [12] выше, где липидом является триацилглицерин.
[14] Способ по [12] выше, где жирная кислота имеет 18 или более атомов углерода.
[15] Пищевой продукт, фармацевтический препарат, косметический препарат или мыло, которые содержат липидную композицию или композицию жирной кислоты, собранные способом по [12] выше.
[0009] Полинуклеотид по изобретению может быть использован для трансформации липид-продуцирующих грибов (например, M. alpina), дрожжей, растений и так далее. Полученные таким образом трансформированные липид-продуцирующие грибы, трансформированные дрожжи или трансформированные растения и так далее могут быть использованы для получения композиций жирных кислот, пищевых продуктов, косметических препаратов, фармацевтических препаратов, мыла и так далее.
[0010] Более конкретно, трансформанты по изобретению имеют чрезвычайно высокую эффективность продукции липидов и жирных кислот. Таким образом, настоящее изобретение можно эффективно использовать для получения фармацевтических препаратов или диетических пищевых продуктов, в которых требуется большое количество липидов или жирных кислот.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0011] На Фигуре 1 показано сравнение геномных и CDS-последовательностей MaADR1.
На Фигуре 2 показана CDS-последовательность MaADR1 и ее расшифрованная аминокислотная последовательность.
На Фигуре 3 показана аминокислотная последовательность MaADR1 предположительного белка, полученного из Volvox carterif. nagariensis (зеленой водоросли) (GENEBANK accession No. XP_002946364) и AYR1p, полученного из S. cerevisiae (на этой фигуре дважды подчеркнутый сегмент представляет NADPH-связывающий сайт и звездочка (*) представляет активный центр).
НАИЛУЧШИЕ ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
[0012] Настоящее изобретение описано ниже более подробно. Нижеприведенные варианты осуществления приведены для иллюстрации изобретения и не предназначены для ограничения изобретения только этими вариантами осуществления. Изобретение может осуществляться в различных вариантах, не выходя за рамки существа изобретения.
Следует отметить, что все публикации, цитируемые в настоящем описании, в том числе документы уровня техники, патентные публикации и другие патентные документы, включены в настоящее описание в качестве ссылки. Кроме того, настоящее описание включает содержание, описанное в описании и фигурах Японской Патентной Заявки No. 2011-171044 (поданной 4 августа, 2011 года), в отношении которой испрашивается приоритет настоящей заявки.
[0013] Как описано более подробно ниже в разделе «Примеры», авторам изобретения удалось, раньше других, клонировать полноразмерную кДНК гена гомолога AYR1 (MaADR1) из липид-продуцирующего гриба, M. alpina. Кроме того, авторы изобретения идентифицировали нуклеотидную последовательность геномной ДНК, полученной из M. alpina MaADR1, и расшифровали аминокислотную последовательность. Эти ORF-последовательность, расшифрованная аминокислотная последовательность, CDS-последовательность и геномная последовательность MaADR1 показаны как SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3 и SEQ ID NO: 4, соответственно. Эти полинуклеотиды и ферменты могут быть получены, как описано далее в разделе «Примеры», известными способами генной инженерии, известными процедурами синтеза и так далее.
[0014] 1. Полинуклеотиды настоящего изобретения
Во-первых, настоящее изобретение относится к любому полинуклеотиду, выбранному из группы, состоящей из (a)-(e), показанных ниже.
(a) полинуклеотид, кодирующий нуклеотидную последовательность SEQ ID NO: 1 или 4;
(b) полинуклеотид, кодирующий белок, который состоит из аминокислотной последовательности SEQ ID NO: 2;
(c) полинуклеотид, кодирующий белок, который состоит из аминокислотной последовательности с делецией, заменой, инсерцией и/или добавлением от 1 до 100 аминокислот в аминокислотной последовательности SEQ ID NO: 2, и который обладает активностью в отношении стимуляции элонгации цепей жирных кислот;
(d) полинуклеотид, кодирующий белок с аминокислотной последовательностью, которая на 60% или более идентична аминокислотной последовательности SEQ ID NO: 2, и который обладает активностью в отношении стимуляции элонгации цепей жирных кислот: и
(e) полинуклеотид, который гибридизируется в строгих условиях с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной этой нуклеотидной последовательности SEQ ID NO: 1 или 4, и который кодирует белок, обладающий активностью в отношении стимуляции элонгации цепей жирных кислот.
[0015] В контексте настоящего изобретения, термин “полинуклеотид” предназначен для обозначения ДНК или РНК.
В контексте настоящего изобретения, выражение “полинуклеотид, который гибридизируется в строгих условиях” обозначает, например, полинуклеотид, который может быть получен способами гибридизации колоний, гибридизации бляшек, гибридизации по Саузерну или другими способами гибридизации с использованием, в качестве зонда, всего полинуклеотида или части полинуклеотида, состоящей из нуклеотидной последовательности, комплементарной нуклеотидной последовательности SEQ ID NO: 1 или 4, или полинуклеотид, состоящий из нуклеотидной последовательности, кодируемой аминокислотной последовательности SEQ ID NO: 2. Для гибридизации можно использовать способы, описанные, например, в "Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor, Laboratory Press 2001" и "Ausubel, Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997".
[0016] В контексте настоящего изобретения, термин “строгие условия” могут быть любыми условиями низкой строгости, условиями умеренной строгости и условиями высокой строгости. “Условия низкой строгости” относятся, например, к условиям 5 × SSC, 5 × раствор Денхардта, 0,5% ДСН, 50% формамид и 32°C. Подобным образом, “условия умеренной строгости” относятся, например, к условиям 5 × SSC, 5 × раствор Денхардта, 0,5% ДСН, 50% формамид и 42°C, или условиям 5 × SSC, 1% ДСЕ, 50 мМ Трис-HCl (pH 7,5), 50% формамид и 42°C. “Условия высокой строгости” относятся, например, к условиям 5 × SSC, 5 × раствор Денхардта, 0,5% SDS, 50% формамид и 50°C или условиям 0,2 × SSC, 0,1% ДСН и 65°C. При таких условиях можно ожидать, что ДНК, имеющая более высокую идентичность, эффективно получают при более высокой температуре. Однако на строгость гибридизации может влиять множество факторов, включающих температуру, концентрацию зонда, длину зонда, ионную силу, время реакции, концентрацию соли и так далее. Специалисты с квалификацией в данной области будут способны получить одну и ту же строгость выбором из факторов соответствующим образом.
[0017] Следует отметить, что при использовании для гибридизации коммерческого набора для этой цели может быть, например, использована система Alkphos Direct Labelling and Detection System (GE Healthcare). В этом случае, гибридизация может выполняться в соответствии с протоколом, приложенным к этому набору, т.е. мембрана может быть инкубирована в течение ночи с меченым зондом и затем промыта первым промывочным буфером, содержащим 0,1% (м/о) ДСН, при условиях 55°C для детектирования гибридизованной ДНК. Альтернативно, если коммерчески доступный реагент (например, смесь для ПЦР-мечения (Roche Diagnostics)) используют для мечения дигоксигенином (DIG) зонда во время приготовления зонда на основе всей нуклеотидной последовательности или части нуклеотидной последовательности, комплементарной нуклеотидной последовательности SEQ ID NO: 1 или 4, или нуклеотидной последовательности, кодирующей аминокислотную последовательность SEQ ID NO: 2, для детектирования гибридизации может быть использован набор детектирования нуклеиновой кислоты DIG (Roche Diagnostics).
[0018] Кроме перечисленных выше, другие гибридизируемые полинуклеотиды включают ДНК, которые на 50% или более, 51% или более, 52% или более, 53% или более, 54% или более, 55% или более, 56% или более, 57% или более, 58% или более, 59% или более, 60% или более, 61% или более, 62% или более, 63% или более, 64% или более, 65% или более, 66% или более, 67% или более, 68% или более, 69% или более, 70% или более, 71% или более, 72% или более, 73% или более, 74% или более, 75% или более, 76% или более, 77% или более, 78% или более, 79% или более, 80% или более, 81% или более, 82% или более, 83% или более, 84% или более, 85% или более, 86% или более, 87% или более, 88% или более, 89% или более, 90% или более, 91% или более, 92% или более, 93% или более, 94% или более, 95% или более, 96% или более, 97% или более, 98% или более, 99% или более, 99,1% или более, 99,2% или более, 99,3% или более, 99,4% или более, 99,5% или более, 99,6% или более, 99,7% или более, 99,8% или более или 99,9% или более, идентичны ДНК SEQ ID NO: 1 или 4 или ДНК, кодирующей аминокислотную последовательность SEQ ID NO: 2, как вычислено с помощью программы поиска гомологов, такой как FASTA или BLAST, с использованием параметров по умолчанию.
[0019] Следует отметить, что идентичность аминокислотных последовательностей или нуклеотидных последовательностей может быть определена FASTA (Science 227 (4693): 1435-1441, (1985)) или алгоритмом Karlin and Altschul, BLAST (Basic Local Alignment Search Tool) (Proc. Natl. Acad. Sci. USA 872264-2268, 1990; Proc Natl Acad Sci USA 90: 5873, 1993). На основе алгоритма BLAST, были разработаны программы, называемые blastn, blastx, blastp, tblastn и tblastx (Altschul SF, et al: J Mol Biol 215: 403, 1990). Если blastn используется для анализа нуклеотидной последовательности, могут быть установлены параметры, например, балл = 100 и длина слова = 12. Подобным образом, если используют blastp для анализа аминокислотной последовательности, параметры могут быть установлены, например, балл = 50 и длина слова = 3. Если используются программы BLAST и Gapped BLAST, в каждой программе могут быть использованы параметры по умолчанию.
[0020] Приведенные выше полинуклеотиды по изобретению могут быть получены с использованием известных способов генной инженерии или известных процедур синтеза.
[0021] 2. Белки изобретения
Настоящее изобретение относится к белкам, показанным в (i)-(iv) ниже.
(i) белок, который кодируется любым из полинуклеотидов, показанных в (a)-(e) выше;
(ii) белок, который содержит аминокислотную последовательность SEQ ID NO: 2;
(iii) белок, который содержит аминокислотную последовательность с делецией, заменой, инсерцией и/или добавлением одной или нескольких аминокислот SEQ ID NO: 2, и который имеет активность в отношении стимуляции элонгации цепей жирных кислот; и
(iv) белок с аминокислотной последовательностью, которая на 75% или более идентична аминокислотной последовательности SEQ ID NO: 2, и который обладает активностью в отношении стимуляции элонгации цепей жирных кислот.
[0022] Приведенный выше белок (iii) или (iv) является обычно мутантом природного белка SEQ ID NO: 2, хотя другие примеры включают белки, которые могут быть получены искусственно сайт-направленным мутагенезом, как описано в "Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor Laboratory Press 2001", "Ausubel, Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997", "Nuc. Acids. Res., 10, 6487 (1982)", "Proc. Natl. Acad. Sci. USA, 79, 6409 (1982)", "Gene, 34, 315 (1985)", "Nuc. Acids. Res., 13, 4431 (1985)", "Proc. Natl. Acad. Sci. USA, 82, 488 (1985)", и так далее.
[0023] В контексте настоящего изобретения, выражение “белок, который состоит из аминокислотной последовательности с делецией, заменой, инсерцией и/или добавлением одной или нескольких аминокислот в аминокислотной последовательности SEQ ID NO: 2, и который обладает активностью в отношении стимуляции элонгации цепей жирных кислот”, обозначает белки, которые состоят из аминокислотной последовательности с делецией, заменой, инсерцией и/или добавлением, например, от 1 до 100 аминокислотных остатков, от 1 до 90 аминокислотных остатков, от 1 до 80 аминокислотных остатков, от 1 до 70 аминокислотных остатков, от 1 до 60 аминокислотных остатков, от 1 до 50 аминокислотных остатков, от 1 до 40 аминокислотных остатков, от 1 до 39 аминокислотных остатков, от 1 до 38 аминокислотных остатков, от 1 до 37 аминокислотных остатков, от 1 до 36 аминокислотных остатков, от 1 до 35 аминокислотных остатков, от 1 до 34 аминокислотных остатков, от 1 до 33 аминокислотных остатков, от 1 до 32 аминокислотных остатков, от 1 до 31 аминокислотных остатков, от 1 до 30 аминокислотных остатков, от 1 до 29 аминокислотных остатков, от 1 до 28 аминокислотных остатков, от 1 до 27 аминокислотных остатков, от 1 до 26 аминокислотных остатков, от 1 до 25 аминокислотных остатков, от 1 до 24 аминокислотных остатков, от 1 до 23 аминокислотных остатков, от 1 до 22 аминокислотных остатков, от 1 до 21 аминокислотных остатков, от 1 до 20 аминокислотных остатков, от 1 до 19 аминокислотных остатков, от 1 до 18 аминокислотных остатков, от 1 до 17 аминокислотных остатков, от 1 до 16 аминокислотных остатков, от 1 до 15 аминокислотных остатков, от 1 до 14 аминокислотных остатков, от 1 до 13 аминокислотных остатков, от 1 до 12 аминокислотных остатков, от 1 до 11 аминокислотных остатков, от 1 до 10 аминокислотных остатков, от 1 до 9 аминокислотных остатков (одного или нескольких аминокислотных остатков), от 1 до 8 аминокислотных остатков, от 1 до 7 аминокислотных остатков, от 1 до 6 аминокислотных остатков, от 1 до 5 аминокислотных остатков, от 1 до 4 аминокислотных остатков, от 1 до 3 аминокислотных остатков, от 1 до 2 аминокислотных остатков или единственного аминокислотного остатка в аминокислотной последовательности SEQ ID NO: 2, и который обладает активностью в отношении стимуляции элонгации цепей жирных кислот. Обычно, меньшее количество является более предпочтительным для приведенных выше делеции, замены, инсерции и/или добавления аминокислотных остатков.
[0024] Кроме того, примеры таких белков включают белки, аминокислотная последовательность которых на 60% или более, 61% или более, 62% или более, 63% или более, 64% или более, 65% или более, 66% или более, 67% или более, 68% или более, 69% или более, 70% или более, 71% или более, 72% или более, 73% или более, 74% или более, 75% или более, 76% или более, 77% или более, 78% или более, 79% или более, 80% или более, 81% или более, 82% или более, 83% или более, 84% или более, 85% или более, 86% или более, 87% или более, 88% или более, 89% или более, 90% или более, 91% или более, 92% или более, 93% или более, 94% или более, 95% или более, 96% или более, 97% или более, 98% или более, 99% или более, 99,1% или более, 99,2% или более, 99,3% или более, 99,4% или более, 99,5% или более, 99,6% или более, 99,7% или более, 99,8% или более, или 99,9% или более идентична аминокислотной последовательности SEQ ID NO: 2, и которые обладают активностью в отношении стимуляции элонгации цепей жирных кислот. Обычно, более высокая величина является более предпочтительной для вышеуказанной идентичности.
[0025] Если определенный белок имеет “активность стимуляции элонгации цепей жирных кислот”, когда этот белок экспрессируется в подходящих клетках-хозяевах, (например, клетках дрожжей, липид-продуцирующих грибов и растений), эти клетки повышают содержание жирных кислот, имеющих больше атомов углерода, среди жирных кислот, которые могут быть синтезированы этими клетками, в сравнении с клетками того же типа, в которых не был экспрессирован этот белок. В этом случае, для синтеза большего числа “жирных кислот, имеющих больше атомов углерода”, используют жирные кислоты, имеющие на два меньше атомов углерода, и, следовательно, количество этих жирных кислот, имеющих на два меньше атомов углерода, может быть уменьшено.
Более конкретно, если белок с активностью в отношении стимуляции элонгации цепей жирных кислот, экспрессируется в вышеуказанных клетках-хозяевах, то эти клетки обнаруживают (i) уменьшение в количестве жирных кислот с 16 атомами углерода, и увеличение в количестве жирных кислот с 18 атомами углерода, (ii) уменьшение в количестве жирных кислот с 17 атомами углерода, и увеличение в количестве жирных кислот с 19 атомами углерода, (iii) уменьшение в количестве жирных кислот с 18 атомами углерода, и увеличение в количестве жирных кислот с 20 атомами углерода, (iv) уменьшение в количестве жирных кислот с 20 атомами углерода, и увеличение в количестве жирных кислот с 22 атомами углерода, (v) уменьшение в количестве жирных кислот с 22 атомами углерода, и увеличение в количестве жирных кислот с 24 атомами углерода, (vi) уменьшение в количестве жирных кислот с 24 атомами углерода, и увеличение в количестве жирных кислот с 26 атомами углерода, (vii) уменьшение в количестве жирных кислот с 26 атомами углерода, и увеличение в количестве жирных кислот с 28 атомами углерода, или (viii) уменьшение в количестве жирных кислот с 28 атомами углерода, и увеличение в количестве жирных кислот с 30 атомами углерода, в сравнении с клетками того же самого типа, где этот белок не экспрессировался.
Следует отметить, что активность в отношении стимуляции элонгации цепей жирных кислот может быть измерена, как описано в Han, G. et al., (2002) J. Biol. Chem. 277, 35440-35449.
Кроме того, для подтверждения активности стимуляции элонгации цепей жирных кислот, могут быть приведены примеры экспериментов, в которых используются подходящие клетки-хозяева, такие как клетки дрожжей, липид-продуцирующих грибов и растений. Когда полинуклеотид, кодирующий белок по изобретению, экспрессируется в клетках-хозяевах, белок или пептид, кодируемый этим полинуклеотидом, может быть определен как обладающий активностью в отношении стимуляции элонгации цепей жирных кислот, если увеличивается продуцирование жирных кислот с более длинной цепью. В разделе «Примеры», авторы настоящего изобретения подтвердили уменьшение в количестве жирных кислот с 16 атомами углерода, и увеличение в количестве жирных кислот с 18 атомами углерода, когда белок настоящего изобретения экспрессируется в клетках дрожжей и композиция жирных кислот, содержащихся в этих клетках дрожжей, анализируется газовой хроматографией.
Поскольку дрожжи были способны синтезировать жирные кислоты, имеющие до 18 атомов углерода, количество жирных кислот с 18 атомами углерода увеличивалось в разделе «Примеры». Если в качестве клеток-хозяев используют клетки, способные синтезировать жирные кислоты с более чем 18 атомами углерода, например, жирные кислоты с 19 или 20 атомами углерода (например, Mortierella alpina), произойдет увеличение в количестве жирных кислот, имеющих больше атомов углерода (например, жирных кислот с 20 атомами углерода, в случае Mortierella alpina), среди жирных кислот, которые могут быть синтезированы этими клетками-хозяевами.
[0026] Белок по настоящему изобретению предпочтительно проявляет активность в отношении стимуляции элонгации цепей жирных кислот на жирных кислотах, содержащихся в триацилглицеринах.
Кроме того, жирные кислоты, подлежащие элонгации белком настоящего изобретения, могут быть либо насыщенными жирными кислотами, либо ненасыщенными жирными кислотами, предпочтительно ненасыщенными жирными кислотами, более предпочтительно одновалентными, двухвалентными, трехвалентными или четырехвалентными ненасыщенными жирными кислотами.
Поскольку белок по настоящему изобретению является белком-гомологом белка AYR1 дрожжей, он мог бы иметь β-кетоацилредуктазную активность, как в случае AYR1. Известно, что в дрожжах не только AYR1, но также IFA38 известен как ген, имеющий β-кетоацилредуктазную активность, и было также известно, что одновременная делеция обоих генов является летальной (Han, G. et al., (2002) J. Biol. Chem. 277, 35440-35449). Имеет или не имеет белок настоящего изобретения β-кетоацилредуктазную активность, можно подтвердить определением, может ли дрожжевой штамм, гены AYR1 и IFA38 которого разрушены, расти, или его β-кетоацилредуктазная активность компенсируется, когда в нем экспрессируется белок настоящего изобретения.
[0027] Делеция, замена, инсерция и/или добавление одного или нескольких аминокислотных остатков в аминокислотной последовательности этого белка по настоящему изобретению означают, что делеция, замена, инсерция и/или добавление одного или нескольких аминокислотных остатков происходит в любом одном или нескольких положениях в одной и той же последовательности, и одновременно могут осуществляться два или более из этих событий, т.е. делеции, замены, инсерции и добавления.
[0028] Примеры взаимозаменяемых аминокислотных остатков показаны ниже. Аминокислотные остатки, включенные в одну и ту же группу, являются взаимозаменяемыми друг с другом. Группа А: лейцин, изолейцин, норлейцин, валин, норвалин, аланин, 2-аминобутановая кислота, метионин, о-метилсерин, т-бутилглицин, т-бутилаланин, циклогексилаланин; группа В: аспарагиновая кислота, глутаминовая кислота, изоаспарагиновая кислота, изоглутаминовая кислота, 2-аминоадипиновая кислота, 2-аминосубериновая кислота; группа С: аспарагин, глутамин; группа D: лизин, аргинин, орнитин, 2,4-диаминобутановая кислота, 2,3-диаминопропионовая кислота; группа E: пролин, 3-гидроксипролин, 4-гидроксипролин; группа F: серин, треонин, гомосерин; группа G: фенилаланин, тирозин.
[0029] Кроме того, белок по настоящему изобретению может быть получен способами химического синтеза, такими как Fmoc-способ (флуоренилметилоксикарбониловый способ) и tBoc-способ (т-бутилоксикарбониловый способ). Альтернативно, белок настоящего изобретения может также синтезироваться химически пептидными синтезаторами, коммерчески доступными из Advanced Automation Peptide Protein Technologies, Perkin Elmer, Protein Technologies, PerSeptive, Applied Biosystems, SHIMADZU, и так далее.
[0030] 3. Вектор по изобретению и трансформант, трансформированный этим вектором
В другом варианте осуществления настоящее изобретение относится также к экспрессирующему вектору, содержащему полинуклеотид по изобретению.
Вектор по настоящему изобретению обычно конструируют так, что он содержит:
(i) промотор, транскрибируемый в клетках-хозяевах;
(ii) полинуклеотид, показанный в любом из (a)-(g) выше, который лигирован с этим промотором; и
(iii) экспрессионную кассету, содержащую, в качестве элементов-компонентов, сигналы, которые функционируют в клетках-хозяевах для терминации транскрипции и полиаденилирования РНК-молекулы.
Такой сконструированный вектор вводят в клетки-хозяева. Примеры подходящих клеток-хозяев, используемых в настоящем изобретении, включают липид-продуцирующие грибы, дрожжи и тому подобное.
[0031] В качестве липид-продуцирующих грибов могут быть использованы штаммы, которые могут быть найдены в MYCOTAXON, Vol. XLIV, No. 2, pp. 257-265 (1992). Конкретные примеры включают микроорганизмы, принадлежащие к роду Mortierella, примерами которых являются микроорганизмы, принадлежащие к подроду Mortierella, такие как Mortierella elongate IFO8570, Mortierella exigua IFO8571, Mortierella hygrophila IFO5941, Mortierella alpina IFO8568, ATCC16266, ATCC32221, ATCC42430, CBS 219.35, CBS224.37, CBS250.53, CBS343.66, CBS527.72, CBS528.72, CBS529.72, CBS608.70, CBS754.68, и так далее., а также микроорганизмы, принадлежащие к подроду Micromucor, такие как Mortierella isabellina CBS194.28, IFO6336, IFO7824, IFO7873, IFO7874, IFO8286, IFO8308, IFO7884, Mortierella nana IFO8190, Mortierella ramanniana IFO5426, IFO8186, CBS112.08, CBS212.72, IFO7825, IFO8184, IFO8185, IFO8287, Mortierella vinacea CBS236.82, и так далее. Особенно предпочтительным является Mortierella alpina.
[0032] Подобным образом, примеры дрожжей включают Saccharomyces cerevisiae EH13-15, NBRC1951, NBRC1952, NBRC1953, NBRC1954, и так далее.
[0033] Клетки-хозяева, трансформированные вектором по настоящему изобретению, обнаруживают увеличение в количестве жирных кислот с длинной цепью (например, жирных кислот, имеющих 18, 19 или 20 атомов углерода, или жирных кислот, имеющих больше атомов углерода) в сравнении с клетками-хозяевами, которые не являются трансформированными вектором настоящего изобретения. Предпочтительно, вышеуказанные жирные кислоты являются кислотами, содержащими триацилглицерины (которые называют также “триглицеридами”).
Для введения в липид-продуцирующие грибы, вектором, доступным для применения, может быть, но не ограничивается им, pDura5 (Appl. Microbiol. Biotechnol., 65, 419-425, (2004)).
[0034] Для введения в дрожжи, может быть использован любой вектор, пока он имеет способность экспрессировать инсерт в дрожжевых клетках. Примеры включают pYE22m (Biosci. Biotech. Biochem., 59, 1221-1228, 1995).
[0035] Для регуляции экспрессии гена в клетках-хозяевах, может быть использована любая комбинация промотора и терминатора, пока они функционируют в этих клетках-хозяевах. Примеры включают промотор гена гистона Н4.1, промотор гена глицеральдегид-3-фосфат-дегидрогеназы и так далее, при использовании в липид-продуцирующих грибах.
Маркеры селекции, доступные для применения в трансформации, включают ауксотрофные маркеры (ura5, niaD, trp1), маркеры устойчивости к лекарственным средствам (гигромицин, зеоцин), ген устойчивости к генетицину (G418r), ген устойчивости к меди (CUP1) (Marin et al., Proc. Natl. Acad. Sci. USA, 81, 337 1984), ген устойчивости к церуленину (fas2m, PDR4) (Junji Inokoshi et al., Biochemistry, vol. 64, p. 660, 1992; Hussain et al., gene, 101, 149, 1991), и так далее.
[0036] Для трансформации клеток-хозяев могут быть использованы обычно используемые известные способы. Например, в случае липид-продуцирующих грибов, можно использовать электропорацию (Mackenxie D. A. et al. Appl. Environ. Microbiol., 66, 4655-4661, 2000) и способ доставки частиц (описанный в JP 2005-287403, озаглавленный “Способ разведения липид-продуцирующих грибов”). Подобным образом, в случае дрожжей, трансформация может выполняться, но не ограничивается ими, посредством электропорации, способа сферопластов (Proc. Natl. Acad. Sci. USA, 75 p1929 (1978)), литий-ацетатного способа (J. Bacteriology, 153, p163 (1983)) и других способов, описанных в Proc. Natl. Acad. Sci. USA, 75 p1929 (1978), Methods in yeast genetics, 2000 Edition: A Cold Spring Harbor Laboratory Course Manual.
[0037] Кроме того, в качестве стандартных способов клонирования, может быть сделана ссылка на "Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor Laboratory Press 2001," "Methods in Yeast Genetics, A laboratory manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY)", и так далее.
[0038] 4. Способ получения липидной композиции или композиции жирных кислот по настоящему изобретению
Еще в одном варианте осуществления настоящее изобретение относится также к способу получения липидной композиции или композиции жирных кислот с использованием вышеупомянутого трансформированного липид-продуцирующего гриба или дрожжей.
В контексте настоящего изобретения термин “липид” обозначает простой липид, в том числе соединение (например, глицерид), который состоит из жирной кислоты и спирта, присоединенного через сложноэфирную связь, или его аналога (например, сложного эфира холестерина); сложного липида, который генерируется из такого простого липида частичной модификацией с фосфорной кислотой, аминокислотой (аминокислотами), сахаридом (сахаридами) и тому подобное; или производного липида, который является гидролизатом вышеописанного липида и является нерастворимым в воде.
В контексте настоящего изобретения, термин “жир или масло” относится к сложному эфиру (глицериду), состоящему из глицерина и жирной кислоты.
В контексте настоящего изобретения, термин “жирная кислота” относится к алифатической монокарбоновой кислоте (т.е. карбоновой кислоте, имеющей единственную карбоксильную группу и атомы углерода, связанные в цепи), представленной общей формулой RCOOH (где R обозначает алкильную группу). Жирные кислоты включают насыщенные жирные кислоты, не имеющие двойной связи в углеводородной цепи, и ненасыщенные жирные кислоты, имеющие двойные связи в этой углеводородной цепи.
[0039] Липидная композиция или композиция жирных кислот по настоящему изобретению может быть экстрагирована следующим образом из клеток, которые были трансформированы в соответствии с данным изобретением:
После культивирования трансформированный штамм организма (например, липид-продуцирующий гриб или дрожжи) обрабатывают стандартным образом, например, центрифугированием или фильтрованием, для получения культивированных клеток. Эти клетки промывают хорошо водой и предпочтительно дополнительно сушат. Сушка может выполняться лиофилизацией, воздушной сушкой и так далее. Высушенные клетки необязательно гомогенизируют, например, при помощи Dynomil или обрабатывают ультразвуком и затем экстрагируют органическим растворителем предпочтительно под током азота. Органические растворители, используемые для этой цели, включают простой эфир, гексан, метанол, этанол, хлороформ, дихлорметан, петролейный эфир и так далее. Альтернативно, хорошие результаты могут быть также получены чередующейся экстракцией метанолом и петролейным эфиром или экстракцией однофазной системой растворителя хлороформ-метанол-вода. При отгонке органического растворителя из этого экстракта при пониженном давлении могут быть получены содержащие жирную кислоту липиды. Эти экстрагированные жирные кислоты могут быть превращены в соответствующие метиловые эфиры способом хлористоводородной кислоты-метанола и так далее.
[0040] Кроме того, жирные кислоты могут быть отделены в состоянии смешанных жирных кислот или смешанных эфиров жирных кислот от вышеупомянутых содержащих жирную кислоту липидов концентрированием и отделением стандартным образом (например, добавлением мочевины, отделением при охлаждении, колоночной хроматографией).
[0041] Липиды, полученные способом настоящего изобретения, являются предпочтительно триацилглицеринами, более предпочтительно триацилглицеринами, содержащими жирные кислоты с 18 или более атомами углерода.
Подобным образом, жирные кислоты, полученные этим способом настоящего изобретения являются предпочтительно жирными кислотами с 18 или более атомами углерода, более предпочтительно жирными кислотами с 18 или более атомами углерода, содержащимися в триацилглицеринах.
Примеры жирных кислот с 18 или более атомами углерода, включают, но не ограничиваются ими, стеариновую кислоту (18:0), олеиновую кислоту (18:1(9)), вакценовую кислоту (18:1(11)), линолевую кислоту (18:2(9,12)), α-линоленовую кислоту (18:3(9,12,15)), γ-линоленовую кислоту (18:3(6,9,12)), элеостеариновую кислоту (18:3(9,11,13)), арахидиловую кислоту (20:0), эйкозеновую кислоту (20:1Δ11), 8,11-эйкозадиеновую кислоту (20:2(8,11)), 5,8,11-эйкозатриеновую кислоту (20:3(5,8,11)), арахидоновую кислоту (20:4(5,8,11,14)), бегеновую кислоту (22:0), лигноцериновую кислоту (24:0), нервоновую кислоту (24:1), церотовую кислоту (26:0), монтановую кислоту (28:0) и мелиссиновую кислоту (30:0).
Кроме того, жирные кислоты, полученные способом по изобретению, могут быть либо насыщенными жирными кислотами, либо ненасыщенными жирными кислотами, предпочтительно ненасыщенными жирными кислотами, более предпочтительно одновалентными, двухвалентными, трехвалентными или четырехвалентными ненасыщенными жирными кислотами.
Следует отметить, что липиды, получаемые способом по изобретению, и жирные кислоты, содержащиеся в этих липидах, могут быть подтверждены в отношении их состава вышеуказанными процедурами для экстракции липидов или разделения жирных кислот или их комбинаций.
[0042] Липидные композиции или композиции жирных кислот, полученные способом по изобретению, могут использоваться, например, для получения содержащих жир или масло пищевых продуктов, фармацевтических препаратов и/или промышленных сырьевых материалов (например, сырьевых материалов для косметических препаратов, мыл и так далее) в соответствии со стандартной практикой.
[0043] Еще в одном варианте осуществления настоящее изобретение относится также к способу получения пищевого продукта, косметического препарата, фармацевтического препарата, мыла или тому подобное с использованием трансформированного липид-продуцирующего гриба или трансформированных дрожжей настоящего изобретения. Этот способ предусматривает стадию получения липидов или жирных кислот с использованием трансформированного липид-продуцирующего гриба или трансформированных дрожжей настоящего изобретения. Пищевой продукт, косметический препарат, фармацевтический препарат, мыло или тому подобное, каждые из которых содержат полученные липиды или жирные кислоты, могут быть приготовлены стандартным способом. Таким путем, такой пищевой препарат, косметический препарат, фармацевтический препарат, мыло или тому подобное, каждый из которых приготовлен способом по настоящему изобретению, содержит липиды или жирные кислоты, полученные с использованием трансформированного липид-продуцирующего гриба или трансформированных дрожжей по настоящему изобретению. Настоящее изобретение дополнительно обеспечивает изготовленный таким образом пищевой продукт, косметический препарат, фармацевтический препарат, мыло или тому подобное
[0044] Косметический препарат (композиция) или фармацевтический препарат (композиция) настоящего изобретения может находиться в любой лекарственной форме, такой как раствор, паста, гель, твердое вещество, порошок и другие лекарственные формы. Кроме того, эта косметическая композиция или фармацевтическая композиция настоящего изобретения может быть использована в косметических изделиях или наружных препаратах для кожи (например, масле, лосьоне, креме, эмульсии, геле, шампуне, кондиционере для волос, эмали для ногтей, тональном креме, губной помаде, пудре для лица, маске для лица, мази, духах, присыпке, одеколоне, зубной пасте, мыле, аэрозоле, очищающей пене), а также защитных и/или улучшающих агентах для стареющей кожи, защитных или улучшающих агентах для воспаления кожи, препаратах для ванн, усилителях роста волос, кожных эссенциях, солнцезащитных агентах или защитных и/или улучшающих агентах для кожных повреждений, вызываемых ранами, щелями или трещинами кожи, и так далее.
[0045] В случае необходимости, косметическая композиция по изобретению может быть дополнительно смешана подходящим образом с дополнительными ингредиентами, такими как жиры или масла, и/или красителями, ароматическими веществами, антисептиками, поверхностно-активными веществами, пигментами, антиоксидантами и так далее. Соотношение этих ингредиентов в смеси может определяться квалифицированным в данной области специалистом подходящим образом для предполагаемой цели (например, жиры или масла могут содержаться в этой композиции при соотношении 1%-99,99% по массе, предпочтительно 5%-99,99% по массе, более предпочтительно 10%-99,95% по массе). Подобным образом, фармацевтическая композиция настоящего изобретения может дополнительно содержать фармацевтически активные ингредиенты (например, противовоспалительный ингредиент) или вспомогательные ингредиенты (например, смягчающий ингредиент, ингредиент-носитель) в случае необходимости. Например, дополнительные ингредиенты, обычно используемые в косметике или наружных препаратах для кожи, включают лекарственные средства для лицевых угрей, защитные средства для перхоти и зуда, дезодоранты против потения, лекарственные средства для ожоговых ран, агенты против клещей и агенты против вшей, смягчители кератина, лекарственные средства для ксеродермы, антивирусные агенты, усилители проникающей через кожу абсорбции, и тому подобное.
[0046] Примеры пищевых продуктов изобретения включают питательные обогащенные пищевые продукты, диетические пищевые продукты, функциональные пищевые продукты, пищевые продукты для детей, модифицированное молоко для младенцев, модифицированное молоко для недоношенных младенцев, гериатрические пищевые продукты и тому подобное Термины “пища” или “пищевой продукт” используются здесь как общее название годных в пищу материалов в форме твердых веществ, жидкостей, напитков или их смесей.
Термин “питательные обогащенные пищевые продукты” относится к пищевым продуктам, обогащенным специфическими питательными ингредиентами. Термин “диетические пищевые продукты” относится к пищевым продуктам, которые являются оздоровительными или хорошими для здоровья, и включают питательные обогащенные пищевые продукты, природные пищевые продукты, диетические пищевые продукты и так далее. Термин “функциональные пищевые продукты” относятся к пищевым продуктам для восполнения питательных ингредиентов, которые способствуют контрольным функциям организма. Функциональные пищевые продукты являются синонимами пищевых продуктов для специального применения для здоровья. Термин “пищевые продукты для детей” относится к пищевым продуктам, даваемым детям до возраста приблизительно 6 лет. Термин “гериатрические пищевые продукты” относится к пищевым продуктам, обработанным для облегчения проглатывания и абсорбции, в сравнении с необработанными пищевыми продуктами. Термин “модифицированное молоко для младенцев” относится к модифицированному молоку, даваемому детям с возрастом приблизительно 1 года. Термин “модифицированное молоко для недоношенных младенцев” относится к модифицированному молоку, даваемому недоношенным младенцам до возраста приблизительно 6 месяцев от рождения.
[0047] Пища и пищевые продукты могут быть в форме природных пищевых продуктов (обработанных жиром или маслом), таких как мясо, рыба и орехи; пищевых продуктов, дополненных жирами или маслами во время приготовления (например, Китайских пищевых продуктов, Китайских лентовидных макаронных изделий, супов); пищевых продуктов, приготовленных с использованием жиров или масел в качестве теплоносителей (например, рыбы и овощей, запеченных в тесте), замороженных при низкой температуре пищевых продуктов, зажаренного соевого творога, Китайского зажаренного риса, пончиков, Японского зажаренного печенья (каринто); пищевых продуктов на основе жира или масла или обработанных пищевых продуктов, дополненных жирами или маслами во время обработки (например, маслом, маргарином, майонезом, заправкой, шоколадом, макаронными изделиями быстрого приготовления, карамельным кулером, бисквитами, печеньем, кексом, мороженым); и пищевых продуктов, опрысканных или покрытых жирами или маслами после окончательной отделки (например, рисовых крекеров, твердого печенья, пасты из вареных соевых бобов). Однако, они не ограничиваются пищевыми продуктами, содержащими жиры и масла, и другие примеры включают сельскохозяйственные пищевые продукты, такие как хлебобулочные изделия, макаронные изделия, вареный рис, сладости (например, конфеты, жевательные резинки, содержащие камеди конфеты, таблетки, Японские сладости), соевый творог и их обработанные продукты; ферментированные пищевые продукты, такие как Японское рисовое вино (саке), медицинский раствор, сладкий кулинарный херес (сладкая водка), уксус, соевый соус и мисо (ферментированная соевая паста из вареных соевых бобов и риса (соевая паста); пищевые продукты скота, такие как йогурт, ветчина, бекон и сосиски; морепродукты, такие как рыбная паста (камабоко), зажаренный рыбный пирог (агетен) и слоеный пирог с рыбой (ханпен); а также фруктовые напитки, безалкогольные напитки, напитки для спорта, алкогольные напитки, чай и тому подобное
[0048] Пищевой продукт по изобретению может также быть в форме фармацевтических препаратов, таких как капсулы, или в форме обработанных пищевых продуктов, таких как ординарные жидкие диеты, полуферментированные усиленные диеты, элементные диеты, пригодные для питья препараты или энтеральные питательные препараты, которые содержат жиры и липиды настоящего изобретения в смеси с белками, сахарами, жирами, микроэлементами, витаминами, эмульгаторами, ароматизаторами и так далее.
[0049] Как описано выше, при экспрессии в клетках-хозяевах, ген настоящего изобретения, ответственный за активность стимуляции элонгации цепей жирных кислот, делает возможной эффективную продуктивность липидов, в частности, триацилглицеринов.
Кроме того, уровень экспрессии этого гена может быть использован в качестве индикатора, например, в исследовании условий культивирования и/или обращении с этой культурой для гарантии эффективного продуцирования липидов, в частности, триацилглицеринов.
ПРИМЕРЫ
[0050] Настоящее изобретение будет ниже описано более подробно посредством следующих примеров, которые не предназначены для ограничения объема настоящего изобретения.
[0051] Геномный анализ Mortierella alpina
Штамм M. alpina 1S-4 инокулировали в 100 мл среды GY2:1 (2% глюкоза, 1% дрожжевой экстракт, pH 6,0) и культивировали при 28°C в течение 2 дней в условиях встряхивания. Эти клетки собирали фильтрованием и их геномную ДНК получали с использованием DNeasy (QIAGEN). Нуклеотидную последовательность приведенной выше геномной ДНК определяли с использованием Roche 454 GS FLX Standard, во время которого секвенирование нуклеотидов проводили в двух экспериментах для библиотеки фрагментов и в трех экспериментах для библиотеки спаренных нуклеотидов. Полученные нуклеотидные последовательности собирали с получением 300 суперконтигов.
[0052] Синтез кДНК и получение библиотеки кДНК
Штамм M. alpina 1S-4 инокулировали в 100 мл среды (1,8% глюкоза, 1% дрожжевой экстракт, pH 6,0) и предварительно культивировали в течение 3 дней при 28°C. 10-литровый культуральный сосуд (Able Co., Tokyo) загружали 5 л среды (1,8% глюкоза, 1% порошок сои, 0,1% оливковое масло, 0,01% Адеканол, 0,3% KH2PO4, 0,1% Na2SO4, 0,05% CaCl2·2H2O, 0,05% MgCl2·6H2O, pH 6,0) и инокулировали со всем предварительно культивированным продуктом, с последующим культивированием в аэробной спин-культуре в условиях 300 об/мин, 1 vvm и 26°C в течение 8 дней. В дни 1, 2 и 3 культивирования добавляли глюкозу в количестве, соответствующем 2%, 2% и 1,5%, соответственно. Эти клетки собирали на каждой стадии культивирования (в день 1, 2, 3, 6 или 8) для получения общей РНК гуанидингидрохлорид/CsCl-способом. С использованием набора для очистки мРНК Oligotex-dT30
[0053] Поиск на гомологи дрожжевого AYR1
Гомологи ScAYR1 (YIL124W), который является геном, ответственным за активность 1-ацил-дигидроксиацетон-фосфатредуктазы и активность β-кетоацилредуктазы в дрожжах, подвергали поиску против геномных баз данных. В результате, “хит” был найден в суперконтиге, содержащем последовательность SEQ ID NO: 4. Этот ген SEQ ID NO: 4 был назван MaADR1.
[0054] Клонирование кДНК MaADR1
Для клонирования кДНК для гена MaADR1, было предсказано, что ATG, локализованные в положениях 1-3 SEQ ID NO: 4, могли бы быть инициирующим кодоном, тогда как положения 1587-1589 SEQ ID NO: 4 могли бы составлять терминирующий кодон, как можно было судить по присутствию инициирующего и терминирующего кодонов, а также по сравнению последовательности с этим гомологом. Затем синтезировали следующие праймеры.
Bam-ADR-F: 5'-GGATCCATGGCCTCGTCTAAAAAGATCGTCCT-3' (SEQ ID NO: 5)
Sal-ADR-R: 5'-GTCGACTACTTTCCAACGACCTTGCCATCC-3' (SEQ ID NO: 6)
С использованием кДНК штамма M. alpina 1S-4 в качестве матрицы, ПЦР-амплификацию проводили посредством KOD-Plus (TOYOBO) с этими праймерами Bam-ADR-F и Sal-ADR-R, посредством чего был амплифицирован ДНК-фрагмент приблизительно 0,87 т.п.н. Этот фрагмент клонировали с использованием набора для клонирования Zero Blunt TOPO PCR (Invitrogen) и полученная плазмида была названа pCR-MaADR1. Последовательность этого инсерта в этой плазмиде, т.е. CDS-последовательность гена MaADR1 показана в SEQ ID NO: 3. Кроме того, ORF-последовательность гена MaADR1 показана в SEQ ID NO: 1.
[0055] Анализ последовательности
Когда было проведено сравнение между геномной последовательностью (SEQ ID NO: 4) и CDS-последовательностью (SEQ ID NO: 3) гена MaADR1, было обнаружено, что геномная последовательность этого гена содержит пять экзонов и четыре интрона (Фигура 1) и было предсказано, что она кодирует белок, состоящий из 242 аминокислотных остатков (Фигура 2).
Эту расшифрованную аминокислотную последовательность (SEQ ID NO: 2) MaADR1 подвергали анализу гомологии BLASTp против аминокислотных последовательностей, зарегистрированных в GENEBANK nr. В результате, аминокислотная последовательность, обнаруживающая самую низкую E-величину против этой последовательности, т.е. имеющая наивысшую идентичность с этой последовательностью, была предположительным белком, произведенным из Volvox carterif. nagariensis (зеленой водоросли) (GENEBANK accession No. XP_002946364), которая на 34,7% идентична аминокислотной последовательности. Кроме того, эта расшифрованная аминокислотная последовательность MaADR1 на 25,6% идентична аминокислотной последовательности AYR1p, полученной из дрожжей S. cerevisiae, и на 13,6% идентична аминокислотной последовательности IFA38.
Фигура 3 показывает сравнение аминокислотной последовательности MaADR1, предположительного белка, полученного из Volvox carterif. nagariensis (зеленой водоросли) (GENEBANK accession No. XP_002946364), и AYR1p, полученного из S. cerevisiae.
[0056] Функциональный анализ MaADR1
Конструирование экспрессирующего вектора дрожжей
Экспрессирующий вектор дрожжей, pYE22m (Biosci. Biotech. Biochem., 59, 1221-1228, 1995), расщепляли рестрикционными ферментами BamHI и SalI для получения ДНК-фрагмента, тогда как плазмиду pCR-MaADR1 также расщепляли рестрикционными ферментами BamHI и SalI с получением ДНК-фрагмента приблизительно 0,87 т.п.н. Эти ДНК-фрагменты лигировали вместе с использованием ligation high (TOYOBO) для конструирования плазмиды pYE-MaADR1.
[0057] Получение трансформированных дрожжей
Плазмиды pYE22m и pYE-MaADR1, каждую, использовали для трансформации штамма дрожжей S. cerevisiae EH13-15 (trp1, MATα) (Appl. Microbiol. Biotechnol., 30, 515-520, 1989) литий-ацетатным способом. Эти трансформированные штаммы подвергали скринингу по способности расти на SC-Trp-агаровой среде (2% агар), содержащей, на литр, 6,7 г азотистого основания дрожжей без аминокислот (DIFCO), 20 г глюкозы и 1,3 г порошка аминокислот (смеси 1,25 аденинсульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 1,8 г лейцина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина и 0,6 г урацила).
[0058] Культура дрожжей
Любые четыре штамма, полученные трансформацией плазмидой pYE22m, и любые четыре штамма, полученные трансформацией плазмидой pYE-MaADR1, подвергали следующему варианту культивирования. А именно, на стадии пре-культивирования, трансфер-петлю каждого дрожжевого штамма инокулировали из планшета в SC-Trp-среду (10 мл) и культивировали при 30°C в течение 1 дня в условиях встряхивания. В стадии основного культивирования, этот пре-культивированный раствор (100 мкл) добавляли в SC-Trp-среду (10 мл) и культивировали при 30°C в течение 2 дней в условиях встряхивания.
[0059] Анализ жирных кислот клеток дрожжей
Эти культивированные растворы дрожжей, каждый, центрифугировали для сбора этих клеток. После промывания 10 мл стерилизованной воды эти клетки собирали опять центрифугированием и затем лиофилизировали. К этим лиофилизированным клеткам добавляли 1 мл смеси хлороформ:метанол (2:1) и стеклянные бусины и эти клетки гомогенизировали с разбивателем бусин, с последующим центрифугированием для сбора супернатанта. К оставшимся клеткам добавляли опять 1 мл смеси хлороформ:метанол (2:1) и супернатант собирали таким же образом. Эту процедуру повторяли для сбора липидов со смесью хлороформ:метанол (2:1) в общем объеме 4 мл. Этот растворитель отгоняли с использованием SpeedVac. Каждую из этих проб растворяли в 1 мл хлороформа.
Брали аликвоту 200 мкл каждой пробы и обрабатывали способом с хлористоводородной кислотой-метанолом для превращения жирных кислот в соответствующие метиловые эфиры с последующей газовой хроматографией для анализа жирных кислот для определения состава общих жирных кислот в этих клетках.
[0060] Полученные результаты показаны в таблице 1. Штамм, высоко экспрессирующий ген MaADR1 показал увеличенную долю С18-жирных кислот и уменьшенную долю С16-жирных кислот в общих жирных кислотах в сравнении с контрольным штаммом. А именно, активировалась реакция элонгации цепей жирных кислот.
[Таблица 1]
[0061] Брали аликвоту 400 мкл из каждой из вышеуказанных проб. После отгонки растворителя, остаток растворяли в малом объеме хлороформа и подвергали тонкослойной хроматографии. А именно, в условиях, использующих планшет из силикагеля 60 (Merck & Co., Inc.) и проявляющий растворитель, гексан:диэтиловый эфир:уксусная кислота (70:30:1), выполняли тонкослойную хроматографию для фракционирования липидов. Этот планшет опрыскивали раствором примулина и облучали ультрафиолетовым светом для детектирования липидов. Фракцию триацилглицерина (TG) и фракцию фосфолипида (PL) выскребали в отдельные тест-пробирки и обрабатывали по способу с хлористоводородной кислотой:метанолом для превращения жирных кислот в соответствующие метиловые эфиры с последующей газовой хроматографией для анализа жирных кислот.
[0062] Таблицы 2 и 3 показывают состав жирных кислот во фракции триацилглицерина и состав жирных кислот в фосфолипидной фракции, соответственно.
[Таблица 2]
[Таблица 3]
[0063] При фокусировании внимания на составе жирных кислот во фракции триацилглицерина, этот штамм, высоко экспрессирующий MaADR1, показал увеличенную долю С18-жирных кислот и уменьшенную долю С16 жирных кислот в сравнении с контролем. С другой стороны, при фокусировании внимания на составе жирных кислот в фосфолипидной фракции, этот штамм, высоко экспрессирующий MaADR1, и контрольный штамм показали одинаковые доли жирных кислот.
А именно, когда MaADR1 высоко экспрессировался в клетках дрожжей, состав жирных кислот, составляющих триацилглицерины, изменялся таким путем, чтобы увеличивать долю жирных кислот с более длинной цепью.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
[0064] При экспрессии в подходящих клетках-хозяевах полинуклеотид настоящего изобретения позволяет осуществлять эффективное продуцирование жирных кислот с длинной цепью, имеющих 18 или более атомов углерода, и триацилглицеринов, содержащих их. Жирные кислоты, продуцируемые в клетках-хозяевах посредством настоящего изобретения, могут быть использованы для получения пищевых продуктов, косметических препаратов, фармацевтических препаратов, мыла, и так далее.
Свободный текст списка последовательностей
[0065] SEQ ID NO: 5: синтетическая ДНК
SEQ ID NO: 6: синтетическая ДНК
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SUNTORY HOLDINGS LIMITED
<120> Белок, обладающий активностью в отношении стимуляции элонгации
цепей жирных кислот, кодирующий его ген и их применение
<130> PCT12-0028
<150> JP 2011-171044
<151> 2011-08-04
<160> 6
<170> PatentIn version 3.5
<210> 1
<211> 726
<212> ДНК
<213> Mortierella alpina
<400> 1
atggcctcgt ctaaaaagat cgtcctcgtc accggctgta ccactggagg cattggttat 60
gaaaccgcaa aggcattcga aaagagtggc tgcaaagtgt atgccgcagc aagacgtctc 120
gaagccataa cgggcattga aggtctagat atcgaaaagg tctacatcga cgtactggac 180
gagaagtcca tcaaagacgc cgtcaacatt gagacaaccc gcaagctgct cgacaccaac 240
atcacctccg tcattctcgt gtccaaagag gtggcgcctc atatgattag acaaaagtct 300
ggtctgattg tcaatgttgg ctcagtcaca gcctatctcg cgacaccttg gggcggtctc 360
tatgctgcca gcaaggccgc agtgcactcc atctcggacg cactgcgcat ggagttggct 420
ccctttggtg ttgatgtttc ggtcgtggcg cctggtgcaa tcaagtccaa catcggtgac 480
aacaacttga aggccttcca tcttcccgag acccctggat gcacacccac tgccaagttt 540
gcaaagtacg tcgtggcaaa gtgcctcaag tcatcccccc ctcgatacat cgattacggc 600
acgctgtcaa acctcttccg attcttgcgc tacgcgccct ggatgatcac ggacttcatc 660
ttctcccgca aatttggtct gaatgttctc cagaagtcgg taaaggatgg caaggtcgtt 720
ggaaag 726
<210> 2
<211> 242
<212> БЕЛОК
<213> Mortierella alpina
<400> 2
Met Ala Ser Ser Lys Lys Ile Val Leu Val Thr Gly Cys Thr Thr Gly
1 5 10 15
Gly Ile Gly Tyr Glu Thr Ala Lys Ala Phe Glu Lys Ser Gly Cys Lys
20 25 30
Val Tyr Ala Ala Ala Arg Arg Leu Glu Ala Ile Thr Gly Ile Glu Gly
35 40 45
Leu Asp Ile Glu Lys Val Tyr Ile Asp Val Leu Asp Glu Lys Ser Ile
50 55 60
Lys Asp Ala Val Asn Ile Glu Thr Thr Arg Lys Leu Leu Asp Thr Asn
65 70 75 80
Ile Thr Ser Val Ile Leu Val Ser Lys Glu Val Ala Pro His Met Ile
85 90 95
Arg Gln Lys Ser Gly Leu Ile Val Asn Val Gly Ser Val Thr Ala Tyr
100 105 110
Leu Ala Thr Pro Trp Gly Gly Leu Tyr Ala Ala Ser Lys Ala Ala Val
115 120 125
His Ser Ile Ser Asp Ala Leu Arg Met Glu Leu Ala Pro Phe Gly Val
130 135 140
Asp Val Ser Val Val Ala Pro Gly Ala Ile Lys Ser Asn Ile Gly Asp
145 150 155 160
Asn Asn Leu Lys Ala Phe His Leu Pro Glu Thr Pro Gly Cys Thr Pro
165 170 175
Thr Ala Lys Phe Ala Lys Tyr Val Val Ala Lys Cys Leu Lys Ser Ser
180 185 190
Pro Pro Arg Tyr Ile Asp Tyr Gly Thr Leu Ser Asn Leu Phe Arg Phe
195 200 205
Leu Arg Tyr Ala Pro Trp Met Ile Thr Asp Phe Ile Phe Ser Arg Lys
210 215 220
Phe Gly Leu Asn Val Leu Gln Lys Ser Val Lys Asp Gly Lys Val Val
225 230 235 240
Gly Lys
<210> 3
<211> 729
<212> ДНК
<213> Mortierella alpina
<400> 3
atggcctcgt ctaaaaagat cgtcctcgtc accggctgta ccactggagg cattggttat 60
gaaaccgcaa aggcattcga aaagagtggc tgcaaagtgt atgccgcagc aagacgtctc 120
gaagccataa cgggcattga aggtctagat atcgaaaagg tctacatcga cgtactggac 180
gagaagtcca tcaaagacgc cgtcaacatt gagacaaccc gcaagctgct cgacaccaac 240
atcacctccg tcattctcgt gtccaaagag gtggcgcctc atatgattag acaaaagtct 300
ggtctgattg tcaatgttgg ctcagtcaca gcctatctcg cgacaccttg gggcggtctc 360
tatgctgcca gcaaggccgc agtgcactcc atctcggacg cactgcgcat ggagttggct 420
ccctttggtg ttgatgtttc ggtcgtggcg cctggtgcaa tcaagtccaa catcggtgac 480
aacaacttga aggccttcca tcttcccgag acccctggat gcacacccac tgccaagttt 540
gcaaagtacg tcgtggcaaa gtgcctcaag tcatcccccc ctcgatacat cgattacggc 600
acgctgtcaa acctcttccg attcttgcgc tacgcgccct ggatgatcac ggacttcatc 660
ttctcccgca aatttggtct gaatgttctc cagaagtcgg taaaggatgg caaggtcgtt 720
ggaaagtag 729
<210> 4
<211> 1589
<212> ДНК
<213> Mortierella alpina
<400> 4
atggcctcgt ctaaaaagat cgtcctcgtc accggctgta ccactggagg cattggttat 60
gaaaccgcaa aggcattcga aaaggtacgc cctcggcagt ctcacttcat ggaaggcccc 120
tctgagaatt accaatcaac caatttgcag ttacagctgg attcattgac taatcgtgca 180
tacctttctt atactaaatg cctattcaga gtggctgcaa agtgtatgcc gcagcaagac 240
gtctcgaagc cataacgggc attgaaggtg agggttgtat tttgccacaa gtcttcgtgg 300
gtcgtgtgcg ggatgcaggc attaacactc cttaaattgc gtgcaggtct agatatcgaa 360
aaggtctaca tcgacgtact ggacgagaag tccatcaaag acgccgtcaa cgtaagatgc 420
ctgctgtcaa ctgtcctact tctacttgca taagttttca attctgatct ctcaggtcct 480
taaacttgca ttgtagcacg ttatcgagaa ggaaggacga atcggtaaga gaagaaacgc 540
gtgttttcag ttcaacggac gattgctcaa catgcaagaa gaccaagcat tgatggctgc 600
cttatattct tcatacagat attctgttca acaatgccgg aatgggactc gcatgcccac 660
tgatcgacat gtctgtaagt aacacagggt ggacatatga acactgaaag gcaaacccca 720
ccttagcaga cggcaagcac taacacttca gccttcattt aatatgtata gattgagaca 780
acccgcaagc tgctcgacac caacatcacc tccgtcattc tcgtgtccaa agaggtggcg 840
cctcatatga ttagacaaaa gtctggtctg attgtcaatg ttggctcagt cacagcctat 900
ctcgcgacac cttggggcgg tctctatgct gccagcaagg ccgcagtgca ctccatctcg 960
gacgcactgc gcatggagtt ggctcccttt ggtgttgatg tttcggtcgt ggcgcctggt 1020
gcaatcaagt ccaacatcgg tgacaacaac ttgaaggcct tccatcttcc cgagagtaag 1080
ttcaaccagc aattccgtcc gcttgaagct gcaatcatat tcccttcagc caaattcctc 1140
atatgctcat actgccttgt tatttttttt ttttccttct ttgttgaaat ctcaagattc 1200
cttctatcag tctgtcatca gctatatcat gtccagagca aatgcttccc aaggtaagac 1260
gcgtcgattt cactgacgat ctgccaacaa tggaaaaaaa accgttgcgt cttgatgcta 1320
ctgacttgtg tctctgacac tcactgtgcc cccatcttgt ttatccatca gcccctggat 1380
gcacacccac tgccaagttt gcaaagtacg tcgtggcaaa gtgcctcaag tcatcccccc 1440
ctcgatacat cgattacggc acgctgtcaa acctcttccg attcttgcgc tacgcgccct 1500
ggatgatcac ggacttcatc ttctcccgca aatttggtct gaatgttctc cagaagtcgg 1560
taaaggatgg caaggtcgtt ggaaagtag 1589
<210> 5
<211> 32
<212> ДНК
<213> Искусственная
<220>
<223> Синтетическая ДНК
<400> 5
ggatccatgg cctcgtctaa aaagatcgtc ct 32
<210> 6
<211> 30
<212> ДНК
<213> Искусственная
<220>
<223> Синтетическая ДНК
<400> 6
gtcgactact ttccaacgac cttgccatcc 30
Реферат
Группа изобретений относится к биотехнологии, в частности к белку, обладающему активностью в отношении стимуляции элонгации цепей жирных кислот, и полинуклеотиду, кодирующему этот белок. Предложены также вектор экспрессии, содержащий полинуклеотид, и трансформант, полученный из S. cerevisiae, для продукции указанного белка, содержащий полинуклеотид либо вектор. Группа изобретений обеспечивает эффективное продуцирование жирных кислот с длинной цепью, имеющих 18 атомов углерода. 5 н. и 4 з.п. ф-лы, 3 ил., 3 табл.
Формула
(a) полинуклеотид, состоящий из нуклеотидной последовательности SEQ ID NO:1 или 4;
(b) полинуклеотид, кодирующий белок, который состоит из аминокислотной последовательности SEQ ID NO:2;
(c) полинуклеотид, кодирующий белок, который состоит из аминокислотной последовательности с делецией, заменой, инсерцией и/или добавлением от 1 до 10 аминокислот в аминокислотной последовательности SEQ ID NO:2;
(d) полинуклеотид, кодирующий белок с аминокислотной последовательностью, которая на 90% или более идентична аминокислотной последовательности SEQ ID NO:2; и
(e) полинуклеотид, который гибридизируется в строгих условиях с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности SEQ ID NO:1 или 4.
Комментарии