Способы получения растворимых многопроходных трансмембранных белков - RU2441071C2
Код документа: RU2441071C2
Чертежи





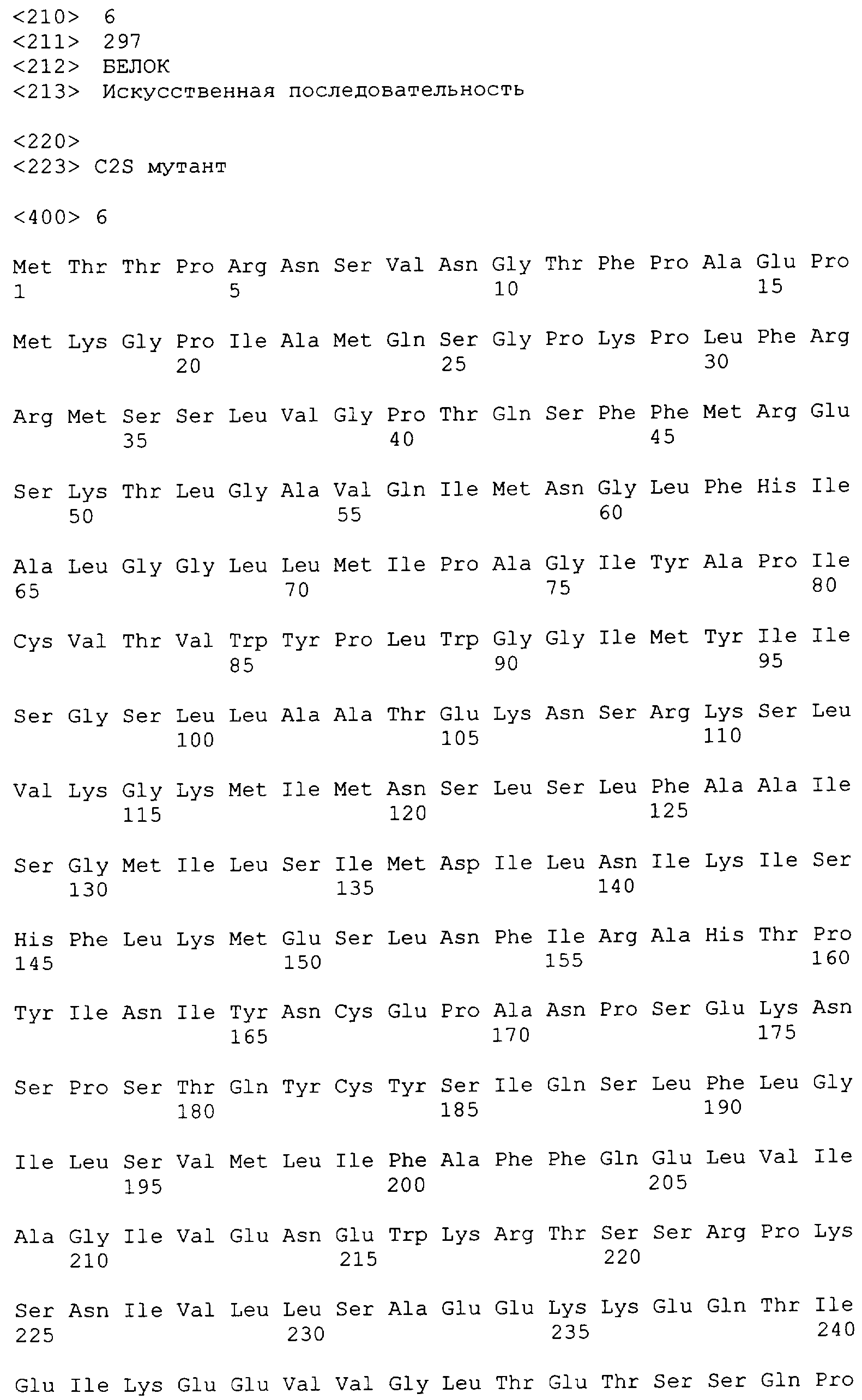






































Описание
Предпосылки создания изобретения
Сложные трансмембранные белки трудно экспрессировать в клетках-хозяевах. Обычно эти белки токсичны для хозяина, а различные системы экспрессии дают низкие выходы экспрессированного белка. Кроме того, эти трансмембранные полипептиды трудно солюбилизировать, агрегация и денатурация затрудняют получение белкового продукта удовлетворительного качества и в количестве, достаточном для эффективного применения.
Примером трансмембранного белка является подсемейство с 4 доменами семейства гена A (MS4A), которое включает CD20, β цепь высокоаффинного рецептора IgE, HTm4 и т.п. Эти белки являются структурно родственными, по меньшей мере, в 4 трансмембранных доменах клеточной поверхности (Ishibashi et al., 2001, Gene 264: 87-93). Хотя интервалы идентичности всей аминокислотной последовательности в полипептидах семейства MS4A составляют 25-40%, аминокислоты первых трех трансмембранных доменов имеют более высокую степень идентичности и гомологии, чем полипептид в целом (Ishibashi et al., 2001, Supra; Liang el al., 2001, Genomics 72: 119-127). Структурно полипептиды MS4A также имеют общий мотив внеклеточной петли. Как N-, так и С-конец полипептида MS4A обнаружены на стороне цитоплазмы клеточной мембраны (Ishibashi et al., 2001, Supra). N- и С-концы выявляют значительно более высокую дивергенцию последовательностей между полипептидами семейства гена MS4A (Ishibashi el al., 2001, Supra).
Несмотря на значительное структурное сходство, полипептиды семейства гена MS4A неодинаково экспрессируются в отдельных типах клеток (Liang et al., 2001, Supra). CD20 экспрессируется исключительно в В клетках (Stashenko et al., 1980, J. Immunol, 125: 1678-1685). β цепь высокоаффинного рецептора IgE (FcεRIβ) экспрессируется исключительно в тучных клетках и базофилах (Kinet, 1999, Annu. Rev. Immunol, 17: 931-972). FcεRIβ связывает IgE и опосредует внутриклеточную передачу сигнала (т.е. дегрануляцию), инициированную связыванием антигена (Dombrowicz et al., 1996, Immunity. 8: 517-529; Lin et al., 1996, Cell, 85:985-995). HTm4 экспрессируется в гемопоэтической ткани и служит в качестве регулятора клеточного цикла гемопоэтических клеток (Donato et al., 2002, J. Clin. Invest., 109: 51-58). Эти белки имеют общее свойство, сложную структуру трансмембранных белков. Это свойство очень затрудняет экспрессию в клетке-хозяине и солюбилизацию при использовании клеточной мембраны в "нативной" конфигурации.
Трансмембранные полипептиды, например CD20, являются потенциальными мишенями для терапевтических средств при лечении заболеваний, таких как рак и аутоиммунные заболевания. Первоначально CD20 был идентифицирован в качестве маркера для В клеток более 20 лет назад, а в настоящее время установлено, что в качестве маркера он присутствует в большинстве В клеточных лимфом. CD20 является мишенью для терапии с помощью моноклональных антител при лечении неходжкинской лимфомы (NHL), и, в частности, мишенью для химерного антитела ритуксимаба (RTTUXAN®), ведущего лекарственного препарата при лечении NHL. Ритуксимаб распознает CD20, экспрессирующийся на В клетках. Связывание ритуксимаба зависит от конформации, он связывается с CD20 в зависимости от структуры петли между третьим и четвертым трансмембранными спиральными доменами, содержащими цистеиновые остатки в положениях 167 и 183.
Значительное препятствие в создании терапевтических препаратов, нацеленных на трансмембранные полипептиды, такие как CD20, представляет неспособность продуцировать достаточные количества этих полипептидов в клетках-хозяевах, в частности в бактериальных клетках, и неспособность получать очищенные рекомбинантные или природные трансмембранные полипептиды в нативной конформации. Необходимы способы получения и солюбилизации природных/или рекомбинантных трансмембранных полипептидов в нативной конформации.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящее время найдено, что трансмембранные полипептиды, например, состоящие из единичного трансмембранного домена или множества трансмембранных доменов, можно эффективно продуцировать в бактериальных клетках-хозяевах и солюбилизировать при использовании клеточных мембран с хорошим выходом и с адекватной нативной конформацией для того, чтобы их можно было использовать в качестве иммуногенов и лигандов, например, в количественном анализе. Трансмембранные полипептиды можно продуцировать, выделять и солюбилизировать способами по данному описанию, с хорошим выходом и с подходящей "нативной" конформацией.
Способы продуцирования трансмембранных полипептидов включают экспрессию в клетках, например в бактериальных клетках, под контролем активного строго регулируемого промотора, например, phoA промотора в Е.coli. В одном варианте изобретения строго регулируемый промотор содержит как элемент позитивного контроля, так и элемент негативного контроля и может содержать ряд таких элементов. Промотор может быть мутантным промотором, например, таким, в который встроен гетерологичный элемент позитивного или негативного контроля. Кроме того, промотор может содержать терминаторы транскрипции, например терминаторы транскрипции лямбда, позиционированные таким образом, чтобы предупредить возможное считывание возможного "апстрим" промотора. Для экспрессии белка в Е.coli промотор может являться, например, phoA или его мутациями, содержащими дополнительные элементы негативного контроля, такими как phac и tphac, мутантные промоторы, раскрываемые ниже, в разделе Примеры, которые содержат дополнительный оператор lac.
Векторы для экспрессии трансмембранных полипептидов включают полинуклеотидную последовательность, кодирующую трансмембранный полипептид под контролем строго регулируемого промотора. Такие полипептиды включают, например, полипептиды, имеющие, по меньшей мере, четыре трансмембранных домена, такие как CD20 и мутантный C2S-CD20, раскрываемый ниже, в разделе Примеры. Кодируемые полипептиды могут иметь один, два, три, четыре, пять, шесть, семь, восемь, девять, десять, одиннадцать, двенадцать или двадцать четыре или более трансмембранных домена. Другие примеры включают рецептор EG-VEGF, GPR73, имеющий семь трансмембранных доменов, бета цепь высокоэффективного IgE рецептора (FcεRIβ), HTm4, MS4A4A, MS4A6, MS4A7 и RA1c. Вектор может также включать гены тРНК с редкими кодонами бактериальной клетки-хозяина, и/или полинуклеотидную последовательность, прилегающую непосредственно к первому кодону, кодирующую лидерный пептид, для стимуляции инициации трансляции. Лидерная последовательность обычно содержит последовательность инициации трансляции (TIS) и спейсер для эффективной элонгации. Последовательность инициации трансляции в данном описании называют TIS, но ее также называют областью инициации трансляции (ТIR).
В одном варианте изобретения лидерная последовательность содержит активную TIS, кодирующую, по меньшей мере, часть trp лидерной последовательности, например около 6-12 аминокислот. Спейсерная последовательность отделяет последовательность инициации трансляции от первой трансмембранной области и обычно кодирует малый внутренний участок белка, известного как хорошо экспрессирующийся в клетке-хозяине, например, такого как "Е" белок в Е.coli. Спейсерная последовательность обычно неструктурирована и в значительной степени гидрофильна.
В одном варианте изобретения вектор для экспрессии растворимых мультитрансмембранных белков содержит строго регулируемый промотор, такой как phoA промотор или его мутант, элементы негативного и/или позитивного контроля и полинуклеотидную последовательность, кодирующую лидерную последовательность, включающую активную последовательность инициации трансляции и спейсерную последовательность для элонгации трансляции, позиционированную между TIS и первой трансмембранной областью белка.
Например, векторы могут содержать phoA, phac или tphac промотор, элемент негативного контроля, такой как оператор lac, лидерную последовательность, кодирующую последовательность инициации трансляции, например, участок trp лидера, такой как аминокислотная последовательность из девяти аминокислот KAIFVLKGS, и спейсерную последовательность, кодирующую последовательность трансляции элонгации, такую как участок гена trp Е, например, найденный в LE лидерной последовательности (SEQ ID NO: 25) или sLE лидерной последовательности (SEQ ID NO: 26) по данному описанию.
Трансмембранные полипептиды можно собирать и очищать от мембран клетки-хозяина солюбилизацией в детергенте. В одном варианте изобретения для солюбилизации трансмембранных полипептидов используют неионные или цвиттерионные детергенты, такие как н-додецилфосфохолин, DDPC. Выделенные мультитрансмембранные полипептиды, такие как CD20, растворимы в этих детергентах. Выделенные растворимые мультитрансмембранные полипептиды содержат адекватную (удовлетворительную) "нативную" структуру для распознавания антителами, которые узнают полипептиды, экспрессирующиеся на поверхности клеток, и применимы в качестве иммуногенов и лигандов для анализа.
КРАТКОЕ ОПИСАНИЕ ФИГУР
На Фигуре 1 в виде диаграммы представлен CD20, присутствующий на В клеточной мембране. Последовательность и предполагаемая общая топология CD20 представлены относительно мембраны клеточной поверхности.
На Фигуре 2 показан Вестерн-блоттинг меченного His CD20 с применением антител к His-метке. На панели показаны: а) фракции, содержащие CD20, после флотации клеточных мембран клеток Е.coli в градиенте (плотности) сахарозы. Аликвоты фракций (указанных номерами наверху) градиента сахарозы подвергают электрофорезу на SDS-PAGE геле. Наносят пятно геля и анализируют с помощью антитела к His метке. Показаны фракции от самой низкой плотности сахарозы до самой высокой. На панели b) показаны уровни His-меченного CD20 на Вестерн-блоте после экстракции Е.coli мембран различными детергентами. После экстракции различными детергентами метят супернатанты (S) и осадок (пеллеты) (Р). (WC) обозначает экстракт цельных клеток (контроль). Цифры 1-7 обозначают различные тестируемые детергенты, они представляют собой SDS (1), н-лаурилсаркозин (2), н-додецил-N,N-диметиламин-N-оксид (LADO) (3), н-додецилфосфохолин (DDPC) (4), н-додецил-β-D-мальтозид (DDM) (5), Трилон-Х 100 (6) и CHAPS (7). На панели с) показаны уровни меченного His полипептида CD20, детектированные Вестерн-блоттингом с применением антитела к метке His, для клеток Е.coli, экспрессирующих меченные His нативный человеческий CD20 и C2S мутантный CD20. Дорожки 1 и 4 изображают контрольный, холостой вектор, дорожки 2 и 5 изображают меченный His человеческий CD20, а дорожки 3 и 6 изображают C2S мутантный CD20. Образцы на дорожках 1, 2 и 3 тестируют в невосстанавливающих условиях; образцы на дорожках 4, 5 и 6 восстанавливают с помощью 100 мМ DTT. Все дорожки содержат равный объем клеток, нормализованный по оптической плотности.
На Фигуре 3 показаны дорожки с окрашенным Кумасси синим SDS гелем очищенного, меченного His человеческого CD20, C2S мутантного и мышиного CD20. Дорожки 1, 2 и 3 панели а) содержат невосстановленные белки: человеческий CD20 (дорожка 1), C2S мутантный (дорожка 2) и мышиный CD20 (дорожка 3). Дорожка 4 содержит маркеры молекулярной массы (Mark 12, Invitrogen). На дорожках 5, 6 и 7 показаны восстановленные белки: человеческий CD20 (дорожка 5), C2S мутантный (дорожка 6) и мышиный CD20 (дорожка 7). На дорожках 8 и 10 показаны невосстановленный и восстановленный очищенные мышиные CD20 соответственно. На дорожке 9 представлены маркеры молекулярной массы. Каждая дорожка содержит 2 мкг (микрограмма) белка. Молекулярные массы маркеров 200, 116, 97, 66, 55, 36, 30, 22, 14 и 6 кДа.
На Фигуре 4 изображен график, показывающий дисульфид-зависимое связывание антитела ритуксимаба с меченным His человеческим CD20 (закрашенные квадраты), восстановленным и алкилированным hCD20 (закрашенные кружки), восстановленный и снова окисленный hCD20 (незакрашенные квадраты) и PBS контроль (незакрашенные кружки).
На Фигуре 5 представлена BIAcore сенсограмма, изображающая связывание между ритуксимабом и меченным His человеческим CD20. Связывание человеческого CD20 с иммобилизованным ритуксимабом происходит при концентрациях CD20 5 мкМ, 2.5 мкМ, 1.25 мкМ, 0.63 мкМ, 0.31 мкМ, 0.16 мкМ, 0.08 мкМ и 0.04 мкМ. Концентрации первых 4 образцов помечены на сенсограмме. Расчетная теоретическая кривая для неассоциированной модели показана для каждой концентрации.
На Фигуре 6 показаны спектры кругового дихроизма белков CD20 в дальней ультрафиолетовой области. На панели (а) показаны спектры C2S мутанта человеческого CD20 в присутствии 0.1% DDPC (черная линия); в 0.1% DDPC и 10 мМ (3-меркаптоэтанола (пунктирная линия), и после термального сканирования до 95°С в присутствии 1% SDS (серая линия). На панели (b) показаны спектры мышиного CD20 в присутствии 0.1% DDPC (пунктирная линия), 0.1% додецилмальтозида (DDM) (серая линия); и в 0.1% DDM с добавлением 1% SDS и β-меркаптоэтанола и после нагревания в течение 2 минут при 95°С (черная линия). Данные выражаются в виде молярной эллиптичности.
На Фигуре 7 показаны кривые замещения ритуксимаба IgG и Fab с выделенными нормальными В клетками. Величина ЕС50 для нативного CD20 в данном анализе равна 9.5 нМ. Связывание определяют конкурентной реакцией немеченого ритуксимаба IgG к125I-IgG за донор 1 (панель а) или немеченого ритуксимаба Fab к125I-Fab за донор 4 (панель b). См. в Таблице 4 величины аффинностей и число рецепторов от каждого донора.
На Фигуре 8 показана конструкция вектора экспрессии и Вестерн-блот, показывающий экспрессию полипептидов семейства MS4A в Е.coli, включая полипептиды MS4A6A, MS4A7 и MS4A4A.
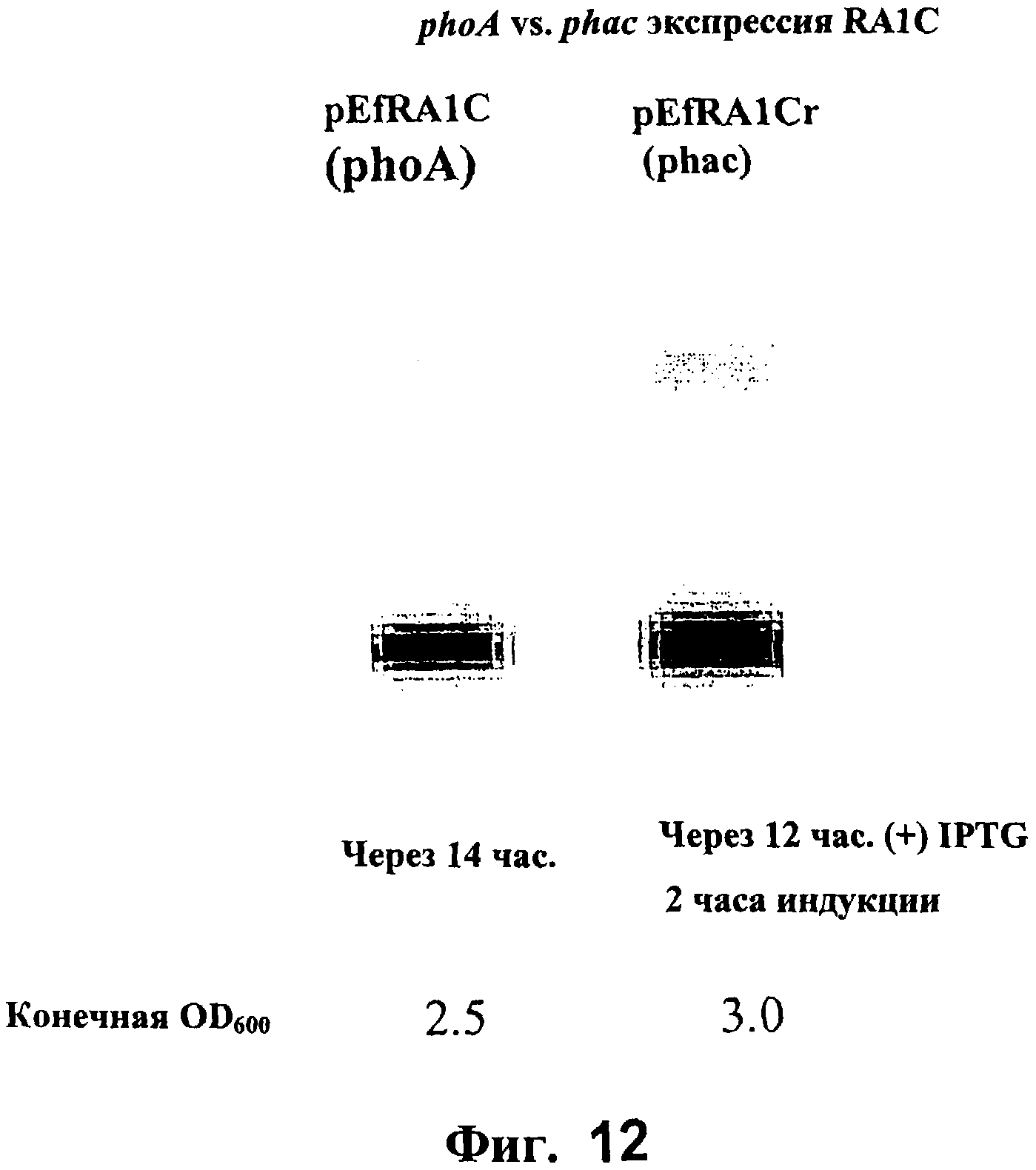
На Фигуре 9 изображен Вестерн-блот, показывающий полипептид RA1c, экспрессирующийся вследствие неполного блокирования при использовании неиндуцированного phoA промотора (pEfRA1c) и мутантного промотора, phac (pEfRA1Cr), детектируемый с помощью антитела к His метке.
На Фигуре 10 показан Вестерн-блот, изображающий продолжительность экспрессии полипептида RA1c при использовании промотора phoA (pEfRA1C), индуцируемого разведением в средах, лимитированных по фосфатам.
На Фигуре 11 показан Вестерн-блот, изображающий продолжительность экспрессии полипептида RA1c при использовании промотора phac (pEfRA1Cr), индуцируемого разведением в лимитированных по фосфатам средах и добавлением IPTG.
На Фигуре 12 представлен Вестерн-блот, сравнивающий экспрессию RA1c при использовании индуцированных промоторов phoA и phac.
На Фигуре 13 представлен Вестерн-блот, показывающий EG-VEGF рецептор, полипептид GPR73, экспрессирующийся в Е.coli вследствие частичного блокирования при использовании неиндуцированного phoA промотора (средняя дорожка) и мутантного промотора, phac (правая дорожка).
На Фигуре 14 представлен Вестерн-блот, изображающий продолжительность экспрессии полипептида GPR73 при использовании промотора phoA, индуцированного разведением в средах, лимитированных по фосфатам.
На Фигуре 15 представлен Вестерн-блот, изображающий продолжительность экспрессии полипептида GPR73 при использовании промотора phac, индуцированного разведением в средах, лимитированных по фосфатам, и добавлением IPTG.
На Фигуре 16 представлен Вестерн-блот, сравнивающий максимальную экспрессию GPR73 при использовании индуцированных промоторов phoA и phac.
На Фигуре 17 представлен Вестерн-блот, изображающий полипептид MS4A4A, экспрессированный вследствие частичного блокирования при использовании промотора phoA (средняя дорожка) и мутантного промотора, tphac (правая дорожка).
На Фигуре 18 представлен Вестерн-блот, изображающий продолжительность экспрессии полипептида MS4A4A при использовании промотора phoA, индуцированного разведением в средах, лимитированных по фосфатам.
На Фигуре 19 представлен Вестерн-блот, изображающий продолжительность экспрессии полипептида MS4A4A при использовании промотора tphac, индуцированного разведением в средах, лимитированных по фосфатам, и добавлением IPTG.
На Фигуре 20 представлен Вестерн-блот, сравнивающий максимальную экспрессию MS4A4A при использовании индуцированных промоторов phoA и tphac.
На Фигуре 21 схематически представлены конструкции для экспрессии мультитрансмембранных полипептидов. Показаны типичные составляющие экспрессирующих векторов.
На Фигуре 22 показана аминокислотная последовательность лидеров trpLE и sLE.
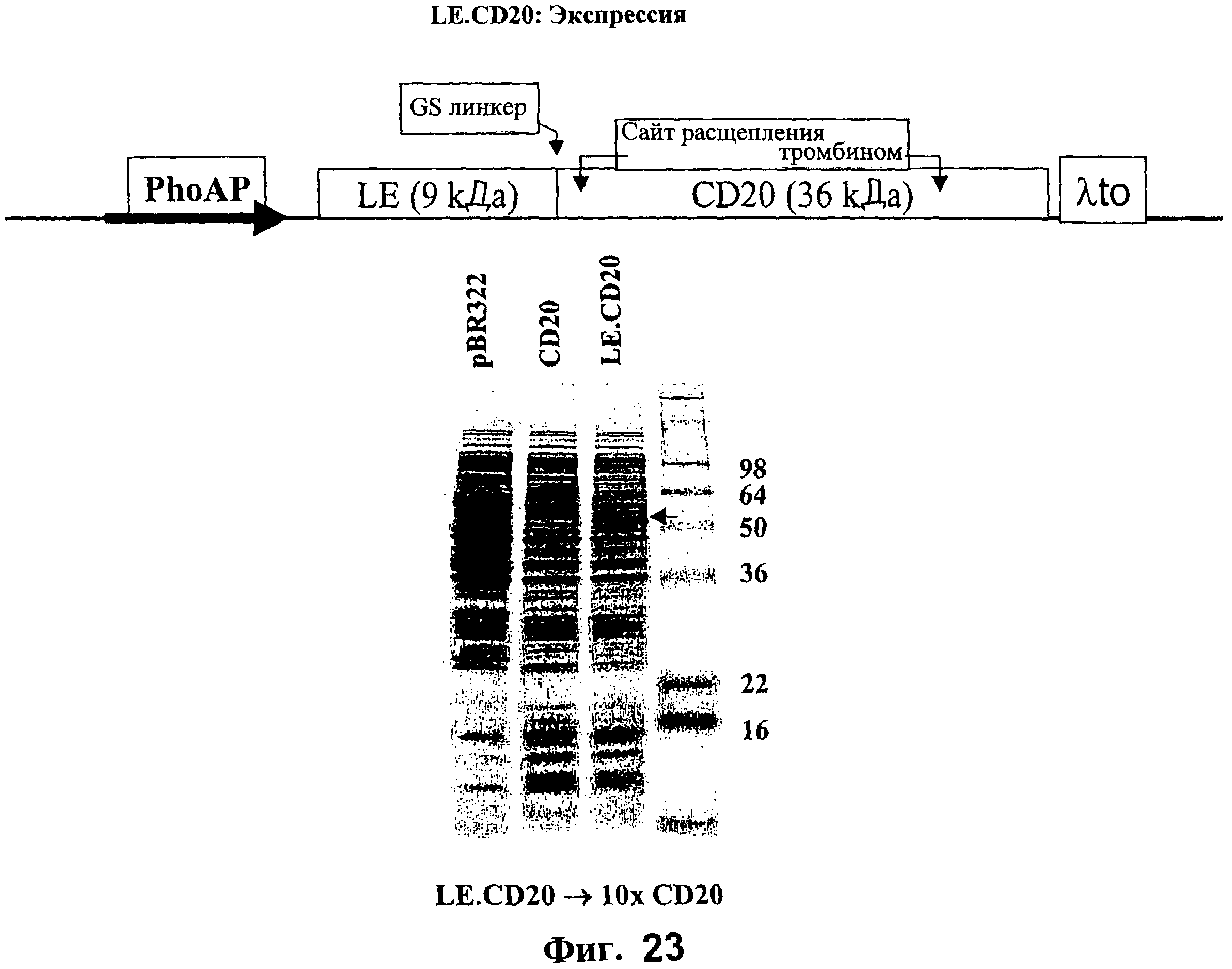
На Фигуре 23 схематически представлена диаграмма экспрессирующего вектора для экспрессии CD20 и окрашенный Кумасси синим гель, показывающий экспрессию и продуцирование CD20 и LE.CD20 в клетках Е.coli.
На Фигуре 24 показан Вестерн-блот и окрашенный Кумасси синим гель, демонстрирующий экстракцию LE.CD20, экспрессированного в клетках Е.coli.
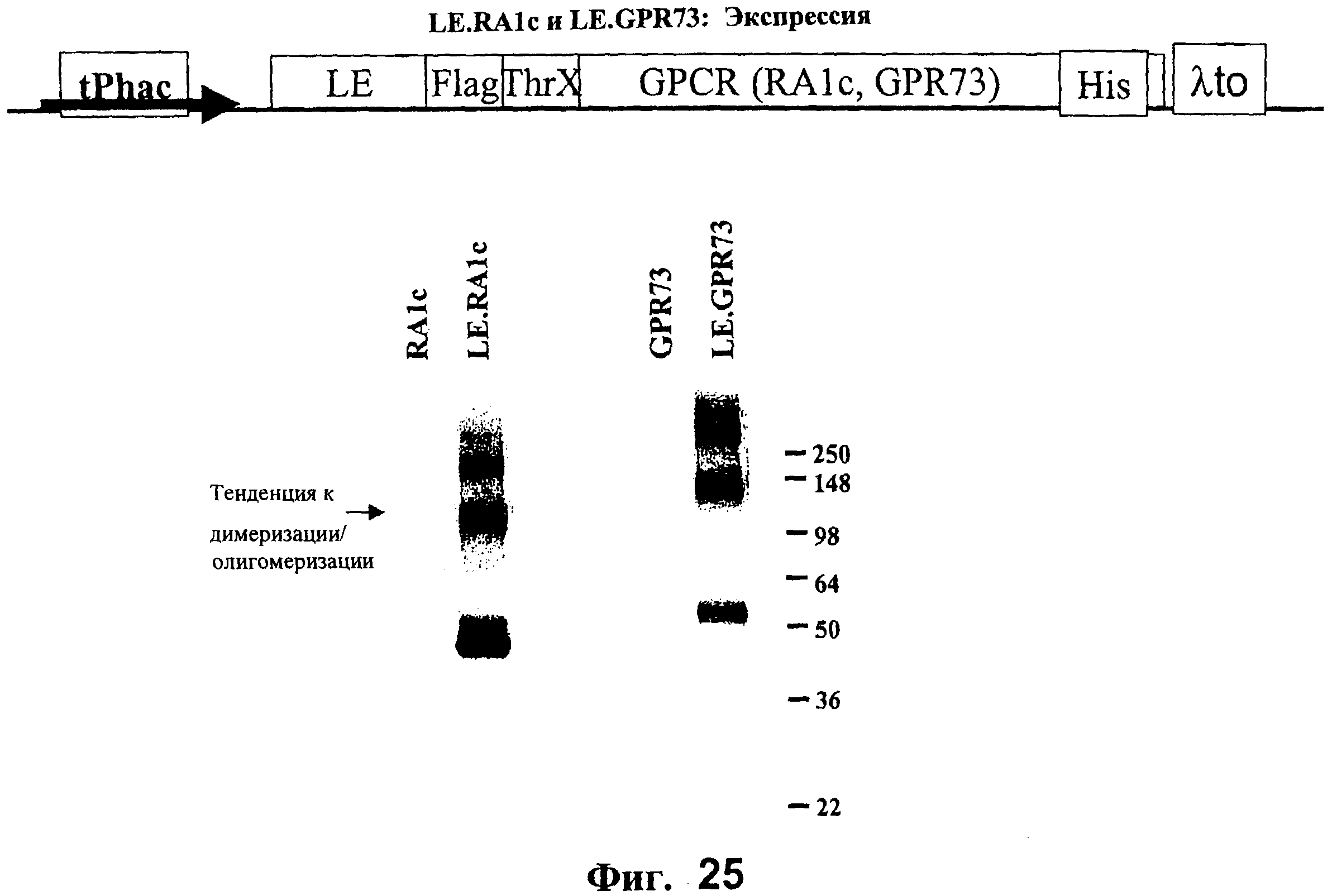
На Фигуре 25 схематически представлена диаграмма экспрессирующего вектора для экспрессии RA1c или GPR73 и Вестерн-блот, показывающий экспрессию LE.RA1c и LE.GPR73 по сравнению с контрольными белками.
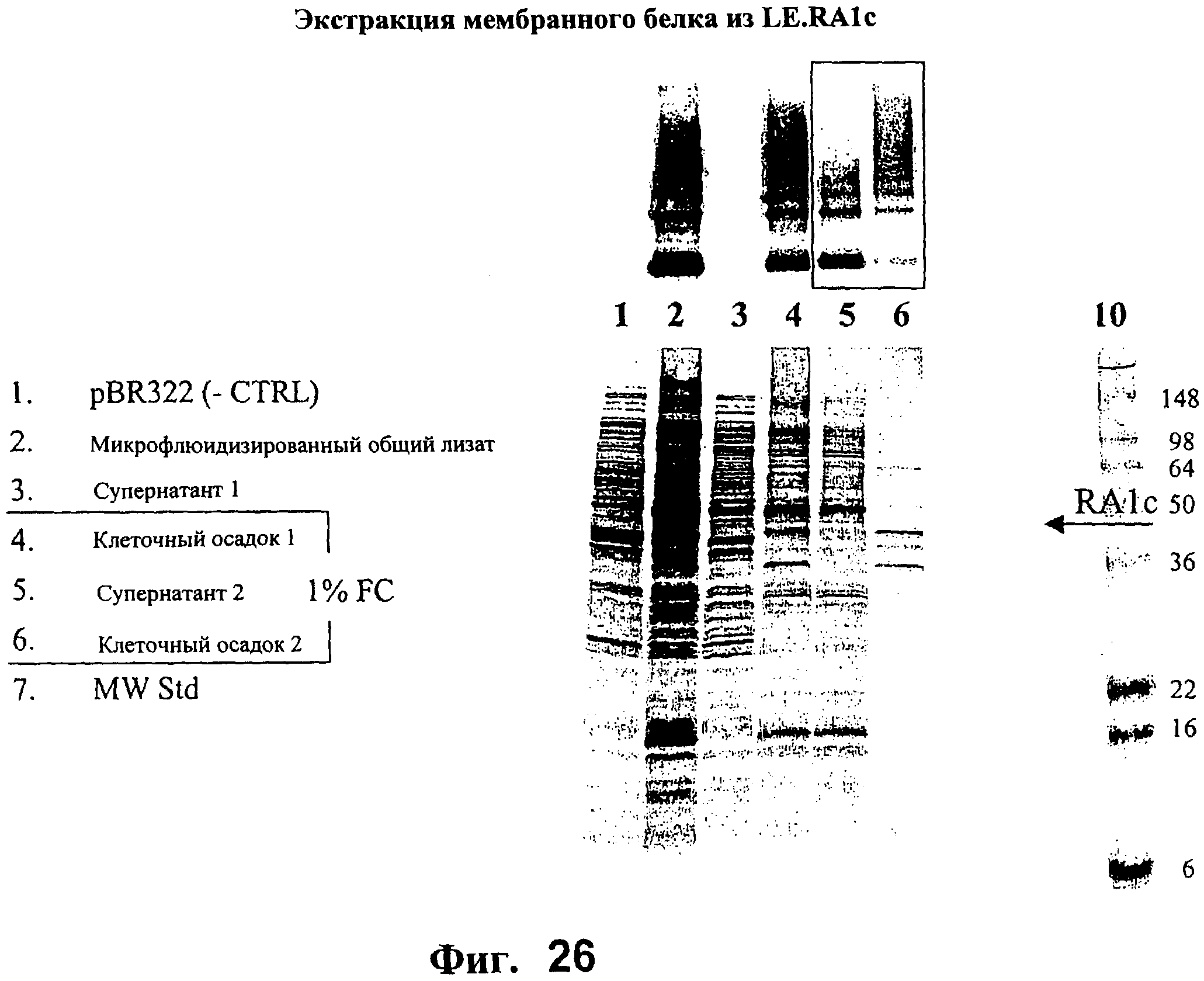
На Фигуре 26 представлен гель, окрашенный Кумасси синим, показывающий белок LE.RA1c, экспрессированный и экстрагированный из клеточных мембран Е.coli.
На Фигуре 27 представлен Вестерн-блот, показывающий LE.GPR73, экстрагированный из клеточных мембран Е.coli cell.
На Фигуре 28 представлен график, изображающий связывание специфичного к конформации CD20 антитела, ритуксимаба, с LE.CD20 и sLECD20, экспрессированными в Е.coli и экстрагированными, как описано в Примере 10. Для образцов sLE и LE метку LE удаляют расщеплением (гидролизом) тромбином, а образцы окисляют диализом. Незакрашенные кружки; hCD20, экспрессированный с LE меткой, незакрашенные треугольники; hCD20, экспрессированный с sLE меткой, закрашенные кружки; hCD20, экспрессированный с меткой HQ (на лидере LE), и х; PBS контроль.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
I. Определения
Антитело "со "зрелой" (повышенной) аффинностью" представляет собой антитело, содержащее одно или более изменений в одной или более гипервариабельных областей, которые повышают аффинность связывания антитела с целевым антигеном. Антитела со "зрелой" аффинностью могут иметь наномолярные или пикомолярные аффинности к целевому антигену. Антитела со "зрелой" аффинностью можно получать методами, известными в уровне техники, таким, например, как "перетасовка экзонов" (шаффлинг) VH и VL доменов (Marks et al., 1992, Bio/Technology, 10: 779-783), случайный мутагенез остатков CDR и/или каркасных остатков (Barbas et al., 1994, Proc. Nat. Acad. Sci. USA, 91: 3809-3813; Scier et al., 1995, Gene, 169: 147-155; Yelton et al., 1995, J. Immunol., 155: 1994-2004; Jackson et al., 1995, J. Immunol., 154: 3310-3319; и Hawkins et al., 1992, J. Mol. Biol., 226: 889-896), и метод фагового дисплея (Lowman et al., 1991, Biochemistry. 30: 10832-10838; Hawkins et al., 1992, J. Mol Biol., 226, 889-896; Патент США 6172213).
Термины "антитело" и "иммуноглобулин" применяются взаимозаменяемо в самом широком смысле и включают моноклональные антитела (полноразмерные или интактные моноклональные антитела), поликлональные антитела, гуманизированные, мультивалентные антитела, полиспецифические антитела при условии, что они проявляют нужную биологическую активность, и фрагменты антитела.
"Фрагменты антитела" содержат участок интактного антитела, обычно антигенсвязывающую или вариабельную область интактного антитела. Примеры фрагментов антитела включают, но без ограничения, Fab фрагменты, Fab′ фрагменты, Fd′ фрагмент, Fv фрагмент, Fd фрагмент, F(ab′)2 фрагмент, dAb фрагмент, антитела без шарнирного участка, одноцепочечные антитела, искусственно созданные мини-антитела (diabodies), "одноплечие" антиген-связывающие молекулы (содержащие легкую цепь, тяжелую цепь и усеченную по N-концу константную область тяжелой цепи, достаточную для образования Fc области, способной повышать период полужизни "одноплечей" антиген-связывающей молекулы) и линейные антитела.
Термин "моноклональное антитело" по данному описанию относится к антителу, полученному из популяции практически гомогенных антител, т.е. индивидуальные антитела, образующие популяцию, практически идентичны, за исключением вариантов, которые могут возникать в процессе продуцирования антитела.
Термин "моноклональные антитела" конкретно включает "химерные" антитела (иммуноглобулины), в которых участок тяжелой и/или легкой цепи идентичен или гомологичен соответствующим последовательностям в антителах из конкретных видов или принадлежащих к конкретному классу или субклассу антител, в то время как остальная часть цепи (цепей) идентична или гомологична соответствующим последовательностям в антителах из других видов или принадлежащим к другому классу или субклассу антител, а также фрагменты таких антител при условии, что они проявляют заданную биологическую активность (патент США 4816567; Morrison et al., 1984, Proc. Natl. Acad. Sci. USA, 81: 6851-6855).
Термин "биологический образец" относится к образцу из организма любого животного, такого как млекопитающее, например человек. Биологический образец можно получать, например, от больных сосудистыми заболеваниями, диабетом, раком. Биологический образец может представлять собой, например, биологические жидкости, такие как сыворотка, плазма, стекловидная жидкость, лимфатическая жидкость, синовиальная жидкость, фолликулярная жидкость, семенная жидкость, околоплодная жидкость, молоко, цельная кровь, моча, спинномозговая жидкость, слюна, мокрота, слезы, пот, слизь и тканевая культуральная среда, а также тканевые экстракты, такие как гомогенизированная ткань, клеточные экстракты, или цельные клетки или ткани. Например, биологический образец может представлять собой сыворотку, плазму или мочу.
"Буфер" по данному описанию относится к забуференному раствору, который устойчив к изменениям рН за счет действия своих компонентов: кислоты и сопряженного основания.
Термин "CD20 мутант" или "CD20 вариант" относится к CD20 полипептиду, который содержит аминокислотную последовательность, отличающуюся от эталонной CD20 аминокислотной последовательности, или кодируемую нуклеотидной последовательностью, отличающейся от эталонной CD20 нуклеотидной последовательности. CD20 мутант включает изменение аминокислотной последовательности, которое можно осуществить заменой, делепией или инсерцией одной или более аминокислот в эталонной последовательности.
Термин "реагент для "захвата"" (захватывающий реагент) относится к реагенту, способному связывать или захватывать целевую молекулу в образце. Комплекс-реагент для захвата - молекула-мишень можно отделять от остальной части образца при определенных условиях. Захватывающий реагент может быть иммобилизованным или иммобилизуемым. Например, в иммуноферментном сэндвич-анализе захватывающим реагентом может являться антитело или смесь различных антител к целевому антигену.
Термин "детергент" относится к агенту, который может содержать соли длинноцепных алифатических оснований или кислот, или гидрофобные частицы, такие как сахара, и который обладает как гидрофильными, так и гидрофобными свойствами. Обладая как гидрофильными, так и гидрофобными свойствами, детергент может вызывать особый эффект. Детергенты по данному описанию обладают способностью разрушать клеточные мембраны и солюбилизировать полипептиды.
Термин "детектируемое (обнаруживаемое) антитело" относится к антителу, которое можно обнаруживать с помощью метки, амплифицированной с помощью метода обнаружения, или опосредованно, например, с помощью другого, меченого антитела. Для прямого мечения антитело обычно конъюгируют с частицей, которую можно обнаружить каким-либо методом. Обычно антитело можно метить детектируемой меткой, включая, но без ограничения, флуоресцентную метку, радиоизотоп или ферментный субстрат ("ферментно-субстратная метка"). Метка может быть непрямо конъюгирована с антителом. Например, антитело может быть конъюгировано с биотином или любым из трех категорий меток, указанных выше, а любая из трех широких категорий указанных выше меток может быть конъюгирована с авидином, или наоборот. Биотин селективно связывается с авидином и таким образом метка может конъюгироваться с антителом таким непрямым способом. Или же, для осуществления непрямого конъюгирования метки с антителом антитело конъюгируют с малым гаптеном (например, дигоксином), а метка одного из различных указанных выше типов конъюгируется с антителом к гаптену (например, антителом против дигоксина).
Термин "детектирующий реагент" (реагент для обнаружения) относится к обнаружению (детектированию) метки и включает детектирующие агенты, которые амплифицируют иммобилизованную метку таким образом, что происходит захват метки на микротитрационном планшете. Способом обнаружения может включать, например, детектирующий агент, такой как авидин или стрептавидин, меченный флуоресцентной или хромофорной частицей.
Термин "метка экспрессии" относится к пептидной последовательности или метке, связанной с N- или С-концом зрелого полипептида или конъюгированной с конкретным остатков в зрелом полипептиде, которая предоставляет один способ идентификации и/или выделения экспрессированного полипептида. Метка экспрессии может кодироваться как компонент вектора или содержать часть встроенной в вектор экспрессии последовательности, кодирующей полипептид. Примеры меток экспрессии включают, но без ограничения, метки поли-His ("хвосты") (патент США 4569794), FLAG, myc, биотин, авидин и т.п. Такие метки общеизвестны и выпускаются промышленностью (см., например, Qiagen, Valencia, CA).
Термин "гетерологичный" относится к элементам, встречающимся там, где обычно они не обнаруживаются. Например, промотор может быть связан с гетерологичной нуклеотидной последовательностью, например с последовательностью, которая обычно не встречается как функционально связанная с промотором. В данном описании в применении к промоторному элементу гетерологичный означает промоторный элемент, который отличается от промоторного элемента, обычно связанного функционально с нативным промотором, либо последовательностью, либо видом, либо числом. Например, гетерологичный контрольный элемент в промоторной последовательности может являться контрольным/регуляторным элементом другого (отличного) промотора, добавленного для усиления контроля промотора, или дополнительным контрольным элементом того же самого промотора.
Выражение "индуцировать экспрессию" по данному описанию означает количественно повысить транскрипцию и/или трансляцию при использовании специфических генов экспозицией клеток, содержащих такие гены, с эффектором или индуктором, которыми могут быть реагент или условие.
"Индуктор" ("индуцирующий фактор") представляет собой химический или физический агент, который, в случае применения его к популяции клеток, количественно повышает транскрипцию при использовании специфических генов. Индукторы обычно представляют собой малые молекулы, чье действие является специфическим в отношении конкретных оперонов или групп генов, и могут включать сахара, фосфат, спирт, ионы металлов, гормоны, тепло, холод и т.п. Например, изопропил (бета)-D-тиогалактопиранозид (IPTG) и лактоза являются индукторами промотора tacII, a L-арабиноза является подходящим индуктором промотора арабинозы. Промотор гена pho, такой как phoA (Chang et al., 1987, Gene, 55: 189-196) и pho5, индуцируется при низкой концентрации фосфата в среде.
Реагент может "иммобилизоваться" на или в подложке за счет образования ковалентной связи между функциональной группой реагента и реактивной (реакционно-способной) группой на поверхности твердой фазы. В другом варианте изобретения реагент "иммобилизуется" на твердой фазе за счет адсорбции и ионного связывания и может улавливаться в твердой фазе, например, в клетках или латексных полимерах или микрокапсулах (см. Holenberg et al., в Enzymes as Drugs, John Wiley & Sons NY (1981), pages 396-411). Реагент практически сохраняет свою способность связываться с нужным полипептидом и/или модифицировать нужный полипептид, иммобилизованный на твердой фазе.
Термин "выделенный" (изолированный) при описании различных полипептидов, раскрываемых в данном описании, означает полипептид, который был идентифицирован и отделен от и/или регенерирован из его естественного окружения. Выделенный полипептид не ассоциирован, по меньшей мере, ни с одним компонентом, с которым он ассоциирован в естественном состоянии. Загрязняющие примеси из окружающей среды представляют собой материалы, которые обычно мешают при применении полипептида в диагностике или терапии и могут включать ферменты и другие белковые и небелковые растворы. Выделенный полипептид включает полипептид in situ в рекомбинантных клетках. Однако обычно выделенный полипептид получают, по меньшей мере, одной стадией очистки.
"Выделенный CD20" по данному описанию относится к белку CD20, не содержащему клетки или мембраны, и может быть, например, в растворимом виде в растворе детергента.
"Выделенная" молекула нуклеиновой кислоты или "выделенный" полинуклеотид представляет собой нуклеотидную молекулу, которая идентифицирована и отделена, по меньшей мере, от одной загрязняющей нуклеотидной молекулы, которая обычно ассоциирована с природным источником. Выделенная нуклеиновая кислота может, например, не ассоциироваться ни с какими компонентами, с которыми она ассоциирована в естественном окружении. Выделенная нуклеотидная молекула имеет либо другую форму, либо другие параметры, чем те, которые она имеет в природе.
"IPTG" представляет собой соединение "изопропил (бета)-D-тиогалактопиранозид" и применяется в данном описании в качестве индуктора оперона lac. IPTG связывается с lac-репрессором, вызывая конформационные переходы в lac-репрессоре, которые приводят к диссоциации и отделению lac-репрессора от lac-оператора. Будучи несвязанным с lac-оператором, функционально связанный промотор активируется и транскрибируются гены "даунстрим".
Термин "lac-оператор" относится к нуклеотидной последовательности, которая может быть связана lac-репрессором, lacI, как описано, например, в Jacob et al., 1961, J. Mol. Biol., 3: 318-356. Промотор не активируется, когда lac-репрессор связан с lac-оператором. Когда lac-репрессор индуцируется и диссоциирует от оператора, промотор активируется.
Термин "лидерная последовательность" относится к нуклеотидной последовательности, расположенной "апстрим" (выше) нужной кодирующей последовательности. Лидерные последовательности по данному описанию содержат специфические последовательности, известные как эффективно связывающиеся с рибосомами, тем самым сообщающие большую эффективность инициации трансляции некоторых полинуклеотидов. Как указывается в данном описании, лидерная последовательность содержит последовательность инициации трансляции и спейсерную последовательность, чтобы способствовать элонгации трансляции по данному описанию.
Термин "среда с низким содержанием фосфата" ("низкофосфатная среда") или "среда с ограниченным содержанием фосфата" ("фосфат-лимитирующая среда") по данному описанию относится к средам, содержащим низкую концентрацию фосфата в растворе. Например, промотор phoA включается, при концентрация фосфата в среде от нескольких капель, примерно, до 4 мкМ (микромолей) или менее. Однако создают среды с ограниченным содержанием фосфата, содержащие более 4 мкМ (микромолей) фосфата, которые дают клеткам возможность расти до включения промотора. Примеры сред с ограниченным содержанием фосфата включают, но без ограничения, C.R.A.P. среды, описанные в Simmons et al., 2002, J. Immunol. Methods, 263: 133-147 (имеющие начальную концентрацию фосфата, примерно, 1.9 М за счет следовых примесей из дрожжевого экстракта и из других источников), и среды, описанные в Chang et al., 1987, Gene 55: 189-196.
Термин "млекопитающее" по данному описанию относится к любому животному классифицированному как млекопитающее, включая человека, домашних и сельскохозяйственных животных и животных в зоопарке, спортивных животных или комнатных животных, таких как собаки, лошади, кошки, коровы и т.д.
Термин "MS4A полипептид"относится к полипептиду, кодированному генами подсемейства А (MS4A) семейства трансмембранного белка с 4 доменами. См., например, Ishibashi et al., 2001, Gene, 264: 87-93. Полипептид MS4A может быть природным полипептидом MS4A или вариантом природного полипептида MS4A. Члены семейства MS4A гена представляют собой структурно сходные полипептиды. Каждый пронизывают клеточную мембрану четыре раза, как N-конец, так и С-конец локализован на стороне цитоплазмы клеточной мембраны, и оба содержат внеклеточную петлю протяженностью, примерно, 50 аминокислот. Полипептиды MS4A включают CD20, β цепь высокоаффинного рецептора IgE, HTm4, MS4A4A, MS4A7, и т.п. Термин также включает варианты и изоформы полипептидов, кодирующихся генами MS4A genes. Это семейство генов является консервативным у млекопитающих, и "MS4A полипептид" включает человеческий, мышиный, крысиный и т.п. полипептиды полипептида MS4A.
"Вариант" MS4A полипептида относится к MS4A полипептиду, который содержит аминокислотную последовательность, отличающуюся от эталонной последовательности, или кодируется нуклеотидной последовательностью, отличающейся от эталонной последовательности. Эталонная (стандартная) последовательность может представлять собой полноразмерную нативную MS4A полипептидную последовательность, внеклеточный домен MS4A полипептида, или любой другой фрагмент полноразмерной последовательности MS4A полипептида. В некоторых вариантах изобретения эталонная последовательность представляет собой нуклеотидную последовательность или аминокислотную последовательность природного CD20, например, такую как SEQ ID NO: 1 (аминокислотная последовательность) или SEQ ID NO: 2 (нуклеотидная последовательность). Аминокислотная последовательность варианта MS4A полипептида обычно, по меньшей мере, на 80% идентична эталонной последовательности.
Варианты MS4A полипептида включают "природные" варианты, в том числе аллельные варианты, а также варианты, полученные изменением одного или более нуклеотидов или одной или более аминокислот. Вариантный полипептид можно получать модификацией нуклеотидной последовательности или аминокислотной последовательности MS4A полипептида. Например, вариант можно получать добавлением, заменой и/или делецией нуклеотидов или аминокислот. Вариантный MS4A полипептид, применимый в способах по изобретению, может иметь идентичность последовательностей, например, по меньшей мере, 80%, по меньшей мере, около 85%, по меньшей мере, около 90%, по меньшей мере, около 91%, по меньшей мере, около 92%, по меньшей мере, около 93%, по меньшей мере, около 94%, по меньшей мере, около 95%, по меньшей мере, около 96%, по меньшей мере, около 97%, по меньшей мере, около 98%, по меньшей мере, около 99% с эталонной последовательностью MS4A, например с эталонной последовательностью человеческого CD20, такой как SEQ ID NO: 1.
Термин "трансмембранный белок" или "пронизывающий мембрану белок" относится к полипептиду, содержащему один или более сегментов, "внедренный" в фосфолипидный бислой клеточной мембраны. Помимо этого, трансмембранный белок может содержать внутриклеточный домен, внеклеточный домен или и тот и другой. Клеточная мембрана может являться мембраной бактериальных, дрожжевых клеток, клеток млекопитающих и т.п.
Термин "мембраносвязывающий домен" или "трансмембранный домен" относится к участку трансмембранного белка, который внедрен в фосфолипидный бислой клеточной мембраны.
Термин "нативная конформация" относится к трехмерной конфигурации полипептида в естественном состоянии. Нативная конформация может относиться к третичной или четвертичной структуре полипептидов. "Нативная конформация" солюбилизированного трансмембранного полипептида по данному описанию достаточна для того, чтобы солюбилизированный полипептид мог применяться в качестве иммуногена для продуцирования антител, распознающих трансмембранный полипептид в клетке или в качестве связывающего лиганда для связывания антител, распознающих трансмембранный белок в клетке.
Термин "неионный" относится к молекуле, которая не ионизируется в растворе, т.е. является инертной в "ионном" смысле.
Нуклеотидная последовательность (последовательность нуклеиновых кислот) или полинуклеотид является "функциональной связанной (или связанным)", когда она (он) функционально связана (связан) с другой нуклеотидной последовательностью. Например, ДНК препоследовательности или секреторного лидера функционально связана с ДНК полипептида, если он экспрессируется как пребелок, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или сайт связывания рибосом функционально связан с кодирующей последовательностью, если он позиционирован таким образом, чтобы содействовать трансляции. Как правило, "функционально связанный" означает, что связанные последовательности ДНК являются смежными (непрерывными) и, в случае секреторного лидера, смежными (непрерывными) в рамке считывания. Связывание можно осуществлять дотированием в подходящих сайтах рестрикции. Если такие сайты отсутствуют, в соответствии с обычной практикой используют синтетические олигонуклеотидные адаптеры или линкеры.
"Плазмиды" обозначаются строчной буквой "р" ("п"), за которой следуют заглавные буквы и/или числа. Исходные плазмиды в данном описании являются, либо продажными, либо, без ограничения, общедоступными, либо их можно создать с помощью доступных плазмид по опубликованным методикам. Кроме того, плазмиды, эквивалентные описанным плазмидам, известны в уровне техники и очевидны для рядового специалиста в данной области техники.
"Процент (%) идентичности аминокислотных последовательностей" в отношении полипептидов, идентифицированных в данном описании, определяется как процентное содержание аминокислотных остатков в кандидатной последовательности, идентичных аминокислотным остаткам в эталонной последовательности после выравнивания последовательностей и, при необходимости, введения гэпов для достижения максимальной идентичности последовательностей, и при этом никакие консервативные замены не рассматриваются как часть идентичности последовательностей. Добиться выравнивания с целью определения идентичности аминокислотных последовательностей в процентах можно различными методами, известными специалистам в данной области техники, например, используя доступные компьютерные программы, такие как BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области техники могут определить подходящие характеристики для определения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Программа ALIGN-2 доступна через Genentech, Inc., South San Francisco, California.
Для целей по данному описанию % идентичности аминокислотных последовательностей данной аминокислотной последовательности А с данной аминокислотной последовательностью, относительно или против данной аминокислотной последовательности В (что иначе можно выразить как: данная аминокислотная последовательность А, которая имеет или содержит определенный % идентичности аминокислотной последовательности с данной аминокислотной последовательностью, относительно или против данной аминокислотной последовательности В, или как: данная аминокислотная последовательность А на определенный % идентична данной аминокислотной последовательности В) рассчитывается следующим образом:
100, умноженное на дробь X/Y,
где Х означает число аминокислотных остатков, оцениваемых как тождественные с помощью программы выравнивания последовательностей по этой программе выравнивания А и В, и где Y обозначает общее число аминокислотных остатков в В. Разумеется, если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, % идентичности аминокислотной последовательности А аминокислотной последовательности В не будет равен % идентичности аминокислотной последовательности В аминокислотной последовательности А.
"Процент (%) идентичности нуклеотидных последовательностей" определяется как процентное содержание нуклеотидов в кандидатной последовательности, идентичных нуклеотидным остаткам в эталонной кодирующей полипептид нуклеотидной последовательности после выравнивания последовательностей и, при необходимости, введения гэпов для достижения максимальной идентичности последовательностей. Добиться выравнивания с целью определения идентичности нуклеотидных последовательностей в процентах можно различными методами, известными специалистам в данной области техники, например, используя доступные компьютерные программы, такие как BLAST, BLAST-2, ALIGN, ALIGN-2 или Megalign (DNASTAR). Специалисты в данной области техники могут определить подходящие характеристики для определения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей.
Для целей по данному описанию % идентичности нуклеотидных последовательностей данной нуклеотидной последовательности С с данной нуклеотидной последовательностью, относительно или против данной нуклеотидной последовательности D (что иначе можно выразить как: данная нуклеотидная последовательность С, которая имеет или содержит определенный % идентичности нуклеотидной последовательности с данной нуклеотидной последовательностью, относительно или против данной нуклеотидной последовательности D, или как: данная нуклеотидная последовательность С на определенный % идентична данной нуклеотидной последовательности D) рассчитывается следующим образом:
100 умноженное на дробь W/Z,
где W означает число нуклеотидов, оцениваемых как тождественные с помощью программы выравнивания последовательностей по этой программе выравнивания С и D, и где Z обозначает общее число нуклеотидных остатков в D. Разумеется, если длина нуклеотидной последовательности С не равна длине нуклеотидной последовательности D, % идентичности нуклеотидной последовательности С нуклеотидной последовательности D не будет равен % идентичности нуклеотидной последовательности D нуклеотидной последовательности С.
Термин "phoA промотор" относится к промотору структурного гена щелочной фосфатазы, phoA. Различные бактерии, в частности, Enterobacteriaceae, имеют phoA ген и phoA промотор. Е.coli phoA промотор приведен в данном описании в качестве примера, он имеет последовательность SEQ ID NO: 5.
Термин "мутантный промотор" или "вариантный промотор" относится к промотору, имеющему нуклеотидную последовательность, отличающуюся от эталонной последовательности. Например, мутантные phac и tphac промоторы отличаются от эталонного промотора phoA, как показано в Таблице 5. Изменение нуклеотидной последовательности промотора может быть вызвано заменой, делецией или инсерцией одной или более нуклеиновых кислот.
Термин "phoA" относится к гену, кодирующему металлофермент щелочную фосфатазу. В Е.coli фермент phoA является частью фосфат-регулона, в котором экспрессия phoA активируется более чем в 100 раз при истощении неорганического фосфата (см., например, Kriakov et al., 2003, J.Bacteriol., 185: 4983-4991). Бактериальные виды, отличные от Е.coli, содержат phoA гомологи (например, Klebsiella spp. Shigella spp. Mycobacterium smegmatis).
Термин "поли-His" обычно относится к аминокислотным остаткам, содержащим кратное число гистидиновых остатков, обычно 6-10 гистидиновых остатков. Кратное число (несколько) гистидиновых остатков часто используют в качестве метки экспрессии, называемой "поли-His метка ("хвост")" (см. патент США 4569794). Поли-his метки можно использовать для обнаружения и/или очистки полипептидов, например, нанося образец на аффинную колонку, такую как никелевая колонка.
Термин "полипептид" по данному описанию обычно относится к пептидам и белкам, содержащим более десяти аминокислот. Полипептиды могут быть "экзогенными", это означает, что они "гетерологичны", т.е. чужеродны для используемой клетки-хозяина, например человеческий полипептид, продуцированный бактериальной клеткой.
"Примат" обозначает любое из разряда млекопитающих, представляющее собой человека, человекообразных обезьян, обезьян и родственные виды, такие как лемуры и долгопяты.
"Очистка" означает повышение степени чистоты, например трансмембранного полипептида в композиции, путем удаления (полного или частичного), по меньшей мере, одной примеси из композиции. "Стадия очистки" может являться частью общего процесса очистки, дающего "практически чистую композицию". Практически чистая композиция содержит, по меньшей мере, около 90% весовых от общего веса композиции нужного полипептида и может содержать около 95% весовых.
Термин "редкие кодоны" или "минорные тРНК" относится к специфическим кодонам или тРНК с низким содержанием в клетках конкретного типа. См., например, Dong et al., 1996, J. Mol. Biol., 260: 649-663, где описывается относительное содержание тРНК и применимость кодонов в клетках Е.coli.
Термин "регуляторный элемент" 'или "контрольный элемент" относится к последовательностям ДНК, контролирующим инициацию транскрипции. Примеры контрольных или регуляторных элементов включают, но без ограничения, ТАТА бокс, операторы, энхансеры и т.п. Регуляторные или контрольные элементы включают элементы негативного контроля и элементы позитивного контроля. Элемент негативного контроля представляет собой элемент, который удаляется для активации транскрипции. Известно множество таких элементов негативного контроля, например системы оператор/репрессор. Например, связывание IPTG с lac репрессором диссоциирует (разрывается) от lac оператора, активируя и способствуя транскрипции. Другие негативные элементы включают Е.coli trp и лямбда системы. Элементом позитивного контроля является элемент, который добавляется для активации транскрипции. Известно множество таких элементов позитивного контроля, включая Е.coli pho бокс и варианты pho бокса, которые связывают phoB, сайт связывания ДНК Ма1Т, сайт связывания ДНК AraC и т.п. Например, связывание phoB промотора с pho боксом phoA промотора индуцирует активацию промотора.
Промоторы, содержащие в естественном состоянии как позитивные, так и негативные регуляторные элементы, встречаются редко. metE промотор является одним из примеров. См., например, Neidhardt, Ed., 1996, Escherichia coli and Salmonella, Second Ed., pages 1300-1309. Описания известных элементов позитивного и негативного контроля можно найти, например, в этом ссылочном материале. В некоторых вариантах изобретения промотор содержит как позитивный, так и негативный контрольный элемент, которые обеспечивают прямой контроль базальной (основной) экспрессии. Известно расположение позитивного или негативного контрольных элементов в промоторе или рядом с промотором для осуществления дополнительной регуляции промотора, оно описано, например, в Escherichia coli and Salmonella (Supra) на страницах 1232-1245.
Термины "ритуксимаб" или "ритуксан®" по данному описанию относятся к созданному методами генетической инженерии химерному мышиному/человеческому моноклональному антителу к CD20 антигену и обозначенному "С2В8" in US Patent No. 5,736,137 в патенте США 5736137, специально вводимому в данное описание в качестве ссылки. Антитело представляет собой иммуноглобулин IgG1 каппа, содержащий мышиные последовательности вариабельной области легкой и тяжелой цепей и человеческие последовательности константной области. Аффинность связывания ритуксимаба с CD20 антигеном составляет, примерно, 8.0 нМ.
Термин "солюбилизация" относится к растворению молекулы в растворе. В одном варианте изобретения рекомбинантный трансмембранный полипептид, экспрессируемый в бактериальном хозяине, представляет собой неионный или цвиттерионный детергент.
Термин "спейсерная последовательность" относится к полинуклеотидной последовательности, кодирующей аминокислоты, расположенные между последовательностью инициации трансляции и первым трансмембранным доменом.
Термин "строго контролируемый промотор" или "строго регулируемый промотор" относится к промотору, проявляющему малую базальную (основную) экспрессию или не проявляющему базальной (основной) экспрессии функционально связанных генов. Строго контролируемый или регулируемый промотор активирует экспрессию в специфически определенных контролируемых условиях.
Термин "терминатор транскрипции" относится к нуклеотидной последовательности, которая дает сигнал РНК полимеразе завершить транскрипцию. Терминаторы транскрипции общеизвестны и включают, например, но без ограничения, терминатор транскрипции лямбда λ0 (tao zero) (SEQ ID NO: 17), терминаторы транскрипции Е.coli rrnB1 Т1 и rrnB2 Т2 и сильный терминатор His оперона.
Выражение "энхансерная последовательность инициации трансляции" или "последовательность инициации трансляции" (TIS) по данному описанию относится к нуклеотидной последовательности, которая может определять сайт и эффективность инициации трансляции гена (см., например, McCarthy et al., 1990, Trends in Genetics, 6: 78-85). "Последовательность инициации трансляции" может также называться как область инициации трансляции (TIR).
Термин "цвиттер-ионный" или "диполярный" относится к молекулам, содержащим заряженные группы с противоположной полярностью.
II. Способы осуществления изобретения
А. Трансмембранные полипептиды
Трансмембранные полипептиды, такие как CD20, MS4A4A, RA1c, GPR73 и т.п., представляют собой потенциальные мишени для лекарственных средств при лечении заболеваний или нарушений, таких как рак. CD20 является целью (мишенью) для химерного антитела ритуксимаба (ритуксана®), ведущего лекарственного препарата для лечения неходжкинской лимфомы. Ритуксимаб распознает CD20 в нативной конформации, экспрессируемый на В клетках. Связывание ритуксимаба зависит от структуры петли между третьей и четвертой трансмембранной спиральной областью CD20, которая содержит цистеиновые остатки в положениях 167 и 183 (см. Фигуру 1).
Значительным препятствием при создании терапевтических препаратов, нацеленных на трансмембранные полипептиды, например CD20, является невозможность получать полезные выделенные и очищенные рекомбинантные или природные трансмембранные полипептиды. Например, для применения в качестве иммуногена или связывающего антигена выделенный или очищенный полипептид должен иметь адекватную "нативную" конформацию, чтобы распознаваться партнером по связыванию. Полипептид сохраняет достаточную нативную конформацию для распознавания лигандом, связывание которого зависит от структурных особенностей нативной конформации полипептида. Настоящее изобретение предусматривает векторы, содержащие новые промоторы, для продуцирования трансмембранных полипептидов в бактериальном хозяине, способы продуцирования трансмембранных полипептидов в бактериальном хозяине и способы выделения трансмембранных полипептидов из бактериального хозяина. Способы по изобретению дают трансмембранные полипептиды с высокими выходами и с достаточной "нативной" конформацией, чтобы применят их, например, в качестве иммуногенов и связывающих антигенов.
Трансмембранные белки содержат один или более трансмембранный домен и могут содержать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или 24 или более таких доменов. В одном варианте изобретения трансмембранный полипептид имеет, по меньшей мере, четыре трансмембранных домена. В другом варианте изобретения трансмембранный полипептид содержит 7 трансмембранных доменов, например, EG-VEGF рецептор, GPR73 и RA1c рецептор. Трансмембранные полипептиды, имеющие четыре трансмембранных домена, включают, например, членов MS4 семейства полипептидов. В другом варианте изобретения трансмембранный полипептид представляет собой CD20 полипептид или его вариант.
В нижеприведенном описании CD20 используется в качестве примера трансмембранного белка, применимого по изобретению. В методах экспрессии и солюбилизации по настоящему описанию применимы также дополнительные полипептиды, включая полипептиды, раскрываемые ниже, в разделе Примеры, так же как другие, нераскрываемые трансмембранные полипептиды.
1. CD20
CD20 представляет собой фосфопротеин протяженностью, примерно, 35 кДа, обнаруживаемый на поверхности более 90% В клеток периферической крови или лимфоидных органов. В литературе встречаются другие названия для CD20 "ограниченный антиген В-лимфоцитов" и "Вр35". CD20 описан, например, в Сlark et al., 1985, Proc. Natl. Acad. Sci. USA, 82: 1766-1770. CD20 экспрессируется в процессе раннего развития пре-В клетки и остается до дифференцировки плазматических клеток. Активация В клеток вызывает дополнительное повышение экспрессии CD20 (Valentine et al., 1987, Proc. Natl. Acad. Sci. USA, 84; 8085-8097). CD20 не экспрессируется в плазматических клетках. CD20 присутствует на нормальных, а также на злокачественных В клетках.
Настоящее изобретение включает выделенный CD20 млекопитающих, не содержащий клеток и клеточных мембран и сохраняющий нативную конформацию, достаточную для того, чтобы связывать ритуксимаб или его антиген-связывающий фрагмент. Примеры CD20 млекопитающих включают, без ограничения, человеческий CD20 и мышиный CD20, изображенные ниже, в Таблице 2 как SEQ ID NO: 1 и 3. Эталонные нуклеотидные последовательности, кодирующие человеческий CD20 (NCBI инвентарный номер ВС002807) и мышиный CD20 (NCBI инвентарный номер NM_007641), находятся в базе данных NCBI, www.ncbi.gov. Человеческий CD20 существует, например, в различной степени фосфорилирования в В клетках, но варианты сплайсинга отсутствуют.
Как графически показано на Фигуре 1, CD20 представляет собой четырежды пронизывающий мембрану полипептид, имеющий оба конца на стороне цитоплазмы клеточной мембраны. Первая внеклеточная петля (петля А) образуется между первым и вторым трансмембранным доменом, а вторая внеклеточная петля (петля В) образуется между третьим и четвертым трансмембранными доменами. Петля В больше, чем петля А. Петля А не слишком выступает из трансмембранных доменов. Петля В имеет протяженность 46 аминокислот и значительно выступает из трансмембранных доменов. Петля В имеет протяженность от Asn140, примерно, до Ser185 и содержит дисульфидную связь между Cys167 и Cysl83. Связывание CD20 ритуксимабом зависит от петли В. См., например, Polyak and Deans, 2002, Blood 99: 3256-3262.
CD20 полипептиды по изобретению растворимы в неионном или в цвиттер-ионном детергенте и сохраняют достаточно "нативную" структуру петли В в детергенте, так что ритуксимаб или антиген-связывающий фрагмент ритуксимаба может связывать полипептид. В одном варианте изобретения петля, образующаяся между третьим и четвертым трансмембранными доменами, сохраняется в выделенном CD20. Петля содержит дисульфидную связь между Cys167 и Cys183 и включает, например, остатки CD20 от I164 до Y184, как показано на Фигуре 1. Петля может содержать, например, около 40-60 аминокислот и может быть около 40-около 50 аминокислот длиной, около 45-около 50 аминокислот длиной, или около 46 аминокислот длиной. В одном варианте изобретения петля имеет протяженность от Asn140 до Ser185 и содержит дисульфидную связь между Cys167 и Cys183. В одном варианте изобретения петля может быть связана ритуксимабом или его антигенсвязывающим фрагментом.
2. Вариантные трансмембранные полипептиды
Настоящее изобретение также включает варианты трансмембранных полипептидов, таких как CD20, которые могут быть природными или рекомбинантными. Варианты включают, например, делеции, инсерции или замены аминокислотных остатков в человеческой эталонной последовательности.
Вариантный трансмембранный полипептид содержит аминокислотную последовательность, по меньшей мере, примерно, на 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более идентичную эталонной последовательности млекопитающего. Например, эталонная последовательность может представлять собой последовательность мышиного или человеческого CD20. В одном варианте изобретения эталонная последовательность CD20 представляет собой SEQ ID NO: 1. В другом варианте изобретения эталонная последовательность CD20 представляет собой SEQ ID NO: 3.
Фрагмент CD20, который включает третий и четвертый трансмембранный домены и петлю между ними (петлю В), может представлять собой эталонную последовательность, например, включающую остатки от К116 до N214 включительно. Эталонная последовательность CD20 включает, например, остатки от I164 до Y184 включительно петли В. Фрагмент CD20 может включать, например, остатки от Х до Y последовательности SEQ ID NO: 1, где Х представляет собой любой остаток последовательности от Т104 до I128, соответствующей последовательности человеческого CD20, показанной на Фигуре 1, а Y обозначает любой остаток последовательности от V196 до Р297, содержащей дисульфидную связь между остатками С167 и С183 в невосстанавливающих условиях (при отсутствии восстановления). Например, фрагмент CD20 может включать остатки от N140 до S185, показанные на Фигуре 1, с дисульфидной связью между остатками С167 и С183 в невосстанавливающих условиях.
Трансмембранные варианты, например варианты CD20, являются растворимыми в неионном или цвиттер-ионном детергенте, таком как DDPC, и сохраняют достаточную "нативную" петлевую структуру в детергенте для связывания известного детектирующего антитела, такого как ритуксимаб (например, ритуксан®) или его антиген-связывающий фрагмент. В одном варианте изобретения трансмембранные варианты включают петлю во внеклеточном домене, например в CD20, петлю, образующуюся между третьим и четвертым трансмембранным доменом. Петля может иметь, например, около 30-100 аминокислот в длину, около 40-60 аминокислот в длину, около 40-50 аминокислот в длину, около 45-50 аминокислот в длину или около 46 аминокислот в длину. В одном варианте изобретения петля содержит дисульфидную связь, например, в случае CD20, дисульфидную связь при остатках, соответствующих Cys167 и Cys183 в SEQ ID NO: 1.
Трансмембранный вариантный полипептид может включать аминокислотные замены, например, в полноразмерном CD20 или в усеченных мутантах, обсуждавшихся выше, которые повышают экспрессию полипептида. Например, замена одного или более Cys111 и Cys220 в CD20 повышает экспрессию человеческого CD20 (см. Пример 1). Так, подходящий CD20 вариант содержит аминокислотную последовательность с заменой одного или более цистеиновых остатков, соответствующих Cys111 и Cys220 человеческого CD20 с аминокислотной последовательностью SEQ ID NO: 1. Можно осуществить неконсервативную замену цистеинового остатка, чтобы предотвратить возможное образование дисульфидной связи. В приведенных ниже Примерах Cys заменяют на Ser. Мутант C2S-CD20 (также называемый "C2S") (SEQ ID NO: 6) (см. таблицу 2) CD20 содержит двойную замену Cys111Ser и Cys220Ser.
CD20 варианты можно получать любыми известными методами введения замен, делеций или инсерций одной или более аминокислот, например, с помощью мутаций в нуклеотидных последовательностях, кодирующих CD20. В одном варианте изобретения "нативная" структура CD20 достаточна для сохранения связывания антитела с петлей В. Аминокислотные инсерции включают амино- и/или карбоксильных концевые слияния протяженностью от одного остатка до полипептида, содержащего сто или более остатков, а также инсерции внутрь последовательности из одного или множества аминокислотных остатков. Инсерции внутри последовательности (например, инсерции внутри последовательности CD20) могут содержать, например, около 1-10 остатков, 1-5 остатков или около 1-3 остатков. Делеции аминокислотной последовательности могут содержать, например, от 1 до 30 остатков, или от 1 до 10 остатков, и обычно являются непрерывными.
Указание для определения аминокислотного остатка (аминокислотных остатков), который (которые) нужно вставить, заменить или удалить, не оказывая вредного влияния на связывание антитела или фрагмента антитела, такого как ритуксимаб, с CD20, можно найти путем сравнения последовательности трансмембранного полипептида, например CD20, с последовательностью известных гомологичных белковых молекул, например, имеющих аналогичную структуру и/или аналогичные функциональные домены, и минимизации числа изменений в аминокислотной последовательности в областях высокой гомологии, например, выше 50%, 55% или 60% идентичности аминокислот. Например, идентичность аминокислотных последовательностей человеческого и мышиного CD20 составляет 72% при степени идентичности во внеклеточной петле между трансмембранными доменами три и четыре (петля В) 63%. Эталонные последовательности, такие как последовательность человеческого CD20 (SEQ ID NO: 1) и мышиного CD20 (SEQ ID NO: 3), используются для выравнивания и сравнения идентичности областей с высокой и низкой гомологией.
Функциональные домены можно также идентифицировать в полипептидах, о которых известно, что они имеют гомологию с трансмембранным полипептидом, таким как CD20. Последовательности функциональных доменов можно сравнивать и выравнивать с другими известными последовательностями, например полипептидами CD20 или семейства MS4A. CD20, β цепь рецептора Ig и HTm4 имеют общую структуру, четырежды пронизывающую мембрану, с N- и С-концевыми доменами. Внеклеточная петля, примерно из 50 аминокислот, представляет собой другой общий мотив семейства гена MS4A. Кроме того, эта структура является общей для различных видов, например человеческого и мышиного. Эти три гена локализованы 11q12-q13.1 у человека и на хромосоме 19 у мыши (Adra et al., 1989, Proc. Natl. Acad. Sci. USA, 91: 10178-10182; Hupp et al., 1989, J.Immunol., 143: 3787- 3791; Tedder et al., 1988, J.Immunol., 141:4388-4394; Tedder et al., 1989, J.Immunol., 142:2555-2559). Полагают, что три гена происходят из общего предшественника (Liang et al., 2001, Supra).
Кандидатные (возможные) положения аминокислотной замены идентифицируют как такие положения, которые обнаруживают высокую степень вариабельности в аминокислотах, т.е., по меньшей мере, 3 различных аминокислоты обнаруживают в этом положении при выравнивании и сравнении, или имеют более низкий процент идентичности последовательностей, т.е. идентичность последовательностей ниже 90%. Когда последовательности выравниваются, положения, проявляющие вариабельность, могут иметь консервативные или неконсервативные аминокислотные замены. Положения, которые имеют консервативные аминокислотные замены, могут быть заменены аминокислотой того же типа, наблюдаемой в тех же положениях в природных белках. Примеры таких замен показаны в Таблице 1.
Модификации биологических свойств полипептидов, таких как CD20, с помощью замен выполняются отбором замен, которые заметно различаются по своему действию на сохранение (а) структуры полипептидного каркаса в области замены, например, в виде складчатой конформации, спиральной конформации или петлевой конформации, (б) степени гидрофобности молекулы в целевом сайте, или (с) объема боковой цепи. Природные остатки делят на группы в зависимости от общих свойств боковых цепей:
(1) гидрофобные: норлейцин, met, ala, val, leu, ile;
(2) нейтральные гидрофобные: cys, ser, thr;
(3) кислые: asp, glu;
(4) основные: asn, gln, his, lys, arg;
(5) остатки, влияющие на ориентацию цепи: gly, pro; и
(6) ароматические: trp, tyr, phe.
Неконсервативные замены представляют собой замену члена одного из этих классов на другой класс. Такие заменяющие остатки можно также ввести в сайты консервативных замен или в остальные (неконсервативные) сайты.
Варианты трансмембранного полипептида, такие как варианты CD20, можно получать известными методами рекомбинантной ДНК, такими как опосредуемый олигонуклеотидами (сайт-направленный) мутагенез, сканирование аланином, ПЦР мутагенез, сайт-направленный мутагенез (Zoller el al., 1987, Nucl. Acids Res., 10: 6487-6500), кассетный мутагенез (Wells et al., 1985, Gene, 34: 315), мутагенез с селекцией рестрикции (Wells et al., 1986, Philos. Trans. R. Soc. London SerA, 317: 415) и т.п.
В. Системы экспрессии
1. Клетки-хозяева
Настоящее изобретение включает способы продуцирования трансмембранных полипептидов в клетках-хозяевах и, в частности, в бактериальных клетках. Бактерии-хозяева, пригодные для продуцирования трансмембранных полипептидов, включают Escherichia, Enterobacter, Bacillus, Pseudomonas, Klebsiella, Proteus, Salmonella, Serratia, Shigella и т.п. Подходящие бактериальные хозяева включают Enterobacteria, такие как Escherichia coli, Shigella dysentariae, Klebsiella pneumoniae и т.п. Подходящие Escherichia coli хозяева включают штаммы W3110 (инвентарный номер в АТСС 27325), 294 (инвентарный номер в АТСС 31446), В, XI776 (инвентарный номер в АТСС 31537), 58F3 и т.п. Можно использовать мутантные клетки любых вышеприведенных бактерий. В качестве иллюстрирующего примера в данном описании приводится клетка-хозяин Е.coli штамма 58F3 (W3110-fhuAΔ (tonAΔ) phoAΔE15 lonΔ galE rpoHts(htpRts) ΔclpP laclq ΔompTΔ(nmpc-fepE) ΔslyD). Предполагается, что векторы, промоторы и т.п. можно использовать и модифицировать аналогично, чтобы содействовать оптимальному продуцированию трансмембранных белков, таких как CD20, в других бактериальных хозяевах.
При выборе бактерий для применения в способах по изобретению принимают во внимание реплицируемость репликона в бактериях. Например, виды Е. coli, Serratia и Salmonella подходят для использования в качестве хозяина, когда в качестве источника репликона применяют pBR322, pBR325, pACYC177, pKN410 и т.п.
2. Промоторы
Для фактического и эффективного продуцирования сложных трансмембранных полипептидов в клетках-хозяевах, таких как бактериальные клетки, промотор выбирают по низкой базальной активности. Вследствие того, что трансмембранные полипептиды обычно являются токсическими в отношении клеток-хозяев, даже низкий уровень экспрессии, вызванной базальной активностью промотора, может повлиять на жизнеспособность клеток-хозяев, вызывая пониженный рост клеток, пониженное продуцирование белка и пониженный выход. Желательно, чтобы промотор был "включен" только на короткий период, в то время как клетки-хозяева являются в достаточной степени взрослыми для того, чтобы были обеспечены высокий уровень продуцирования и хороший выход белка. Соответственно, промотор выбирают и/или создают по данному описанию, исходя из минимальной базальной активности.
Два традиционно применяемых промотора, сильных и, как утверждают, с строгим контролем, например, в бактериальных клетках, основаны на нативных Т7 и лямбда PL промоторах.
Сильный Т7 промотор выпускается в рЕТ векторах (Novagen, Stratagen и другие) и используется в штамме, имеющем ген Т7 РНК полимеразы, интегрированный в хромосому (Лямбда DE3 лизогенные штаммы). Т7 РНК полимераза находится под контролем lac промотора/оператора, и, наконец, Т7 промотор, функционально связанный с нужным геном, индуцируется IPTG. Эта система, отдельно представляющая собой достаточно ликовые (растекающиеся) и токсические белки, такие как "многопроходные" трансмембранные белки, вызывает проблемы, включая отсутствие роста перед индукцией. Для ужесточения контроля и уменьшения базального уровня экспрессии ингибитор Т7 РНК полимеразы, Т7 лизоцим, можно коэкспрессировать на отдельной совместимой плазмиде (pLysS и pLysE от Novagen) в той же самой клетке. Полученная система экспрессии (pET/DE3 штамм/pLys) все еще индуцируется помимо IPTG, причем высокий уровень Т7 РНК полимеразы превосходит более низкий уровень ингибитора Т7 лизоцима, включая Т7 промотор.
Лямбда PL промотор представляет собой второй сильный промотор, менее общеупотребительный. Он выпускается на продажном векторе pLEX (Invitrogen). PL промотор, функционально связанный с нужным геном, позиционирован в pLEX плазмиде, а cI репрессор, который контролирует PL промотор, интегрирован в хромосому штамма. cI репрессор находится под контролем trp промотора/оператора.
Известные индуцибельные бактериальные промоторы можно применять в способах по изобретению при условии, что промотор обладает низкой базальной активностью или создан так, чтобы понизить базальную активность, как представлено в данном описании. Такие примеры включают промоторы бета-лактамазы, лактозы и триптофановый промотор. Другие промоторы, пригодные для применения в конкретных систем клеток-хозяев, общеизвестны и доступны и могут быть созданы для понижения базальной активности, как представлено в данном описании.
3. Элементы контроля (регуляторные элементы)
Промотор phoA представляет собой строго контролируемый промотор с низкой базальной активностью в Е. coli. Промотор phoA позитивно регулируется через pho бокс, который связывает активатор phoB (см. Таблицу 5). Промоторная активность включается истощением фосфата в средах, например, разведением в ограниченной по фосфату среде. Несмотря на эти механизмы контроля, промотор phoA не проявляет какой-либо базальной активности.
Можно выбирать или создавать промотор, содержащий один или более элемент негативного контроля и один или более элемент позитивного контроля. См., например, позитивные или негативные контрольные элементы, приведенные в Neidhardt, Ed., 1996, Escherichia coli and Salmonella, Second Ed., ASM Press, Washington D.C.
Промоторы как с позитивными, так и с негативными контрольными элементами встречаются редко. metE промотор является одним таким примером. См., например, Neidhardt, Ed., 1996, Escherichia coli and Salmonella, Second Ed., p.p.1300-1309. Описания известных элементов позитивного и негативного контроля можно найти, например, в этом ссылочном материале. В некоторых вариантах изобретения промотор содержит, по меньшей мере, один позитивный и, по меньшей мере, один негативный контрольный элементы, которые обеспечивают прямой контроль базальной (основной) экспрессии. Известно расположение позитивного или негативного контрольного элементов в промоторе или рядом с промотором для осуществления дополнительной регуляции промотора, оно описано, например, в Escherichia coli and Salmonella (Supra) на страницах 1232-1245.
Элементы негативного контроля включают, например, lac репрессор/lac оператор, Е. coli trp peпpeccop/trp оператор, лямбда репрессор/оператор и т.п. Элементы позитивного контроля включают, например, pho бокс промотора Е.coli и варианты, которые связывают phoB, сайт связывания ДНК MalT промотора оперона мальтозы, сайт связывания ДНК AraC промотора оперона арабинозы и т.п. Например, можно создать phoA промотор, имеющий pho бокс в качестве элемента позитивного контроля, таким образом, чтобы он содержал гетерологичный негативный контрольный элемент, такой как lac оператор lac оператор индуцируется при добавлении IPTG.
Два обычно применяемых элемента позитивного контроля для промоторов представляют собой PhoB/pAo box и сайт связывания ДНК AraC/araI. Эти и многие другие позитивные и негативные регуляторные элементы транскрипции описаны, например, в Neidhardt, Supra. Применяемые обычно элементы негативного контроля включают lac peпpeccop/lac оператор, trp peпpeccop/trp оператор и лямбда репрессор/оператор.
4. Терминаторы транскрипции
Для того чтобы избежать считывания с применением различных промоторных систем, можно установить один или более терминатор транскрипции для того, чтобы прекратить считывание в процессе транскрипции до того, как будет достигнут промотор, функционально связанный с экспрессируемой нуклеотидной последовательностью. Например, Лямбда последовательность терминации транскрипции AACG CTCGGTTGCCGCCGGGCGTT TTTTATT (SEQ ID NO: 17) можно включить "апстрим" (выше) промотора phoA. Известны другие последовательности терминации транскрипции, такие как терминатор His оперона, и их можно использовать. Встроенные контрольные элементы можно позиционировать таким образом, чтобы они были функционально связаны с другими промоторными элементами, для контролируемой экспрессии трансмембранного полипептида.
Индукция промотора с помощью агента, который влияет на быстрое и строго контролируемое "включение" и не оказывает вредного воздействия на клетки-хозяева, также является желательным свойством промоторной системы. Для экспрессии в Е.coli и родственных бактериях промотор phoA обеспечивает строгий контроль экспрессии. При мутации с добавлением негативного контрольного элемента, такого как терминаторы транскрипции lac оператор и "апстрим" Лямбда (Lambda), базальная экспрессия с использованием промотора фактически была исключена, как показано ниже в разделе Примеры.
Применимые phoA промоторы включают phoA промотор (SEQ ID NO: 5) и мутантный phoA, содержащий один или более негативный контрольный элемент, такой как мутантный промотор phac (SEQ ID NO: 15) и/или один или более 3′-5′ ("апстрим") терминатор транскрипции, такой как в мутантном промоторе tphac (SEQ ID NO: 16). Можно использовать мутантные промоторы, созданные с целью понизить базальную промоторную активность.
В бактериальных хозяевах, отличных от Е.coli, бывает полезно заменить phoA промотор функционально эквивалентным индуцибельным промотором, выбранным или созданным с низкой базальной активностью, как известно, совместимый с выбранным бактериальным хозяином. Подходящие промоторы включают, но без ограничения, промоторные системы бета-лактамазы и лактозы, триптофановую промоторную систему или гибридные промоторы, такие как tac или trc промотор, который может быть мутантным, чтобы понизить базальную активность. Выбранный промотор может также быть мутантным, содержащим как позитивные, так и негативные регуляторные элементы. Например, можно создать естественно негативно регулируемый промотор, добавляя позитивную регуляцию заменой последовательность -35 бокс не- -35 консенсусной последовательностью, затем добавить последовательность позитивного регуляторного элемента, такую как pho бокс. Может пригодиться замена необязательных генов tPHK редких кодонов генами tPHK редких кодонов, известными как совместимые с выбранным бактериальным хозяином.
5. Векторы
Векторы, применимые для экспрессии трансмембранных полипептидов, обычно содержат строго контролируемый промотор, функционально связанный с полинуклеотидом, кодирующим трансмембранный полипептид. Такие плазмиды, как pBR322, pBR325, pACYC177 или pKN410, можно использовать в качестве каркаса векторов. В одном варианте изобретения плазмида pBR322 образует каркас (скелет).
Векторы для экспрессирования трансмембранных полипептидов обычно включают сильный промотор, негативные и позитивные контрольные элементы, терминаторы транскрипции и дополнительные элементы для строгого контроля и эффективной экспрессии и трансляции.
Векторы могут включать короткую последовательность, кодирующую аминокислотный лидер, расположенный непосредственно перед первым кодоном кодированного полипептида. Лидерная последовательность содействует правильной инициации трансляции, и обычно содержит около 6-12 аминокислот, и может содержать, например, 6, 7, 8, 9, 10, 11 или 12 аминокислот, хотя может содержать и более. Одним из примеров является последовательность MKHQHQQ (SEQ ID NO: 7), кодируемая нуклеотидной последовательностью: (ATG)AAACACCAACACCAACAA (SEQ ID NO: 28), как показано на Фигуре 21. Более длинная лидерная последовательность, например, содержащая 30-50 или более аминокислот, применима для того, чтобы содействовать элонгации трансляции трансмембранных полипептидов. См., например, trpLE лидеры LE и sLE, показанные на Фигуре 22, которые содержат последовательность инициации трансляции (TIS) и спейсерную последовательность для того, чтобы способствовать элонгации трансляции.
Вектор может также включать гены tPHK редких кодонов клетки-хозяина. Примеры генов tPHK редких кодонов для Е. coli включают, но без ограничения, argU, glyT и pro2.
6. Лидерная последовательность
В предпочтительном варианте изобретения на экспрессирование трансмембранных полипептидов лидерная последовательность содержит "сильную" последовательность инициации трансляции (TIS) и спейсерную последовательность, расположенную между TIS и первым трансмембранным сегментом (ТМ-1). Один лидер, применимый для экспрессии "многопроходных" трансмембранных полипептидов в Е.coli, содержит участок trpLE лидера. См., например, LE и sLE лидеры, кодирующие участок N-концевой области Е.coli irpE белка, раскрываемого ниже в разделе Примеры.
7. Последовательность инициации трансляции
Известные последовательности инициации трансляции можно использовать для повышения эффективности инициации трансляции гена. Энхансерная последовательность инициации трансляции может простираться, включая последовательности 5' и 3' от сайта связывания рибосом. По определению сайт связывания рибосом включает, минимум, область Шайна-Дальгарно и стартовый кодон до любых оснований в промежутке. Кроме того, энхансерная последовательность инициации трансляции может включать нетранслируемую лидерную последовательность и конец "апстрим" цистрона, и, таким образом, стоп-кодон трансляции. См., например, патент США 5840523.
Существует множество способов получать высокие интенсивности инициации трансляции, включая применение, примерно, первых 6-12 или около того кодонов белка, высокоэкспрессирующегося в клетке-хозяине. Например, в Е. coli несколько белковых лидеров, имеющих оптимальные области инициации трансляции в начале кодирующей последовательности, включают β-галактозидазу (Ruther et al., 1983, EMBO J., 2: 1791-1794), Протеин A (Nilsson et al., 1990, Methods Enzymol., 185: 144-161), Глутатион-S-трансфераза (Smith et al., 1988, Gene, 67: 31-40) и т.п. Другим примером является последовательность MGSSHHHHHH (SEQ ID NO: 33), например, кодируемая нуклеотидной последовательностью: ATGGGCAGCAGCCATCATCATCATCATCAT (SEQ ID NO: 34). См. также общий обзор о таких лидерах: Lavallie et al., 1995, Current Biology, 6:501-506.
Или же можно создать сильную последовательность TIS, например, как описано в 1990, Methods in Enzymol., 185:89-119. Сильную TIS можно также выбрать, как описано, например, в Yansura et al., 1992, Methods: A companion to Methods in Enzymology, 4: 151-158.
"Сильная последовательность инициации трансляции" обычно представляет собой последовательность кодонов, которые кодируют около 6-12 аминокислот (например, 6, 7, 8, 9, 10, 11 или 12 аминокислот). Последовательность может быть природной или созданной методами генетической инженерии и допускает высокую интенсивность инициации трансляции.
В одном варианте изобретения сильная последовательность инициации трансляции содержит первые девять аминокислот trp лидера (M)KAIFVLKGS (SEQ ID NO: 27), кодируемых нуклеотидной последовательностью: ATGAAAGCAATTTTCGTACTGAAAGGTTCA (SEQ ID NO: 35). Другие последовательности включают нуклеотидные последовательности, кодирующие первые 6-12 аминокислот β-галактозидазы.
8. Спейсерная последовательность
Спейсерная последовательность, отделяющая TIS от ТМ-1 транслируемого белка, применима для содействия элонгации трансляции трансмембранных белков. Подходящая спейсерная последовательность содержит минимильные барьеры для элонгации, например минимальные редкие аминокислоты, плохо связывается с рибосомами и обычно неструктурированна, например не складывается, чтобы способствовать, при необходимости, транслокации через мембрану и, таким образом, допускать высокую скорость элонгации трансляции. Высказано предположение, что "спейсерная последовательность" функционирует как буферное (амортизирующее) пространство, способствующее тому, чтобы согласовать замедление трансляции в первом трансмембранном сегменте без утраты рибосомной нагрузки в TIS с продолжением элонгации. Спейсер должен быть достаточно длинным для того, чтобы эффективно отделять TIS от ТМ-1, но все же недостаточно длинным для того, чтобы допустить фолдинг транслируемого полипептида. Спейсерная последовательность обеспечивает эффективную и быструю элонгацию трансляции, не нарушая нормальное внедрение белка в мембрану.
Спейсерная последовательность может содержать, например, последовательность из 50 или более аминокислот, например, 60 или более, 70 или более, 80 или более аминокислот, и, предпочтительно, менее 120 аминокислот. В одном варианте изобретения "спейсерная последовательность" является гидрофильной и может содержать, примерно, от 20% до 50% имеющих заряд аминокислот, например, около 30% - около 40% аминокислот, имеющих заряд.
В другом варианте изобретения спейсерная последовательность содержит, по меньшей мере, часть бактериального гена и может быть образована из последовательности, в природе обнаруженной в клетке-хозяине, например, Е ген в Е.coli trp опероне для экспрессии полипептидов в клетках Е.coli. Как описано ниже в разделе Примеры, лидеры LE и sLE содержат часть trpE гена.
9. Метка экспрессии
Как правило, метка экспрессии может являться фрагментом вектора, или частью ДНК полипептида, встроенной в вектор. Метки экспрессии служат для идентификации и выделения экспрессируемого белка. Примеры включают, но без ограничения, поли-His метки (патент США 4569794), HisGln метку, биотин, авидин и т.п. Такие метки хорошо известны и выпускаются промышленностью (см., например, Qiagen, Valencia, СА). Поли-His метка содержит несколько гистидиновых остатков, обычно 6-10 гистидиновых остатков. Полипептиды с His меткой можно обнаруживать, нанося образец на колонку, связанную с антителом к His метке или на никелевую колонку.
На Фигурах 8, 21 и 22 графически изображены типичные экспрессирующие конструкции, применимые в способах по изобретению для экспрессии трансмембранных полипептидов. Например, на Фигуре 8 показаны функционально связанные последовательности промотора, лидерной последовательности, гена трансмембранного белка, метки экспрессии, генов терминатора транскрипции (лямбда t0) и tPHK. Нижеприведенные Примеры показывают применение таких экспрессирующих конструкций для экспрессии CD20, RA1c, GPR73 и MS4A4A.
С. Экспрессия трансмембранных полипептидов в бактериальных клетках
Экспрессированные трансмембранные полипептиды ассоциируются с мембраной бактериальной клетки в нативной конформации. Локализация трансмембранных полипептидов с мембранной бактериальной клетки можно определить, например, центрифугированием в градиенте плотности или другими известными методами.
Бактериальные хозяева культивируются в известных подходящих средах. Любые добавки к средам, помимо источников углерода, азота и неорганических фосфатов, включаются в подходящих концентрациях и вводятся индивидуально или в виде смеси с другой добавкой или средой, такой как источник комплексного азота. Клетки- хозяева культивируются при подходящих температурах. Например, Е.coli можно выращивать при комнатных температурах, примерно, от 20°С до 39°С, например, 25°С-37°С или около 30°С. рН культуральной среды может иметь любое значение, примерно, от 5 до 9, в зависимости от организма-хозяина. Культуральная среда для Е.coli может иметь рН, примерно, от 6.8 до 7.4, например, около 7.0.
Полинуклеотиды, кодирующие трансмембранные полипептиды, получают известными методами рекомбинантной ДНК. Эти методы включают, но без ограничения, выделение из природного источника, ПЦР, опосредуемый олигонуклеотидами (сайт-направленный) мутагенез, сканирование аланином, ПЦР мутагенез, сайт-направленный мутагенез (Zoller el al., 1987, Nucl. Acids Res., 10: 6487-6500), кассетный мутагенез (Wells et al., 1985, Gene, 34: 315), мутагенез с селекцией рестрикции (Wells et al., 1986, Philos. Trans. R. Soc. London SerA, 317: 415) и т.п.
Полинуклеотид, кодирующий трансмембранный полипептид, можно экспрессировать непосредственно или в виде слияния с другим полипептидом, или в виде полипептида, имеющего специфический сайт расщепления на С-конце зрелого полипептида, например.
В методах по изобретению используются стандартные методики рекомбинантной ДНК для получения трансмембранных полипептидов. Гетерологичный полинуклеотид, кодирующий трансмембранный полипептид (например, кДНК или геномная ДНК), включают в реплицирующийся вектор для экспрессии в бактерии. Многие векторы пригодны для этой цели, и селекция подходящего вектора зависит, главным образом, от размера нуклеиновой кислоты, встраиваемой в вектор, и конкретной клетки-хозяина, трансформируемой при использовании вектора. Каждый вектор содержит различные компоненты (фрагменты), зависящие от его функции (амплификация ДНК или экспрессия ДНК) и конкретной клетки-хозяина, совместимой с ним. Компоненты вектора для бактериальной трансформации обычно включают, но без ограничения, один или более следующих компонентов: ориджин репликации, один или более маркерных генов и индуцибельный промотор. Примеры подходящих векторов представлены в данном описании. В одном варианте изобретения векторы содержат промотор с высокой регуляцией, функционально связанный с геном, кодирующим трансмембранный полипептид. Примеры подходящих промоторов представлены в данном описании и включают промоторы phoA, phac и tphac промоторы, и другие такие промоторы под строгим контролем, например, как позитивного, так и негативного контрольных элементов. По данному описанию векторы могут содержать сильные последовательности инициации трансляции и спейсерные последовательности для повышения элонгации многопроходных трансмембранных полипептидов.
Как правило, применяют плазмидные векторы, содержащие репликон и контрольные последовательности видов, совместимых с бактериальной клеткой-хозяином. Обычно векторы несут сайт репликации, а также маркирующие последовательности, способные обеспечить фенотипическую селекцию в трансформированных клетках. Например, Е.coli обычно трансформируют, используя pBR322, плазмиду, образованную из видов Е.coli (см., например. Bolivar et al., 1977, Gene, 2: 95). Плазмида pBR322 содержит гены устойчивости к ампициллину и тетрациклину и поэтому обеспечивает простые способы идентификации трансформированных клеток. Плазмида pBR322, или другая плазмида, или фаг микробного происхождения обычно также содержат или модифицируются таким образом, чтобы содержать промоторы, которые могут использоваться микробным организмом для экспрессии селективных маркерных генов.
Как экспрессирующий, так и клонирующий векторы содержат нуклеотидную последовательность, которая дает возможность вектору реплицироваться в одной или более выбранных клеток-хозяев. Обычно в клонирующих векторах эта последовательность является последовательностью, которая позволяет вектору реплицироваться независимо от хромосомной ДНК хозяина, и индуцирует ориджин репликации или автономно реплицирующиеся последовательности. Такие последовательности хорошо известны для различных бактерий. Ориджин репликации из плазмиды pBR322 подходит для большинства грамотрицательных бактерий.
Экспрессирующий и клонирующий векторы обычно содержат также ген селекции, также называемый селективным маркером. Этот ген кодирует белок, необходимый для жизнеспособности или роста трансформируемых клеток-хозяев, выращенных в селективной культуральной среде. Клетки-хозяева, не трансформированные вектором, содержащим ген селекции, не выживают в культуральной среде. Типичные гены селекции кодируют белки, которые (а) придают устойчивость к антибиотикам или другим токсинам, например, к ампициллину, неомицину, метотрексату или тетрациклину, (б) восполняют ауксотрофную недостаточность, или (в) снабжают важными питательными веществами, недоступными из комплексных сред, например ген, кодирующий D-аланин рацемазу для Bacillus spp. В одном из примеров схемы селекции используется лекарственное вещество для прекращения роста клетки-хозяина. Клетки, которые успешно трансформированы с помощью гетерологичного гена, продуцируют белок, придающий устойчивость к лекарству и, таким образом, выдерживают схему селекции.
Промоторы можно индуцировать стандартными методами. См., например, Sambrook el al., Molecular Cloning: A Laboratory Manual (New York: Cold Spring Harbor Laboratory Press, 1989). Обычно бактериальные клетки культивируют до достижения определенной оптической плотности, в этот момент инициируют индукцию, вводя индуцирующий фактор (индуктор), используя истощение компонента среды, или с помощью того и другого, в соответствии с требованиями выбранного промотора. Промотор phoA индуцируется при истощении фосфата, как описано, например, в: Chang et al., 1987, Gene, 55: 189-196; Simmons et al., 2002, J. Immunol. Methods, 263: 133-147; и/или патенты США 5304472 и 5342763.
Когда промотор содержит как позитивные, так и негативные контрольные элементы, например, pho бокс и lac оператор, имеющиеся в мутантных промоторах phac и tphac, желательно согласовывать индукцию промотора с помощью обоих контрольных элементов. Например, индукцию путем удаления негативного контрольного элемента в lac операторе при добавлении IPTG можно согласовывать с низкой точкой истощения среды фосфатом, так чтобы оба контрольных элемента "включали" промоторную активность одновременно или как можно ближе по времени друг к другу. Продолжительность направляемой промотором экспрессии обычно ограничена по времени, чтобы сохранить жизнеспособность экспрессирующих клеток, например, менее 3 часов, менее 2 часов, или какое-то время между 1 и 2 часами. Длительность экспрессии может меняться в зависимости от клетки-хозяина и конкретного экспрессируемого полипептида.
Клетки лизируют, разделяют растворимую и нерастворимую фракции и трансмембранные полипептиды экстрагируют из нерастворимой мембранной фракции. Типичные методы солюбилизации обсуждаются ниже.
Генную экспрессию можно измерять в образце опосредованно (непрямыми методами), обычным Нозерн-блоттингом, для количественного определения транскрипции tPHK (Thomas, 1980, Proc. Natl. Acad. Sci. USA, 77: 5201-5205). Можно использовать различные метки, наиболее часто, радиоизотопы, в частности,32Р. Также можно использовать другие методы, такие как мечение биотином. Модифицированные биотином нуклеотиды, введенные в полинуклеотид, могут служить в качестве сайта связывания с авидином или антителами, которые можно метить разнообразными метками, такими как радионуклеотиды, флуоресцирующие вещества, ферменты и т.п. Генную экспрессию можно также измерять прямыми методами, анализируя экспрессированные полипептиды, например, Вестерн-блоттингом.
D. Выделение и очистка трансмембранных полипептидов
Трансмембранные полипептиды можно выделять из клеток-хозяев, таких как бактериальные клетки, не содержащие клеток или клеточных мембран, способами по данному описанию, и растворимые в детергенте, сохраняя достаточную "нативную" конформацию, так что полипептиды можно распознавать в виде иммуногена или в виде полипептида, связанного с лигандом. Выделенный полипептид сохраняет достаточную "нативную" конформацию для того, чтобы связываться с лигандом, связывание которого зависит от структурного признака в нативной конформации полипептида. Например, связывание ритуксимаба с CD20 зависит от внеклеточной петли В в нативной конформации CD20 (например, когда CD20 экспрессируется на клеточной мембране). CD20, солюбилизованный в неионном или в цвиттер-ионном детергенте по данному описанию, содержит в детергенте структуру петли В, достаточно "нативную" для того, чтобы связывать ритуксимаб или его антиген-связывающий фрагмент, такой как фрагмент Fab.
1. Дезинтеграция клеток-хозяев
Клетки-хозяева, в которых находятся (укрывающие) трансмембранные полипептиды, можно разрушать различными физическими или химическими методами, включая, но без ограничения, цикл замораживания-оттаивания, ультразвук, механическая дезинтеграция, агенты, лизирующие клетки, и т.п. Трансмембранный полипептид можно высвобождать из клетки или клеточной мембраны с помощью подходящего детергента или ферментативным расщеплением. Трансмембранные полипептиды регенерируются из дезинтегрированных клеток путем солюбилизации в детергенте.
2. Мягкие неденатурирующие детергенты
Обычный метод солюбилизации мембранных белков представляет собой использование мягких неденатурирующих детергентов, таких как додецилмальтозид, н-додецил-N,N-диметиламин-N-оксид, н-додецилфофохолин (FOS-Choline-12) и тритон X-100. Вариантом этого метода является применение смешанных мицелл, состоящих из детергента и липида (обычно в соотношении 10:1). Такая методика недавно была применена при выделении и кристаллизации калиевого канала Kv1.2 (Long et al., 2005, Science 309 (5736): 897-903 и Long et al., 2005, Science 309 (5736): 903-8). Сильные денатурирующие детергенты, такие как SDS, не будучи предпочтительными, могут использоваться для солюбилизации и денатурации полипептидов. Далее, перед биохимическим исследованием белок обычно реконструируют в более мягкий детергент, или липосому, или другую, не поверхностно-активную среду. Однако может быть трудно восстановить нормальную функцию после экспозиции белков с денатурирующими детергенами.
3. Неионный и цвиттер-ионный детергент
В одном варианте изобретения трансмембранные полипептиды солюбилизируют в неионном или в цвиттер-ионном детергенте. Примеры неионных детергентов, применимых для солюбилизации сложных трансмембранных полипептидов, таких как СВ20, включают, но без ограничения, TRITON® и додецилмальтозид. Примеры цвиттер-ионных детергентов, применимых для солюбилизации сложных трансмембранных полипептидов, таких как СВ20, включают, но без ограничения, цвиттергенты, такие как цвиттергент 3-08, 3-10, 3-12, 3-16 (Cal Bio Chem), ASB-14, ASB-16, ASB-C80 (Anatrace), PMAL-B 100 и производные фосфохолина, такие как додецилфосфохолин (DDPC), и т.п. Фосфохолиновые детергенты содержат холин, имеющий четвертичную аммониевую группу с положительным зарядом, связанную с фосфатом, являются ионными и полярными. Производные содержат неполярные углеводородные цепи, соединенные с фосфатом, например, додецил для DDPC. Применимые производные фосфохолина включают, например, DDM, DM, LADO, DDPC, DHPC, LOPC, LMPC, DLPC, LLPG (выпускаемые Avanti Polar Lipids (Alabaster, AL) и/или Anatrace Inc. (Maumee, ОН)), и н-децил-N,N-диметиламин-N-оксид.
4. Ионный или денатурирующий детергент
Экстракция ионными или денатурирующими детергентами может не дать выделенные трансмембранные полипептиды, сохраняющие достаточную для применения "нативную" конформацию. Ресуспендирование выделенных полипептидов в неионном или цвиттер-ионном детергенте позволяет реконструировать полипептид и дает трансмембранные полипептиды с подходящей "нативной" конформацией. Например, трансмембранные полипептиды можно экстрагировать из клеточных мембран ионным детергентом. Для применения, например, в иммунном анализе ионный детергент можно заменять на неионный или цвиттер-ионный детергент.
5. Очистка в малых масштабах
Методы солюбилизации и очистки могут в большой степени зависеть от масштабов экстракции и очистки. В случае экстракции и очистки в малых масштабах, например, около 1 г клеток, клеточный осадок можно инкубировать в детергенте, а растворимые и нерастворимые фракции можно регенерировать без стадий дополнительной очистки.
6. Очистка в больших масштабах
Для крупномасштабной экстракции и очистки, например, около 100 г клеток или более, клетки можно смешивать с детергентом и центрифугировать. Полученный супернатант можно очищать, используя известные методы, включая, но без ограничения, фракционирование на ионообменной колонке; аффинную хроматографию, например, при использовании антитела против His-метки или антитела против CD20; осаждение этанолом; обращенно-фазовую хроматографию; хроматографию на силикагеле или на катионообменной смоле, такой как DEAE; хроматофокусирование; SDS-PAGE; осаждение сульфатом аммония; гель-фильтрацию, например, на Sephadex® G-75; колонки с Протеин А Сефарозой® для удаления примесей, таких как IgG; колонки с хелатируемым металлом для связывания форм полипептидов, меченных эпитопом, смолы с аффинностью к гидрофобным соединениям, аффинность к лигандам с применением подходящего лиганда, иммобилизованного на матрице, центрифугирование в градиенте плотности сахарозы и т.п. Известны и могут применяться различные методы очистки белков. См., например, Deutscher, 1990, Methods in Enzymology, 182; Scopes, 1982, В: Protein Purification: Principles and Practice, Springer-Verlag, New York; Ausubel et al., (ed.), 1998, В: Current Protocols in Molecular Biology, John Wiley & Sons.
7. His-метка ("хвост")
В одном варианте изобретения трансмембранные полипептиды являются меченными His-меткой. Очистку меченных His-меткой полипептидов можно осуществлять, например, пропуская солюбилизированную в детергенте фракцию через колонку с хелатируемым металлом, например, такую как колонка Ni-NTA, или колонку, содержащую антитела к His. После захвата полипептиды, меченные His-меткой, элюируют подходящим буфером. В одном варианте буфер содержит 0.1% н-додецил-β-D-мальтозид, 150 мМ NaCl и 20 мМ цитрата натрия, рН 3.5.
8. Аффинная очистка
В другом варианте изобретения трансмембранные полипептиды очищают аффинной очисткой солюбилизированной в детергенте фракции, пропуская фракцию через колонку с иммобилизованным лигандом, который связывает "нативную" структуру полипептида. Например, CD20 можно очистить от детергента с помощью ритуксимаба. Элюированные фракции, содержащие трансмембранный полипептид, можно далее концентрировать и очищать гель-фильтрацией, аффинной очисткой и т.п. Концентрации белка можно определять различными общеизвестными методами, например, такими как определение с помощью ВСА (Smith et al., 1985, Anal Biochem., 150: 76-85).
9. Обработка не-детергентами
После очистки белков при желании их можно обрабатывать в отличной от детергентов среде. Наиболее обычная отличная от детергентов среда представляет собой липосому, которая лучше имитирует нативную клеточную среду, чем мицеллы детергента. Липосомы состоят, главным образом, из длинноцепных липидов. См., например, Rigaud et al., 1995, Biochim Biophys Acta. 1231(3): 223-46 и Ollivon et al., 2000, Biochim Biophys Acta. 1508(1-2): 34-50. Бицеллы представляют собой вариант липосом, образующийся, главным образом, из бислоя, образующего липид, смешанного с короткоцепным липидом. См., например, Czerski and Sanders, 2000, Anal Biochem. 284(2): 327-33.
Е. Методы с использованием выделенных белков
1. Созревание аффинности/Селекция антител
Трансмембранные полипептиды, экспрессированные, выделенные и/или очищенные по данному описанию, имеющие подходящую "нативную" конформацию, можно применять в качестве целевых антигенов для селекции вариабельных доменов антител методом фагового дисплея и для созревания аффинности антител. Известны методы фагового дисплея вариабельных доменов антител и селекции специфических связующих для антигена. Антитело со "зрелой аффинностью" содержит одно или более изменений в гипервариабельной области, которые дают в результате повышенную аффинность антитела к антигену по сравнению с родительским антителом, в котором нет изменения (изменений). Антитела со зрелой аффинностью могут иметь наномолярную или даже пикомолярную аффинность к целевому антигену.
Фаговый дисплей представляет собой метод, с помощью которого вариантные полипептиды визуализуются в виде белков, слитых, по меньшей мере, с участком оболочечного белка на поверхности фаговых частиц, например частиц нитевидного фага. Использование фагового дисплея основано на том факте, что из больших библиотек рандомизированных белковых вариантов можно быстро и эффективно выбрать последовательности (провести быстрый и эффективный сортинг), которые связываются с целевым антигеном с высокой аффинностью. Дисплей пептидных и белковых библиотек на фаге использовался для скрининга миллионов полипептидов на полипептиды со свойствами специфического связывания. Методы поливалентного фагового дисплея применялись для визуализации малых случайных пептидов и малых белков с помощью слияния либо с геном III, либо с геном VIII нитевидного фага. Wells and Lowman, Curr. Opin. Struct. Biol., 3:355-362 (1992), и приведенные в этой статье ссылки. При одновалентном (моновалентном) фаговом дисплее белковую или пептидную библиотеку сливают с геном III или его участком и экспрессируют с очень низкими уровнями экспрессии в присутствии белка дикого типа, кодируемого геном III, так что фаговые частицы визуализуют одну копию слитых белков или совсем не визуализуют слитых белков. Эффекты авидности снижаются по сравнению с поливалентным фагом, так что сортинг проводят, исходя из внутренней аффинности лигандов, и используются фаг(е)мидные векторы, которые упрощают операции с ДНК. Lowman and Wells, Methods: A companion to Methods in Enzymology, 3: 205-0216 (1991). Типичные методы фагового дисплея вариабельных доменов антитела можно найти в опубликованной патентной заявке США No. 2005-0119455-A1, которая в виде ссылки вводится в данное описание.
Marks et al., 1992, Bio/Technology, 10:779-783, описывает созревание аффинности с помощью "перетасовки" экзонов VH и VL доменов (шаффлинг). Случайный мутагенез остатков CDR и/или каркаса описан Barbas et al., 1994, Proc. Nat. Acad. Sci. USA, 91: 3809-3813; Scier et al., 1995, Gene, 169: 147-155; Yelton et al., 1995, J. Immunol., 155: 1994-2004; Jackson et al., 1995, J. Immunol., 154: 3310-3319; и Hawkins et al., 1992, J. Mol. Biol., 226: 889-896.
"Созревание аффинности при использовании фагового дисплея" (AMPD) относится к процессу, описанному Lowman et al., 1991, Biochemistry 30(45): 10832-10838. См. также Hawkins et al., 1992, J. Mol. Biol. 226, 889-896 и патент США 6172213. Не ограничивая строго нижеприведенное описание, этот процесс можно кратко описать следующим образом: проводят мутации в нескольких сайтах гипервариабельной области (например, в 6-7 сайтах) с целью получения всех возможных аминокислотных замен в каждом сайте. Мутанты антитела, полученные при этом, визуализуют одновалентным способом при использовании фаговых частиц нитевидного фага в виде слияний с генным III продуктом М13, агрегированным с каждой частицей. Фаг, экспрессирующий различные мутанты, можно ввести в цикл при использовании раундов селекции на связывание с последующим выделением и секвенированием тех мутантов, которые проявляют высокую аффинность. Этот метод описан также в Международной заявке WO 92/09690. Модифицированная методика, включающая фаговый дисплей антител низкой аффинности, описана в Cunningham et al., 1994, EMBO J. 13(11), 2508-2515.
Созревание аффинности методом фагового дисплея обеспечивает селекцию новых связывающих полипептидов. Например, используя следующие стадии:
а) конструирование реплицируемого экспрессирующего вектора, содержащего первый ген, кодирующий полипептид, второго гена, кодирующего, по меньшей мере, часть оболочечного белка природного фага или фага дикого типа, причем первый и второй ген являются гетерологичными, а регуляторный элемент транскрипции функционально связан с первым и вторым генами, тем самым образуя генное "слияние", кодирующее слитый белок;
б) мутирование вектора в одном или более выбранных положений внутри первого гена, тем самым образуется семейство родственных плазмид;
в) трансформацию подходящих клеток-хозяев с помощью плазмид;
г) инфицирование трансформированных клеток-хозяев хелперным фагом, имеющим ген, кодирующий оболочечный белок фага;
д) культивирование трансформированных инфицированных клеток-хозяев в условиях, подходящих для образования рекомбинантных фаг(е)мидных частиц, содержащих, по меньшей мере, часть плазмиды и способных трансформировать хозяина, при этом условия корректируют таким образом, чтобы не более чем минорное количество фаг(е)мидных частиц визуализировало более одной копии слитого белка на поверхности частицы;
е) контактирование фаг(е)мидных частиц с целевой молекулой таким образом, что, по меньшей мере, часть фаг(е)мидных частиц связывается с целевой молекулой; и
ж) отделение связывающихся фаг(е)мидных частиц от несвязывающихся.
Методы созревания аффинности могут дополнительно содержать трансформацию подходящих клеток-хозяев рекомбинантными фаг(е)мидными частицами, которые связываются с целевой молекулой, и повторение стадий г)-ж) один раз или более.
Или же, метод включает полипептиды, которые состоят из более чем одной субъединицы, причем реплицируемый вектор экспрессии, содержащий регуляторный элемент транскрипции, функционально связанный с ДНК, кодирующей нужную субъединицу, слит с оболочечным белком фага.
Или же, поливалентный фаг (McCafferty et al., 1990, Nature 348, 552-554; Clackson et al., 1991, Nature 352, 624-628) также можно использовать для экспрессии случайных точковых мутаций, например, образующихся при использовании способствующей ошибкам ДНК полимеразы, с целью генерации библиотеки фрагментов фагового антитела, которые затем можно подвергать скринингу на аффинность к антигену (Hawkins et al., 1992, J. Mol. Biol. 226: 889-896).
2. Методы скрининга
Можно провести точный и высокочувствительный скрининг для идентификации и/или количественного определения нужной целевой молекулы, такой как молекула человеческого, человеческого-химерного или гуманизированного антитела, или фрагмент таких антител. Трансмембранные полипептиды получали, как представлено в данном описании, например, в виде специфических в отношении мишени иммобилизованных полипептидов.
Применимый метод анализа обычно включает следующие стадии: (1) реакцию трансмембранного полипептида с целевой молекулой в образце; и (2) количественное определение связанной целевой молекулы. Трансмембранный полипептид можно иммобилизовать на поверхности в виде захваченного реагента.
3. ELISA
Иммуноаналитические системы включают, например, твердофазный ELISA и метод захвата ELISA. В методе захвата, разновидности ELISA, иммобилизация трансмембранных полипептидов на твердой фазе осуществляется известными методами. Полипептид может сорбироваться твердой фазой, которая содержит анализируемую поверхность или матрицу (см., например, патент США 3720760). Полипептид может связываться, нековалентной или ковалентной связью, с аналитической (анализируемой) поверхностью, с предварительной активацией подложки или без предварительной активации. Отложение трансмембранного полипептида в виде иммобилизованного реагента может также представлять собой иммунопреципитацию, например, после связывания антитела в образце. В одном варианте изобретения трансмембранный полипептид иммобилизуют разведением раствора полипептида до величины мицелл ниже критического значения. Отложение растворимого белка на анализируемой поверхности можно осуществлять в мягких денатурирующих условиях, например в слабоосновной или слабокислой среде. Или же белок может захватываться за счет ковалентного связывания на поверхности анализа, или связываться с помощью белка, такого как антитело, расположенное на анализируемой (аналитической) поверхности.
В одном варианте изобретения реагент для захвата представляет собой трансмембранный полипептид, такой как CD20, в выделенной нативной конформации, получаемый способами по изобретению. Также можно использовать фрагменты полипептида. Трансмембранный полипептид связывает антитело из образца.
Твердая фаза, используемая для иммобилизации, может представлять собой любую инертную подложку или носитель, практически нерастворимые в воде и применяемые в иммуноанализах, включая подложку в виде, например, поверхностей, частиц, пористых матриц и т.п. Примеры обычно употребляемых подложек включают малые пластины, Сефадекс, поливинилхлорид, пластиковые гранулы, микрочастицы, аналитические планшеты, пробирки из полиэтилена, полипропилена, полистирола, и т.п. Такие подложки включают 96-луночные микротитрационные планшеты, а также конкретные материалы, такие как фильтровальная бумага, агароза, сшитый декстран и другие полисахариды. Или же, реакционно-способные нерастворимые в воде матрицы, такие как углеводы, активированные цианогенбромидом, и реакционно-способные субстраты, описанные в патентах США 3969287; 3691016; 4195128; 4247642; 4229537 и 4330440, подходят для иммобилизации реагента для захвата. Иммобилизованный реагент для захвата можно наносить на микротитрационный планшет. Твердой фазой может являться многолуночный микротитрационный планшет, который можно использовать для одновременного анализа нескольких образцов.
Твердую фазу покрывают реагентом для захвата, который, при желании, может быть связан за счет нековалентного или ковалентного взаимодействия или физического связывания. Методы связывания включают методы, описанные в патенте США 4 376 110 и в приведенных в нем ссылках.
Если используют планшеты из полистирола или полипропилена, лунки в планшете можно покрывать реагентом для захвата (обычно разведенным в буфере, таком как 0.05 М карбонат натрия) инкубацией, по меньшей мере, в течение, примерно, 10 часов, например, в течение ночи, при температурах около 4-20°С, например, 4-8°С, и при рН около 8-12, например, в интервале 9-10 или около 9.6. Если желательно более короткое время покрытия (?~2 часа), планшет можно покрывать при 37°С или на дне его может находиться нитроцеллюлозный фильтр, например, можно использовать Millipore MULTISCREEN™ (Billerica, MA). Трансмембранные белки могут поступать на аналитическую поверхность в виде растворимых в детергенте белков. Разведение детергентом ниже критической величины мицелл вызывает осаждение полипептида на аналитической поверхности.
Покрытые планшеты обычно обрабатывают блокирующим агентом, который неспецифически связывается с сайтами связывания и насыщает их, предупреждая нежелательное связывание свободного лиганда с лишними сайтами связывания на стенках планшета. Блокирование обычно происходит при комнатной температуре в течение, примерно, 1-4 часов, например, в интервале 1.5-3 часа.
После нанесения покрытия и блокирования сывороточный образец для анализа разводят соответствующим образом и добавляют к иммобилизованной фазе. Степень разведения обычно составляет около 5-15%, например, 10 объемных %. Чтобы обеспечить достаточную чувствительность, иммобилизованный реагент для захвата можно брать в молярном избытке максимальной молярной концентрации аналита, определяемом в образце после соответствующего разведения. Условия инкубации образца и реагента для захвата выбирают таким образом, чтобы сделать чувствительность анализа максимальной, а диссоциацию минимальной. Время инкубации зависит, прежде всего, от температуры.
Образец отделяют от иммобилизованного реагента для захвата (улавливающего) раствором для отмывания, удаляя незахваченный аналит из системы. Раствор для отмывания обычно представляет собой буфер.
Как правило, систему можно отмывать три раза. Температура раствора для отмывания обычно около 0-40°С, например, в интервале 4-30°С. Можно использовать автоматический промыватель (вошер) для микропланшетов. К раствору для отмывания можно добавлять сшивающий агент или другой подходящий агент для того, чтобы связать ковалентной связью уловленный (иммобилизованный) аналит с реагентом для захвата.
После удаления незахваченных молекул аналита из системы захваченные (иммобилизованные) молекулы аналита контактируют с детектирующим реагентом, таким, например, как антитело, при комнатной температуре. Температура и время контактирования аналита с детектирующим агентом зависит, главным образом, от используемого метода обнаружения. Например, когда пероксидаза хрена (HRP), конъюгированная с антителом овцы к мышиному IgG, применяется как способ обнаружения, детектирующий агент можно инкубировать с захваченным аналитом в течение, примерно, 0.5-2 часа, например, около 1 часа. Систему отмывают, как указано выше, удаляя несвязанный детектирующий агент из системы, и проявляют, добавляя субстрат пероксидазы и инкубируя планшет при комнатной температуре в течение, примерно, 5 минут или до проявления четкого цвета.
Молярный избыток детектирующего агента можно добавлять к системе после того, как несвязанный аналит вымыт из системы. Детектирующий реагент может представлять собой поликлональное или моноклональное антитело, или их смесь. Детектирующий реагент можно обнаруживать непосредственно или опосредованно.
Аффинность детектирующего реагента достаточно высока, так что можно обнаруживать некие количества аналита. Флуоресцентная или хемилюминесцентная метка имеет более высокую чувствительность по сравнению с обычной колориметрической меткой. Следует рассматривать аффинность связывания выбранного детектирующего агента с учетом аффинности связывания агента для захвата, так чтобы детектирующий реагент не отбирал аналит от реагента для захвата.
Метка (меченая частица) представляет собой детектируемую функциональность, которая не мешает связыванию захваченного аналита с детектирующим агентом.
Примеры подходящих меченых частиц включают частицы, которые можно обнаружить непосредственно, такие как люминофорная, хемилюминесцентная и радиоактивная метки, а также частицы, такие как ферменты, для обнаружения которых нужно, чтобы они прореагировали или были дериватизированы. Примеры таких меток включают, но без ограничения, радиоизотопы32Р,14С,125I,3H и131I, люминофоры, такие как хелаты редкоземельных металлов или флуоресцеин и его производные, родамин и его производные, люциферазы, например, люцифераза светляков и бактериальная люцифераза (патент США 4737456), пероксидаза хрена (HRP), щелочная фосфатаза и фермент, который использует пероксид водорода для окисления предшественника красителя, такой как НРР, биотин/авидин, биотин/стрептавидин и т.п.
Конъюгация меченой частицы с детектирующим агентом, таким как антитело, является стандартным методом иммуноанализа. См., например, O′Sullivan et al., 1981, Methods in Enzymology, 73: 147-166. Для ковалентного связывания меченой частицы с белками или полипептидами применяют обычные методы.
Количество аналита, связанного с улавливающим реагентом (реагентом для захвата), можно определить, отмывая несвязанный детектирующий агент от иммобилизованной фазы и измеряя количество детектирующего агента, связанного с аналитом, методом обнаружения, пригодным для данной метки. В одном варианте изобретения меченая частица представляет собой фермент. В случае ферментных частиц количество проявленного цвета является прямым методом измерения количества захваченного аналита. Например, когда меченой частицей является HRP, цвет обнаруживают, количественно рассчитывая оптическую плотность (O.D.) при 650 нм. В другом варианте изобретения количество аналита, связанного с улавливающим реагентом (реагентом для захвата), можно определять опосредованно.
4. Получение антител
Трансмембранный полипептид, такой как CD20, солюбилизированный в детергенте, можно использовать непосредственно в качестве иммуногена для генерации антител к трансмембранному белку. Можно также использовать другие методы получения антител, включая методы фагового дисплея по данному описанию.
Антитело получают против антигена из млекопитающего первого вида, например млекопитающее первого вида может быть человеком. Однако рассматриваются другие млекопитающие, такие как сельскохозяйственные, домашние животные или животные из зоопарка, например, если предполагается, что антитело используется для лечения таких млекопитающих.
Для получения мутантного антитела в известной аминокислотной последовательности делают несколько изменений аминокислот (например, вводят замену, делению, добавление).
(i) Приготовление антигена. Трансмембранный антиген, применяемый для продуцирования антител, может представлять собой растворимую форму полноразмерного полипептида или его фрагмента, например солюбилизированная полноразмерная молекула или фрагмент, такой как внеклеточный домен трансмембранного полипептида.
(ii) Поликлональные антитела. Поликлональные антитела обычно получают в организме животных с помощью множественных подкожных (sc, пк) или интраперитонеальных (и.п.) инъекций релевантного антигена и адъюванта. Они могут применяться для конъюгации релевантного антигена с белком, иммуногенным в виде, который иммунизируют, например, гемоцианином лимфы улитки, сывороточным альбумином и т.п.
Животных можно иммунизировать антигеном, иммуногенными конъюгатами или производными, смешивая, например, 100 мкг или 5 мкг белка или конъюгата (для кроликов или мышей соответственно) с 3 объемами полного адъюванта Фрейнда и инъецируя раствор интрадермально в нескольких местах. Через месяц животных иммунизируют, вводят вторичную дозу в количестве от 1/5 до 1/10 первоначального количества пептида или конъюгата в полном адъюванте Фрейнда в виде подкожной инъекции в нескольких местах. Через 7 или 14 дней у животных отбирают кровь и анализируют титр антител. Животным делают бустерные инъекции (иммунизируют) до достижения плато на кривой титра. Например, животных можно иммунизировать конъюгатом того же антигена, но конъюгированного с другим белком и/или с помощью другого сшивающего агента. Конъюгаты можно также получать в рекомбинантной клеточной культуре в виде слитых белков. Также для усиления иммунного ответа годятся агрегирующие агенты, такие как квасцы.
(iii) Моноклональные антитела. Моноклональные антитела можно получать, используя метод гибридом, впервые описанный Kohler et al., 1975, Nature, 256: 495, или методами рекомбинантной ДНК, например методами, описанными в патенте США 4816567, или другими известными методами.
По методу гибридом мышь или другое подходящее животное-хозяин, такое как хомяка или макак, иммунизируют, как описано выше, для выявления лимфоцитов, продуцирующих или способных продуцировать антитела, которые специфически связываются с белком, применяемым для иммунизации. Затем лимфоциты сливаются с клетками миеломы с помощью подходящего фактора, вызывающего слияние клеток, такого как полиэтиленгликоль, с образованием клетки гибридомы (Goding, 1986, Monoclonal Antibodies: Principles and Practice, pp.59-103, Academic Press). Полученные таким образом клетки гибридомы засевают в подходящую культуральную среду, например, содержащую одно или более веществ, которые ингибируют рост или жизнеспособность неслитых родительских клеток миеломы, и выращивают в этой среде.
После того, как идентифицированы клетки гибридомы, которые продуцируют антитела заданной специфичности, аффинности и/или активности, клоны можно субклонировать методом ограниченных разведений и выращивать стандартными методами (Goding, 1986, см. выше). Моноклональные антитела, секретируемые субклонами клеток, соответствующим образом выделяют из культуральной среды, асцита или сыворотки соответствующими методами очистки иммуноглобулинов, такими, например, как аффинная хроматография на протеин А-Сефарозе®, колоночная хроматография с гидроксилапатитом, гель-электрофорез, диализ, аффинная хроматография и т.п.
ДНК, кодирующая моноклональные антитела, легко выделяется и секвенируется обычными методами. В одном примере применяют олигонуклеотидные зонды, способные специфически связываться с генами, кодирующими тяжелые и легкие цепи моноклональных антител, клетки гибридомы могут служить в качестве источника такой ДНК.
Аффинность связывания моноклонального антитела можно определять, например, методом Скэтчарда, описанным в Minson et al., 1980, Anal. Biochem., 107: 220.
(iv) Гуманизация и варианты аминокислотной последовательности. Примеры метода гуманизации антител приведены в патенте США 6037454 (антитела к CD11a), патенте США 6329509 (антитела к IgE), патенте США 5821337 (антитела к p185HER2) и в Международной заявке WO 98/45331 (антитела к VEGF) и вводятся в данное описание в качестве ссылки.
Как описано ранее, для выбора антител, специфических в отношении трансмембранных белков по данному описанию, можно использовать ряд методов, таких как фаговый дисплей, традиционная иммунизация, созревание аффинности и другие методы.
Все публикации, патенты и патентные документы вводятся в данное описание в виде ссылки, как если бы они вводились ссылкой индивидуально. Раскрытие описано со ссылкой на различные конкретные и предпочтительные варианты изобретения и методы. Разумеется, однако, что можно осуществить многие варианты и модификации, не отходя от сущности и объема изобретения.
Примеры
Изобретение легче понять с помощью нижеприведенных Примеров, являющихся иллюстративными и никоим образом не служащими для ограничения.
Пример 1
Клонирование и экспрессия CD20 при использовании phoA промотора
Материалы
Все детергенты получают от Anatrace Inc., Maumee, ОН. Если не указано иначе, все химические реактивы получают от Sigma-Aldrich, St. Louis, МО. Полноразмерное антитело ритуксимаб получают от Genentech Manufacturing. Fab ритуксимаба экспрессируют в Е.coli и очищают с помощью протеина А и катионообменной хроматографией. Экспрессирующие конструкции, применяемые в нижеприведенных Примерах, графически изображены на Фигурах 8 и 21, а применяемые лидерные последовательности показаны на Фигуре 22. Если не указано иначе, используют Е.coli клетки штамма 58F3.
Клонирование и экспрессия
кДНК человеческого и мышиного CD20 субклонируют стандартными методами молекулярной биологии (Ausubel et al., eds., 2003, Current Protocols in Molecular Biology, 4 Vols., John Wiley & Sons) в плазмиду, образованную из BR322, содержащую ген β-лактамазы и гены tPHK трех редких для Е.coli кодонов (argU, glyT и pro2). Добавляют короткие полинуклеотиды для кодирования лидерной последовательности, MKHQHQQ (SEQ ID NO: 7) на N-конце CD20 для гарантии инициации высокого уровня трансляции и для кодирования последовательности метки, окта-His (SEQ ID NO: 8) на С-конце с целью содействовать обнаружению и очистке экспрессированного белка. Транскрипция гена осуществляется под контролем phoA промотора, а экспрессию индуцируют, лимитируя среду по фосфату. Насыщенную LB карбенициллином культуру разводят в C.R.A.P. фосфат-лимитирующих средах (Simmons et al., 2002, J. Immunol. Methods, 263: 133-147). После этого культуру выращивают при 30°C в течение 24 часов.
Мутантный CD20 продуцируют, заменяя остатки Cys111 и Cys220 на серин методом сайт-направленного мутагенеза с образованием C2S мутанта. Мутант проверяют на улучшенные свойства белка по сравнению с немутантным CD20, включая агрегацию, экспрессию, растворимость и сохранение нативной конформации. Экспрессию CD20 в ферментере осуществляют, как описано в Simmons et al., 2002, см. выше.
Выделение белка
Для анализа различных детергентов на способность солюбилизироватъ меченный His человеческий CD20, экспрессируемый в E.coli, 5 г клеток ресуспендируют в гомогенизаторе Polytron (Brinkmann, Westbury, NY) в 50 мл буфера А (20 мМ Tris, pH 8.0, 5 мМ EDTA) и центрифугируют при 125000×g в течение 1 часа. Затем клеточный осадок ресуспендируют в буфере А, лизируют дезинтеграцией клеток на микрофлюидизаторе (Microfluidics Corp, Newton, MA) и центрифугируют при 125000×g в течение 1 часа. Клеточный осадок отмывают один раз в том же буфере без EDTA и центрифугируют как описано выше. Осадок ресуспендируют в 20 мл буфера В (20 мМ Tris, pH 8.0, 300 мМ NaCl), отбирают аликвоты и испытуемые детергенты добавляют к аликвотам в следующих концентрациях:
1% SDS;
1% н-лаурилсаркозин;
1% н-додецил-N,N-диметиламин-N-оксид (LADO);
1% додецилфосфохолин (DDPC, Fos-Choline® 12);
1% н-додецил-β-D-мальтозид (DDM);
1% Тритон® Х-100;
2.5% CHAPS.
Клеточный осадок экстрагируют в течение ночи при 4°С, за исключением SDS образца, который экстрагируют при комнатной температуре. На следующий день образцы центрифугируют и супернатанты удаляют. Осадки и супернатанты ресуспендируют в восстанавливающем SDS лодинг-буфере (буфере для нанесения образца) до равных объемов и анализируют методом SDS-PAGE и иммуноблоты на нитроцеллюлозных мембранах анализируют с помощью конъюгированных с пероксидазой хрена антител к His (Roche Applied Science, Indianapolis, IN).
Для экстракции в больших масштабах лизируют 100-200 г клеток и нерастворимую фракцию готовят, как описано выше. Для экстракции CD20 из нерастворимой фракции конечный клеточный осадок ресуспендируют в буфере В при соотношении около 1:2.5 вес./об., от начального сырого веса клеток, добавляют DDPC до 1% и раствор перемешивают в течение ночи при 4°С. На следующий день нерастворимую в детергенте фракцию центрифугируют на ультрацентрифуге при 125000×g в течение 1 часа. Супернатант наносят на колонку Ni-NTA Superflow (Qiagen Inc. Valencia, CA), предварительно уравновешенную буфером В и 5 мМ DDPC. Колонку промывают буфером А, содержащим 20 мМ имидазола в объеме, равном 10 объемам колонки, и связанный белок элюируют буфером А, содержащим 250 мМ имидазола. Все стадии очистки путем нанесения на колонку проводят при 4°С.
Фракции элюента, содержащие CD20, концентрируют и нагружают на колонку Superdex 200 (Amersham Biosciences, Piscataway, NJ), предварительно уравновешенную в буфере А с 5 мМ DDPC. Меченный His человеческий CD2Q и мышиный CD20 дополнительно очищают на колонке HiTrap HP Q на 5 мл (Amersham Biosciences, Piscataway, NJ) перед гель-фильтрацией. Для обмена детергента образцы пропускают через колонку Superdex 200 в буфере С, (0.1% DDM, 150 мМ NaCl, 20 мМ HEPES, pH 7.2.). Или же, образцы связывают с маленькой колонкой Ni-NTA, промывают буфером С и элюируют буфером С, содержащим 300 мМ имидазола. Эти образцы затем подвергают диализу против буфера С для удаления имидазола.
Для аффинной очистки человеческого CD20 ритуксимаб иммобилизуют в количестве 6 мг/мл на 10 мл смолы Actigel ALD Superflow (Sterogene, Carlsbad, CA). Эту смолу помещают в колонку и уравновешивают в буфере (0.1% DDM, 150 мМ NaCl, 20 мМ HEPES, pH 7.2). Человеческий мутантный CD20, C2S, экспрессируют и очищают, как описано выше, для нативного человеческого CD20. Очищенный C2S мутант пропускают через колонку и несвязанный белок удаляют интенсивной промывкой буфером В. Белок элюируют в 0.1% DDM, 150 мМ NaCl и 20 мМ цитрата натрия, pH 3.5. Элюированные образцы сразу же нейтрализуют, концентрируют и диализуют против буфера С. Концентрацию белка определяют с помощью ВСА, как описано в Smith et al., 1985, Anal. Biochem., 150: 76-85 (Pierce Biotechnology, Rockford, IL 61101). До момента использования образцы хранят при -80°С.
Центрифугирование в градиенте плотности
Очистку в ступенчатом градиенте плотности осуществляют, помещая слои 1.9 М, 1.4 М и 0.8 М растворы сахарозы, забуференные 150 мМ NaCl и 20 мМ HEPES, pH 7.2, в центрифужные пробирки. Клетки, экспрессирующие белок CD20, лизируют дезинтеграцией в буфере А, содержащем 1 мМ EDTA. Нерастворимую фракцию выделяют центрифугированием при 38,000×g в течение 1 часа. Супернатант отбрасывают и осадок ресуспендируют в буфере для лизиса с добавлением 0.25 М сахарозы в соотношении 1:5 вес./об., 100 мкл (микролитров) этой "ресуспензии" смешивают с 0.9 мл 1.9 М раствора сахарозы. Конечная концентрация сахарозы в полученной смеси составляет 1.75 М. Эту смесь затем помещают на дно центрифужной пробирки, а выше помещают слои по 1 мл остальных двух растворов сахарозы. Затем наверху центрифужных пробирок помещают последний слой 200 мкл (микролитров) 0.25 М раствора сахарозы. Образцы помещают в ротор SW55 и центрифугируют в течение 1 часа при 100,000 × g. Затем образцы осторожно выгружают в аликвоты 200 мкл (микролитров) из верхней части пробирки и анализируют методом SDS-PAGE, переносят на нитроцеллюлозу и анализируют с помощью антитела против His, конъюгированного с пероксидазой хрена.
Анализы ELISA
Белок CD20 иммобилизуют на 96-луночные планшеты в течение ночи при 4°С, используя 100 мкл (микролитров) CD20 при концентрации 1 микрограмм/мл в PBS, содержащем солюбилизирующий детергент, разведенный до концентрации мицелл ниже критической. Затем планшеты трижды отмывают PBS, содержащим 0.05% Твин-20 (PBST), и в течение 45 при комнатной температуре блокируют с помощью 200 мкл PBST, содержащим 0.5% BSA (блокирующий и аналитический буфер). Планшеты снова трижды отмывают PBST, а затем анализируют с применением первичного антитела. Объем 150 мкл (микролитров) ритуксимаба при концентрации 60 мкг/мл (микрограмм/мл) в буфере для анализа добавляют в соответствующие лунки и делают трехкратные серийные разведения в последующих лунках, отбирая 50 мкл из первой лунки и смешивая со 100 мкл (микролитрами) аналитического буфера в следующей и в дальнейших лунках до конечной концентрации около 2 нг/мл.
После инкубации при комнатной температуре в течение 90 минут планшеты отмывают PBST и связанный ритуксимаб обнаруживают, добавляя 100 мкл конъюгата пероксидазы хрена с антителом козы к человеческому F(ab′)2 (Jackson ImmunoResearch Laboratories Inc., West Grove, PA), разведенного 1:2000 в аналитическом буфере, отмывают шесть раз с помощью PBST и проявляют, добавляя 100 мкл/лунка системы субстрата ТМВ Microwell Peroxidase Substrate System (KPL, Gaithersburg, MD), смешанной в соответствии с инструкциями производителя. Реакцию останавливают, добавляя 100 мкл/лунка 1.0 М фосфорную кислоту (100 мкл/лунка), и измеряют оптическую плотность при 450 нм на планшет-ридере.
Образцы восстановленного и алкилированного CD20 готовят восстановлением, добавляя 10 мМ DTT, и алкилированием, добавляя 25 мМ йодацетамида. Реакцию прекращают, добавляя еще 100 мМ DTT. После каждой стадии реакционную смесь оставляют на 30-60 минут при комнатной температуре при рН 8.0. Для восстановления и повторного окисления (реокисления) образец CD20 восстанавливают, добавляя перед засеванием 10 мМ DTT, и оставляют на несколько часов на планшете в отсутствие DTT для повторного окисления перед связыванием с антителом.
Поверхностный плазменный резонанс
Аффинность и кинетику связывания ритуксимаба с выделенными человеческими CD20 белками определяют на приборе BIAcore™-3000 (BIAcore, Inc. Piscataway, NJ). Сенсорный датчик СМ5 активируют для ковалентного связывания ритуксимаба или ритуксимаба Fab с помощью N-этил-N′-(3-диметиламинопропил)-карбодиимида гидрохлорида и N-гидроксисукцинимида в соответствии с инструкциями производителя. Ритуксимаб или Fab ритуксимаба разводят в 5-10 раз до концентрации 100 мкг/мл в 10 мМ растворе ацетата натрия, рН 5.0, и вводят в активированный датчик (чип). Остальные активные сайты связывания блокируют с помощью 1 М раствора этаноламина. Интактный ритуксимаб осаждается при 8000-12000 RU (единиц ответа, относительных единиц), a Fab ритуксимаба осаждается при 4000-7000 RU.
Для кинетических измерений семь двукратных разведений (всего восемь образцов) человеческого CD20 начальной концентрации 5 мкМ в 0.1% DDM, 150 мМ NaCl и 20 мМ HEPES, рН 7.2, при 25°С вводят при скорости потока 30 мкл/минута на 100 секунд. Связанный белок оставляют диссоциировать в течение 720 секунд. В конце измерения каждого образца сенсорные поверхности регенерируют, инъецируя 20 мкл 10 мМ HCl. После корректировки сенсограмм на сигналы от эталонного потока проводят расчеты кинетики, используя простую 1:1 модель с помощью программы BIAevaluation 3.0 (BIAcore).
Круговой дихроизм (КД)
Растворы CD20 в детергенте, либо в 0.1% DDPC, либо в 0.1% DDM, готовят диализом против 100 мМ раствора фосфата натрия, рН 7.2, и либо 0.1% DDPC, либо 0.1% DDM. Результаты кругового дихроизма, полученные на приборе AVIV202 на образцах 2-5 мкМ белка в кварцевой кювете 1 мм; проход длины волны осуществляют при 25°С в указанных областях, инкременте 2 нм, при среднем времени 10 секунд. По данным строят кривую в интервале от 185 до 285, но за исключением образцов, содержащих β-меркаптоэтанол. Так как β-меркаптоэтанол мешает регистрации данных при более низких длинах волн, регистрацию этих данных прекращают при 200 нм.
Анализ связывания ритуксимаб IgG и Fab с нормальными человеческими В клетками методом Скэтчарда
Константы равновесной диссоциации (Кd) определяют для связывания ритуксимаба IgG и фрагмента Fab ритуксимаба с В клетками, используя белок, меченный радиоактивной меткой. Все разведения осуществляют в буфере для анализа связывания (среды DMEM, содержащие 0.5% бычий сывороточный альбумин, 25 мМ HEPES, рН 7.2, и 0.01% азид натрия). Аликвоты (0.05 мл) ритуксимаб125I-IgG, йодированного йодогеном (Iodogen), или125I-Fab, йодированного йодогеном (Iodogen) или лактопероксидазой, помещают в лунки для анализа с М-образным дном 96-луночного планшета для микроанализа с концентрацией 0.005 или 0.05 нМ соответственно. Добавляют серийные разведения (0.05 мл) холодного антитела и перемешивают. Затем в лунки добавляют очищенные человеческие В-клетки (125000 в 0.05 мл). Планшеты герметично закрывают и инкубируют при комнатной температуре в течение 24 часов, затем центрифугируют 15 минут при 2500 об/мин. Супернатант отсасывают, а клеточный осадок отмывают и центрифугируют. Супернатант снова отсасывают, а клеточный осадок растворяют в 1 N NaOH и переносят в пробирки для гаммаметрии. Данные используют для анализа Скэтчарда, описанного в Munson et al., 1980, Anal. Biochem., 107: 220-239, с помощью программы Ligand, описанной в McPherson, 1983, Comput. Programs. Biomed., 17: 107-113.
Нормальные человеческие В клетки выделяют из 100 мл гепаринизированной крови здорового человека негативной селекцией, используя смесь RosetteSep™ В Cell Enrichment Cocktail (Stemcell Technologies, Vancouver, Canada) в соответствии с протоколом производителя. Далее В клетки отделяют центрифугированием через слой Ficoll-Paque (Amersham Biosciences, Peapack, NJ), а затем выделяют и отмывают в фосфатно-солевом буферном растворе. Все оставшиеся эритроциты лизируют, в течение 30 секунд экспонируя с гипотоническим раствором. Затем доводят концентрацию очищенных В клеток до 1-2 миллионов клеток на миллилитр связывающего буфера.
Экспрессия мембраносвязанного человеческого CD20 в Е.coli
Первичная структура человеческого CD20 показана на Фигуре 1. Предполагаемая топология CD20 представляет собой топологию четырежды пронизывающего мембрану белка с обоими концами в цитоплазме. Две внеклеточных петли CD20 поразительно отличаются размерами. Первая петля, между спиралью один и спиралью два, чрезвычайно мала и невероятным кажется, что она значительно выступает из мембраны. Размер этой петли в высшей степени сохраняется в других членах семейства MS4A (см., например, Ishibashi et al., 2001, Gene, 264: 87-93 и Liang et al., 2001, Genomics, 72:119-127).
Вторая петля, между спиралями три и четыре, имеет длину около 46 аминокислот, и тянется от области остатка 140 до области остатка 185, и содержит одну возможную дисульфидную связь между остатками 167 и 183. Размер этой петли меняется в широких пределах в аминокислотных последовательностях генов семейства MS4A, хотя большая часть этих последовательностей сохраняет способность образовывать внеклеточную дисульфидную связь (Ishibashi et al., 2001, Supra и Liang et al., 2001, Supra). На цитоплазматической стороне мембраны в покоящихся В клетках CD20 фосфорилирован (Valentine et al., 1989, J. Biol. Chem., 264: 11282-11287). Фосфорилирование увеличивается в ответ на сшивание антитела (Tedder et al., 1988, J. Biol. Chem., 263: 10009-10015). Никакие другие посттрансляционные модификации не идентифицированы на CD20, а в человеческом белке отсутствуют какие-либо консенсусные сайты N-гликозилирования во внеклеточной области.
Для подтверждения структуры CD20 и возможной роли образования дисульфидной связи в связывании антител меченный His человеческий белок CD20 экспрессируют в Е. coli, как описано выше. Возможность нативной экспрессии в Е. coli оценивают локализацией клеточной экспрессии CD20 на мембране с помощью центрифугирования в градиенте плотности и тестирования растворимости белка в нативных детергентах.
На Фигуре 2 показана локализация выделенного CD20 в градиенте сахарозы. Фракции 1-16 указаны над гелем на панели а. Аликвоты фракций градиента сахарозы подвергают электрофорезу на SDS-PAGE геле. Гель блоттируют и анализируют с помощью антитела к His метке. Фракции представляют собой фракцию с самой низкой плотностью сахарозы (1) до фракции с самой высокой плотностью сахарозы (14). Найдено, что примерно половина всего белка, экспрессированного в бактериях, локализована во фракции сахарозы (12), имеющей плотность меньше 1.28 г/см2 (1.75 М сахарозы), как показано на Фигуре 2, панель а, тогда как остальная часть обнаружена в клеточном осадке (фракция 16).
Плотность типичных растворимых белков равна 1.33-1.42 г/см2 (Creighton, 1993, Proteins Structures and Molecular Properties, 1 Ed., W.H.Freeman and Company, New York, USA), что выше плотности в нижнем слое сахарозы (1.28 г/см2, 1.75 М). Растворимые белки можно обнаружить в нижней части градиента, тогда как мембраносвязанные белки всплывают к более низкой плотности из-за липида вокруг белка. Эти результаты, показанные на Фигуре 2, панель а, согласуются с локализацией человеческого CD20 во фракции клеточной мембраны бактерий, так как сообщалось, что плотность Е.coli мембран 1.15-1.25 г/см2 (Ishidate et al., 1986, J. Biol. Chem., 261: 428-443). Эти наблюдения согласуются также с предыдущими сообщениями, в которых описана экспрессия других эукариотических мембранных белков в нативной конформации в клеточных мембранах бактерий (Berlin el al., 1992, J. Biol. Chem., 267: 8200-8206; Grisshammer et al., 1993, Biochem J., 295(Pt2): 571-576).
Солюбилизация CD20 детергентом
Детергенты, пригодные для солюбилизации CD20, определяют скринингом набора неденатурирующих и денатурирующих детергентов методами по данному описанию. Клеточный осадок и супернатант мембран клеток Е.coli после экстракции детергентом анализируют с антителом к His. На Фигуре 2, панель b, показаны результаты этого скрининга. В Вестерн-блоте человеческого CD20 супернатанты, полученные при использовании различных детергентов, помечены (S), тогда как клеточный осадок помечен (Р). (WC) обозначает экстракт цельных клеток (контроль).
Различные детергенты, используемые для экстракции белка из мембран, представляют собой SDS (1), н-лаурилсаркозин (2), н-додецил-N,N-диметиламин-N-оксид (LADO) (3), н-додецилфосфохолин (DDPC) (4), н-додецил-β-D-мальтозид (DDM) (5), Тритон-Х 100 (6) и CHAPS (7). Как показано на Вестерн-блоте, растворимый белок получают, используя разнообразные детергенты. Существенная часть CD20 растворима в неденатурирующем цвиттер-ионном детергенте додецилфосфохолине (DDPC) (4). Этот детергент выбирают для дальнейшей работы по экстракции и очистке CD20.
Меченный His человеческий CD20 экстрагируют из Е.coli мембран с помощью DDPC и очищают металл-хелатирующей хроматографией, эксклюзионной хроматографией по размеру и анионообменной хроматографией, описанными выше. Около 10-20 мкг очищенного меченного His белка CD20 получают из одного грамма бактериальных клеток.
На Фигуре 3 показаны окрашенные Кумасси синим дорожки SDS гелей экстрагированного и очищенного меченного His человеческого белка CD20, мутанта C2S и мышиного CD20. На дорожках 1, 2 и 3 показаны невосстановленные белки: человеческий CD20 (дорожка 1), C2S мутант (дорожка 2) и мышиный CD20 (дорожка 3). Дорожка 4 содержит маркеры молекулярной массы (Mark 12, Invitrogen). Дорожки 5, 6 и 7 изображают восстановленные белки: человеческий CD20 (дорожка 5), C2S мутант (дорожка 6) и мышиный CD20 (дорожка 7). Дорожки 8 и 10 показывают невосстановленный (дорожка 8) и восстановленный (дорожка 10) мышиный CD20, прилегающие к маркерам молекулярной массы (дорожка 9). Каждая дорожка содержит 2 мкг (микрограмма) белка. Молекулярные массы белковых маркеров составляют 200, 116, 97, 66, 55, 36, 30, 22, 14 и 6 кДа. Типичный SDS-PAGE очищенного меченного His человеческого CD20 показан на Фигуре 3 (дорожки 1 и 5). Белок движется со средней молекулярной массой около 38 кДа в восстанавливающих условиях, что корректно согласуется с расчетной молекулярной массой 35 кДа.
В очищенном CD20 также наблюдается незначительное изменение подвижности в невосстанавливающих и восстанавливающих условиях на SDS-PAGE. Это отчетливо видно на Фигуре 3, где восстановленный (дорожка 10) и невосстановленный (дорожка 8) мышиный CD20 идут на соседних дорожках для сравнения. Это изменение наводит на мысль, что невосстановленный CD20 существует в виде более компактной, быстрее движущейся структуры, чем восстановленная форма, из-за дисульфидной связи, расположенной в большой внеклеточной петле. Эта дисульфидная связь исчезает при добавлении восстановителя.
Хотя детергенты додецилмальтозид (DDM) и LADO демонстрируют лишь ограниченную способность солюбилизировать CD20, предпринята попытка очистки в больших масштабах с применением этих растворителей, чтобы оценить, действительно ли солюбилизация детергентом из Е. coli достаточно точно количественно определяется Вестерн-блоттингом в условиях, описанных выше и показанных на Фигуре 2, панель b. Коротко говоря, экспрессируют нерастворимые фракции Е. coli, меченный His человеческий CD20 экстрагируют с помощью 1% указанного детергента. Образцы центрифугируют и клеточный осадок и супернатанты суспендируют в равных объемах SDS буфера. Равные объемы каждого образца подвергают электорофорезу на SDS-PAGE в восстанавливающих условиях. Для сравнения равный объем фракции цельных клеток (WC) суспендируют в SDS буфере после лизиса, но не проводят никаких операций.
Белок, очищенный с применением либо DDM, либо LADO, является значительно менее чистым, и метод дает значительно меньше белка, чем очистка, осуществляемая с применением DDPC. Однако, CD20 может успешно обмениваться в неионные детергенты после очистки, это показывает, что способность DDPC солюбилизировать CD20 не является уникальной.
Экспрессия мышиного CD20
Мышиный CD20 экспрессируют и очищают в условиях, аналогичных условиям экспрессии и очистки человеческого CD20. Результаты этой очистки показаны на Фигуре 3 (дорожки 3 и 7). Белковый материал мышиного белка имеет лучшие свойства, чем человеческий CD20, проявляя меньшую агрегацию на невосстанавливающих SDS гелях (сравните дорожки 1 и 3, Фигура 3) и давая более высокий конечный выход белка. Изучение первичной последовательности мышиного и человеческого CD20 показывает, что остаток цистеина 111 (Фигура 1) в человеческой последовательности заменяется на серин в мышином белке. Эта замена наводит на мысль, что Cys 111 не является существенным для активности CD20. Кроме того, показано, что цистеин 220 не является существенным для активности CD20, так как замена Cys220 на аланин дает в результате аналогичную экспрессию и зависящую от антитела ассоциацию с липидным рафтом, что и белок дикого типа при экспрессии в эукариотических клетках. См., например, Polyak et al., 1998, J. Immunol., 161: 3242-3248.

C2S Мутант
Для оценки экспрессии и связывания CD20 с антителом осуществляют мутацию с заменой остатков Cys111 и Cys220 с заменой на серин с образованием C2S мутанта. C2S мутант экспрессируют и очищают, как описано выше, для меченного His человеческого CD20. C2S мутант проявляет улучшенные свойства белка по сравнению с нативным белком, включая пониженную агрегацию и повышенный выход.
На панели с Фигуры 2 показаны образцы Е.coli клеточных экстрактов, демонстрирующие экспрессию меченного His человеческого CD20 и C2S мутанта CD20. Дорожки 1 и 4 содержат контрольный, холостой вектор; Дорожки 2 и 5 содержат человеческий CD20; и Дорожки 3 и 6 содержат C2S мутант CD20. Образцы на дорожках 1, 2 и 3 пропускают в невосстанавливающих условиях; Образцы на дорожках 4, 5 и 6 восстанавливают с помощью 100 мМ DTT. На каждую дорожку наносят равный объем клеток, нормализованных по оптической плотности.
Уровень экспрессии C2S мутанта (дорожки 3 и 6) в Е.coli выше, а дисульфид-зависимая агрегация меньше, чем у нативного человеческого CD20 (дорожки 2 и 5). Меньшая агрегация и, примерно, в два раза более высокий выход белка в случае C2S мутанта показан также на Фигуре 3, панель а, где дорожки 2 и 6, содержащие C2S, сравниваются с нативным человеческим CD20 (дорожки 1 и 5) и мышиным CD20 (дорожки 3 и 7).
Экспрессированный rCD20 связывает ритуксчмаб
Чтобы проверить, действительно ли очищенный рекомбинантный CD20 принимает нативную конформацию, анализируют способность очищенного белка связывать химерное антитело к CD20, ритуксимаб. Известно, что это антитело связывается со структурно напряженной внеклеточной петлей человеческого CD20, экспрессируемого на поверхности В клеток (Polyak et al., 2002, Blood, 99: 3256-3262). Метод ELISA осуществляют с учетом связывания ритуксимаба с очищенным человеческим CD20. Результаты показаны на Фигуре 4, кривые изображают дисульфид-зависимое связывание антитела ритуксимаба с меченным His человеческим CD20 (закрашенные квадраты), восстановленным и алкилированным hCD20 (незакрашенные, полые квадраты), восстановленным и повторно окисленным hCD20 (закрашенные кружки) и PBS контроль (полые кружки). Кривые, проходящие через точки связывания с ритуксимабом, строят оптимизацией по 4 параметрам.
Как показано на Фигуре 4, в этом анализе ритуксимаб связывается с меченным His человеческим CD20 с наномолярной аффинностью. Значение ЕС50 9.4 нМ получают оптимизацией данных по 4 параметрам.
Восстановленный rCD20 не связывается с ритуксимабом в анализе ELISA
Связывание с ритуксимабом локализовано во внеклеточной петле CD20 между остатками К142 и Y184 (Polyak el al., 2002, Supra). Полагают, что два остатка в этой области, С167 и С183, образуют дисульфидную связь (Einfeld et al., 1988, EMBO J., 7: 711-717). Полагают, что связывание ритуксимаба в свою очередь в высшей степени зависит от присутствия этой дисульфидной связи. Чтобы оценить важность любое значение связи в CD20 для связывания ритуксимаба, CD20 восстанавливают, и алкилируют, и анализируют связывание с ритуксимабом. Эти превращения значительно уменьшили способность ритуксимаба связывать CD20. В качестве дополнительного контроля восстанавливают CD20, удаляют DTT и белок повторно окисляют. Связывание ритуксимаба частично восстанавливается в результате этой процедуры, в соответствии с корректировкой дисульфидной связи в CD20, это демонстрирует, что связывание с антителом зависит от образования дисульфидной связи. Антитела к внеклеточной области мышиного CD20 в настоящее время отсутствуют, невозможно провести аналогичный анализ для мышиного CD20.
BIAcore анализ rhCD20 и C2S мутанта
В методе анализа ELISA белка CD20, описанном выше, можно принимать в расчет эффекты авидности при связывании с антителом. Чтобы оценить связывание ритуксимаба с человеческим CD20 независимо от таких эффектов, осуществляют анализ методом поверхностно-плазменного резонанса. Этот метод имеет дополнительное преимущество, давая как информацию о кинетическом связывании, так и равновесные константы связывания. В этих экспериментах ритуксимаб или Fab ритуксимаба осаждают на сенсорном чипе BIAcore и растворимый человеческий CD20 в различных концентрациях пропускают через этот чип. Полноразмерный ритуксимаб осаждается при 10000 RU, а человеческий CD20 в указанных концентрациях наносят на сенсорный чип при скорости потока 20 мкл/минута в растворе 150 мМ NaCl, 20 мМ HEPES (рН 7.2) и 0.1% DDM.
Интересно отметить, что хотя CD20 можно выделять в DDPC, связывание CD20 с иммобилизованным ритуксимабом значительно понижается в присутствии этого детергента (данные не приводятся). Поэтому аффинности в экспериментах по поверхностно-плазменному резонансу определяют в присутствии DDM.
Типичные данные этих экспериментов можно видеть на Фигуре 5, на BIAcore сенсограмме, показывающей связывание между ритуксимабом и человеческим меченным His CD20. Связывание человеческого CD20 с иммобилизованным ритуксимабом происходит при концентрациях CD20 в интервале от 0.04 мкМ до 5.0 мкМ, включая 5 мкМ, 2.5 мкМ, 1.25 мкМ, 0.63 мкМ, 0.31 мкМ, 0.16 мкМ, 0.08 мкМ и 0.04 мкМ. Концентрации первых 4 образцов помечены на сенсограмме. Связывание ритуксимаба показано для невосстановленного CD20 (закрашенные квадраты), восстановленного и алкилированного CD20 (закрашенные кружки), восстановленного CD20, который окислен повторно (полые квадраты), и контрольного PBS (незакрашенные кружки), кривые связывания строят оптимизацией по четырем параметрам.
В Таблице 3 приведены параметры связывания ритуксимаб IgG или Fab ритуксимаба с человеческим CD20, C2S мутантом и очищенным аффинной хроматографией C2S мутанта. Данные являются репрезентативными из нескольких экспериментов и оптимизированы по модели с единственным сайтом связывания. Величины Kd и Ка рассчитывают из скоростей ассоциации и диссоциации. На этой таблице можно видеть, что оба меченных His белка, человеческий CD20 и C2S мутант человеческого CD20, проявляют, примерно, одинаковый свойства связывания с полноразмерным антителом ритуксимабом и с Fab фрагментом. Данные демонстрируют, что мутации Cys- на- Ser в CD20 не изменяют связывания с антителом, например, не изменяют "нативную" конформацию, необходимую для связывания антитела.
hCD20, экспрессированный в E.coli, находится в "нативной" конформации
Для определения процентного содержания CD20 в подходящей конформации, присутствующего в наших препаратах, мы дополнительно очищаем C2S мутант человеческого CD20 на аффинной колонке с ритуксимабом. Хотя выходы низкие, результаты связывания до и после аффинной очистки в целом согласуются, демонстрируя, что основная часть очищенного человеческого CD20 находится в конформации, способной связывать ритуксимаб. Незначительное повышение связывания антитела продемонстрировано после аффинной очистки с иммобилизованным ритуксимабом. Эта повышенная аффинность может быть вызвана либо повышенной чистотой CD20, либо удалением неактивных молекул CD20. Наблюдается небольшая разница в аффинности CD20 к полноразмерному антителу по сравнению с Fab. Эту незначительную разницу можно отнести за счет поверхностных эффектов вследствие связывания меньшего фрагмента Fab с сенсорным чипом или вследствие минорных изменений структуры Fab после удаления области Fc.
Возможно, любой CD20, агрегированный в мицеллу детергента, может внести вклад в эффект аффинности связывания как IgG, так и Fab. Хотя влиянием эффектов авидности трудно управлять, полагают, что они вносят незначительный вклад в связывание, наблюдаемое в данном методе анализа, по двум причинам. Во-первых, между аффинностью растворимого CD20 к интактному ритуксимабу IgG или к фрагменту Fab наблюдается разница менее чем в два раза. Даже если связывание Fab допускает некоторые эффекты авидности вследствие близости Fab фрагментов на сенсорном чипе, маловероятно, что случайное связывание и ориентация фрагмента Fab предусматривают такое близкое согласие измерений, кроме того, как результаты для IgG, так и результаты для Fab показывают прекрасное соответствие с теоретической моделью, предсказанной для одновалентного связывания. Отклонение от теоретического одновалентного связывания должно быть особенно очевидно для IgG BIAcore эксперимента, однако, как показано на Фигуре 5, теоретическая одновалентная модель и действительные экспериментальные данные для связывания CD20 с ритуксимабом IgG очень хорошо согласуются. Таким образом, не требуется теоретически допускать никаких дополнительных вариантов с учетом экспериментальных данных.
Второе, показатели аффинности как ритуксимаба IgG, так и фрагмента Fab, методом BIAcore, хорошо соответствуют показателям аффинности фрагмента Fab по определению методом Скэтчарда связывания на нормальных человеческих В клетках. Связывание ритуксимаба IgG или Fab с нормальными человеческими В клетками определяют конкуренцией за немеченый ритуксимаб IgG или Fab против125I-IgG или125I-Fab. Показанные результаты являются средними в анализе на нормальных клетках от двух доноров. Типичные кривые замещения в экспериментах по связыванию IgG и Fab показаны на Фигуре 7. Связывание определяют конкуренцией немеченого ритуксимаба IgG с125I-IgG за донор 1 (верхняя панель) или немеченого Fab ритуксимаба с125I-Fab за донор 4 (нижняя панель), каждое измерение делают в тройном повторе на клетках единственного донора. В Таблице 4 показана аффинность и число рецепторов от каждого донора. Величина ЕС50 для нативного CD20 в этом анализе равна 9.5 нМ.
Результаты показывают, что между аффинностью одновалентного Fab ритуксимаба к выделенному человеческому CD20, по определению методом поверхностно-плазменного резонанса (BIAcore) и аффинностью одновалентного Fab ритуксимаба к CD20, экспрессированному на выделенных человеческих В-клетках, существует малая разница, в три-четыре раза (аффинность 190-280 нМ в BIAcore экспериментах по сравнению с аффинностью 50-60 нМ при анализе методом Скэтчарда). Эта малая разница может объясняться, помимо других причин, имманентными различиями методов анализа, физическими различиями в окружении белка, присутствием детергента или отсутствием партнеров CD20 по связыванию в изолированном материале. Интересно отметить, большое различие между связыванием IgG и Fab с В клетками. Это наводит на мысль, что эффекты авидности могут играть роль в связывании ритуксимаба с В клетками.
Как следовало ожидать, учитывая Kd ритуксимаба с человеческим CD20 (84-170 нМ), скорости ассоциации и диссоциации связывания относительно высоки, в частности, по сравнению с высокоаффинными антителами, такими как зрелое по аффинности антитело против VEGF (Chen et al., 1999, J. Mol. Biol., 293: 865-881), имеющее Kd менее 0.15 нМ, kon 3.6×104 (М-1 сек-1) и koff, менее или равную 0.05×104 (сек-1) при 25°С. Однако, так как аффинность ритуксимаба IgG или фрагмента Fab ритуксимаба, определяемые с помощью BIAcore к выделенному CD20, хорошо соответствуют аффинности Fab ритуксимаба, определенной методом Скэтчарда на нормальных В клетках, кажется вероятным, что низкая величина одновалентной (моновалентной) аффинности является правдоподобной, а не получается в результате большого числа неправильных или ненативных конформаций в препаратах выделенного hCD20.
Анализ вторичной структуры методом кругового дихроизма
Далее анализируют вторичную структуру CD20 методом кругового дихроизма (КД) человеческого C2S и мышиного CD20. Простые спектры, полученные в результате этого анализа, показаны на Фигуре 6.
На верхней панели показаны спектры мутантного C2S человеческого CD20 в присутствии 0.1% DDPC (сплошная черная линия); в 0.1% DDPC и 10 мМ β-меркаптоэтанола (пунктирная линия), и после термического сканирования до 95°С в присутствии 1% SDS (точечная линия). На нижней панели показаны спектры мышиного CD20 в присутствии 0.1% DDPC (пунктирная линия), 0.1% додецилмальтозида (DDM) (точечная линия); и в 0.1% DDM с добавлением 1% SDS и β-меркаптоэтанола и после нагревания в течение 2 минут при 95°С (сплошная черная линия). Результаты выражаются в виде молярной эллиптичности.
С учетом предсказанной топологии CD20 как четырехкратно пронизывающего трансмембранного (тетратрансмембранного) белка, CD20 должен иметь содержание спиралей около 35%. Можно видеть, что C2S мутант человеческого CD20 (Фигура 6, верхняя панель), мышиного CD20 (Фигура 6, нижняя панель) имеют значительный сигнал в области спектра 222 нм, ожидавшийся для белков со значительным альфа-спиральным компонентом. Добавление восстановителя не вносит значительных изменений во вторичную структуру как мышиного CD20 (нижняя панель b, сплошная (черная) линия) так и C2S мутанта человеческого CD20 либо в присутствии DDPC (верхняя панель, пунктирная линия), либо в присутствии SDS (верхняя панель, тонкая линия). Кроме того, по-видимому, вторичная структура CD20 очень устойчива в разнообразных детергентах и при различных температурах. КД спектры C2S мутанта человеческого CD20 в присутствии SDS (верхняя панель, тонкая линия) или мышиного CD20 в присутствии SDS и восстановителя (нижняя панель, сплошная (черная) линия) после термического сканирования или кратковременного нагревания почти идентичны спектрам нативных белков (см. Фигуру 6). Результаты выражаются в виде молярной эллиптичности.
Температурное сканирование от 25°С до 95°С показывает, что человеческий CD20 теряет около 35% сигнала от спиралей при 222 нм при 95°С (данные не показаны). Хотя нет доказательства согласованного анфолдинга или согласованного рефолдинга, этот сигнал в основном возвращается, когда образец охлаждается до более низких температур, по меньшей мере, после кратковременного нагревания (Фигура 6). Небольшая разница между нагревавшимся и ненагревавшимся образцами CD20 может указывать, что некая структура утрачивается безвозвратно и количество безвозвратно денатурированного белка теоретически может увеличиваться при более длительном нагревании. Добавление восстановителя к мышиному CD20 незначительно влияет на термическую устойчивость белка в SDS (панель b, черная линия). Возможно, дисульфидная связь, расположенная вне областей CD20, которые, предположительно, должны иметь альфа-спиральную структуру, вносит очень малый вклад в общую структурную устойчивость CD20. По-видимому, изменения в структуре CD20 вследствие восстановления дисульфидной связи просто не поддаются наблюдению с помощью КД. Ранее отмечалось, что восстановление дисульфидной связи β2-адренергического рецептора также оказывает ограниченное влияние на вторичную структуру, определяемую методом КД (Lin et al., 1996, Biochemistry, 35: 14445-1444(5?)1). Аналогично, на вторичную структуру диацилглицеринкиназы не влияет SDS в условиях, сходных с условиями, применяемыми в данном описании (Lau et al., 1997, Biochemistry, 36:5884-5892).
Пример 2
Экспрессия белков MS4A при использовании phoA промотора
Другие члены семейства MS4A тетратрансмембранных полипептидов экспрессируют в Е.coli под контролеме phoA промотора по методу, описанному выше в Примере 1. Генные продукты MS4A4A (SEQ ID NO: 10), MS4A6A (SEQ ID NO: 12) и MS4A7 (SEQ ID NO: 14) экспрессируют в E.coli в соответствии с методами, описанными выше в Примере 1. При использовании клеток получают высокий выход белка, как определяют с помощью иммуноблотов с антителами к His. Как показано на Фигуре 8, детектируемые MS4A полипептиды имеют молекулярную массу, близкую молекулярным массам, установленным Liang and Tedder (2001, Genomics 72: 119-127):
Пример 3
Мутация промотора phoA
E.coli промотор, phoA, выбирают для применения в экспрессирующих сложных трансмембранных белках, включая MS4A семейства тетратрансмембранных белков, таких как CD20. Как показано ниже в Таблице 5, промотор phoA включает типичные элементы промотора, такие как сайт инициации трансляции

Обычно экспрессия многопроходных трансмембранных белков млекопитающих в E.coli рассматривается как иммунизирующая (заражающая) (см., например, Grisshammer, R., 1998, В: Identification and Expression of G Protein-Coupled Receptors, pp 133-150, Ed. K.R.Lynch, Wiley-Liss Inc.; Laage et al., 2001, Traffic 1:99-104). Некоторые из этих белков очень токсичны для бактериальных клеток даже при экспрессии с очень низкими уровнями, что еще более усложняет их получение и выделение. Простая трансформация конструкций экспрессии этих белков в подходящие экспрессирующие штаммы часто дает в результате колонии аномально малого или переменного размера, сообщая токсичность клетке, даже когда промотор "выключен". Базальный уровень экспрессии этих белков, вследствие низких уровней транскрипции, также может привести к слабому бактериальному росту на богатых средах, что приводит в результате к бедной физиологии клетки до полного включения промотора. Конечным результатом часто является экспрессия, меняющаяся от одного эксперимента к другому, а также низкие выходы. Как показано далее, продуцирование таких белков в клетке-хозяине требует строго контролируемого промотора для ограничения избыточной базальной экспрессии белка в клетках.
Для предупреждения транскрипции на базальном уровне создают два новых регулируемых промотора при использовании основной последовательности и регуляторных элементов E.coli phoA промотора (Kikuchi, el al., 1981, Nucl. Acids Res. 9: 5671-5678). Промотор phoA дикого типа обычно регулируется с помощью связывания белка phoB в фосфорилированной форме в pho бокс непосредственно "апстрим" (3′-5′) от -10 или Прибнов-бокса. Связывание заставляет РНК-полимеразу также связываться и начинать процесс транскрипции при использовании этого промотора. В свою очередь PhoB фосфорилируется в цитоплазматической мембране в ответ на низкие концентрации фосфата в периплазме и в средах (Wanner, 1996, Escherichia coli and Salmonella, Neidhardt (ed.), p.1357-1381). Связывание фосфорилированного phoB с промотором в ответ на низкие уровни фосфата в средах представляет собой позитивную регуляцию phoA промотора. В отсутствие фосфорилированного phoB РНК полимераза может все еще слабо связываться с промотором phoA, используя для контакта вместо этого последовательность -10 и какую-либо -35 последовательность, что приводит к низким базальным уровням транскрипции.
Для ингибирования связывания РНК полимеразы с phoA промотором в отсутствие фосфорилированного phoB негативный контрольный элемент, в данном случае последовательность lac оператора AATTGTGAGCGGATAACAA (SEQ ID NO: 18) (Gilbert et al., 1973, Proc. Natl. Acad. Sci. USA 70: 3581-3584), встраивают "апстрим" (3′-5′) от +1 положения начала транскрипции, как показано в Таблице 5. Это добавление уменьшает транскрипцию с помощью phoA промотора, когда фосфат присутствует (или отсутствует) в средах в значительных концентрациях (см. Фигуру 9).
Вновь созданный промотор, phac, осуществляет как позитивную, так и негативную регуляцию через позитивный контрольный элемент (phoB связывающий pho бокс) и через добавленный негативный контрольный элемент, lac оператор. Промотор phac должен индуцироваться путем истощения фосфата, а также освобождаться от контроля lac репрессора при добавлении индуцирующих факторов, таких как изопропил β-D-тиогалактозид (IPTG). Сообщалось о добавлении контрольной последовательности lac оператора к другим промоторам (De Boer et al. 1983, Proc. Natl. Acad. Sci, USA, 80: 21-25; Yansura et al., 1984, Proc. Natl. Acad. Sci. USA, 81: 439-443), однако в этих промоторах последовательность lac оператора использовалась для замены негативных контрольных элементов, которыми трудно управлять, или добавлялись к конститутивным промоторам для обеспечения новых контрольных элементов.
Вторым возможным источником базальной транскрипции, которую надо контролировать, являются промоторы "апстрим" (3′-5′) от промотора, используемого для управления экспрессией нужного мембранного белка. Такие промоторы картированы, например, на плазмиде pBR322, чаще всего применяемой для конструирования векторов Е.coli. Промоторы "апстрим" от phoA теоретически могут вызывать базальные уровни транскрипции вследствие "считывания". Для предупреждения "апстрим" транскрипцию при прохождении через hac промотор и кодирующую последовательность какого-нибудь мембранного белка терминатор транскрипции, в данном случае терминатор транскрипции лямбда t0: AACGCTCGGTTGCCGCCGGGCGTTTTTTATT (SEQ ID NO: 17) (Scholtissek et al., 1987, Nucl. Acids Res., 15: 3185), вводят "алстрим" (3′-5′) от pho бокса в корректной ориентации внутри добавленной "апстрим" (3′-5′) последовательности: AGGCCTAACGCTCGGTTGCCGCCGGGCGTTTTTTATTGTTAACCATGGA (SEQ ID NO: 19). Новый промотор, tphac, индуцируется таким же образом, что промотор phac, например, разведением в фосфат-лимитирующих средах путем добавления IPTG (данные не показаны). Транскрипцию на базальном уровне дополнительно снижают по сравнению с phac, используя tphac.

Пример 4
phoA vs. phac для экспрессии RA1c
Для проверки влияния phoA мутантного промотора phac на экспрессию и выход сложных трансмембранных полипептидов в бактериальных клетках-хозяевах методами генной инженерии создают рекомбинантный RA1c N-концевым лидером, MKHQHQQ (SEQ ED NO: 7) для инициации достаточной трансляции, и две С-концевых метки, Flag и 6-His, вводят (методами генной инженерии) в экспрессирующие векторы и функционально связывают с phoA промотором (вектор pEfRA1c) и с phac промотором (вектор pEfRA1cr). RA1c представляет собой трансмембранный полипептид, имеющий семь трансмембранных сегментов.
Трансформация
Обе плазмиды экспрессируют человеческий RA1c (Kretschmer et al., 2001, Gene 278: 41-51). Плазмиды трансформируют в Е.coli штамм 58F3 (W3110-fhuAΔ(tonAΔ) phoAΔЕ15 lonΔ galE rpoHts(htpRts) ΔclpP lacIq ΔompTΔ(nmpc-fepE) ΔslyD) и выбранные трансформанты инокулируют в 5 мл среды Луриа-Бертани (Luria-Bertani), дополненной карбенициллином (50 мкг/мл) и выращивают при 30°С на культуре (culture wheel) в течение 14-16 часов. OD600 культуры с pEfRA1c (промотор phoA) равна 1.55, а культуры с pEfRA1cr (промотор phac) 3.57. Отбирают 1 мл OD600 образца из LB инокулята.
Индукцию осуществляют встряхиванием колб. Затем каждую культуру разводят (1: 100) C.R.A.P. фосфат-лимитирующими средами (Simmons et al., 2002, J. Immunol. Methods 263: 133-147) с 50 мкг/мл карбенициллина. Все культуры выращивают при 30°С в двухлитровой колбе с разделительной перегородкой при конечном объеме индукции 500 мл на шейкере со скоростью 200 об/мин. OD600 образец 1 мл отбирают через 10, 12, 14, 16, 18, 22 и 24 часа после инокуляции в среды C.R.A.P. для pEfRA1c культуры (промотор phoA). Для культур, экспрессирующих pEfRA1cr (промотор phac), 1 мМ IPTG добавляют в среды C.R.A.P. через 10 и 12 часов после инокуляции для прекращения контроля lac репрессора. Соответствующие значения OD600 культуры при индукции IPTG равны 2.79 и 2.97. Образец OD600 (1 мл) отбирают перед добавлением IPTG и каждые два часа после добавления IPTG вплоть до 24 часов для всех культур, индуцированных IPTG.
Для анализа методом Вестерн-блоттинга восстановленные лизаты цельных клеток 1 мл OD600 образцов готовят следующим образом:
(1) 1 мл OD600 образцов центрифугируют в центрифужной пробирке.
(2) Каждый клеточный осадок ресуспендируют в 100 мкл ТЕ (10 мМ Tris pH 7.6, 1 мМ EDTA).
(3) Для восстановления дисульфидных связей в каждый образец добавляют по 10 мкл 1 М дитиотреитола (Sigma D-5545).
(4) К каждому образцу добавляют 20 мкл 20% SDS.
Образцы встряхивают, нагревают при 90°С в течение 5 минут, а затем снова встряхивают. По охлаждении образцов до комнатной температуры к осажденному белку добавляют 800 мкл ацетона. Образцы встряхивают при комнатной температуре в течение 15 минут. После центрифугирования в течение 5 минут в микроцентрифуге супернатант каждого образца отсасывают и каждый клеточный осадок ресуспендируют в 10 мкл 1 М дитиотреитола плюс 40 мкл dH2O и 50 мкл буфера для образцов 2Х NOVEX SDS. Затем образцы нагревают в течение 5 минут при 90°С, хорошо встряхивают и оставляют охлаждаться до комнатной температуры. Затем центрифугируют последний раз в течение 5 минут и супернатанты переносят в чистые пробирки.
После приготовления SDS образца образцы инокулята, 8 мкл "прединдукционных" образцов, наносят на 10 лунок геля 1.0 мм NOVEX16% Трис-Глицин SDS-PAGE и подвергают электрофорезу при примерном напряжении 120 вольт в течение, примерно, 1.5 часов. Образцы, индуцированные в C.R.A.P. среды (pEfRA1c), готовят таким же образом. 15.3 мкл образцов, индуцированных как в средах C.R.A.P., так и с помощью IPTG (образцы pEfRA1cr), наносят на 15 лунок геля. Полученные гели используют затем для Вестерн-блот анализа.
Вестерн-блот анализ
Гели SDS-PAGE подвергают электроблоттингу на нитроцеллюлозные мембраны (NOVEX) в 20 мМ натрий-фосфатного буфера, рН 6.5. Затем мембраны блокируют, используя раствор 1X NET (150 мМ NaCl, 5 мМ EDTA, 50 мМ Tris, рН 7.4, 0.05% Тритон Х-100)+0.5% желатина, примерно, от 30 минут до одного часа на шейкере при комнатной температуре. После стадии блокирования мембраны помещают в раствор 1X NET + 0.5% желатина + антитело к His (конъюгированное с пероксидазой моноклональное антитело, специфичное к his6, от Roche Diagnostics) для анти-His Вестерн-блот анализа. Разведение антитела против His составляет 1:5000, и мембраны оставляют в растворе антитела при встряхивании на качалке при комнатной температуре в течение ночи. На следующее утро мембраны отмывают минимум 3×10 минут в IX NET, а затем 1×15 минут в TBS (20 мМ Tris рН 7.5, 500 мМ NaCl). Полосы белка, связанного антителом против His, визуализуют, используя метод Amersham Pharmacia Biotech ECL и экспонируя мембраны на рентгеновскую пленку.
Действие phac промотора видно уже на стадии трансформации плазмиды. Трансформацию при использовании pEfRA1c (phoA) и pEfRA1cr (phac) сравнивают после выращивания в течение ночи на LB планшетах. Клетки, трансформированные с помощью pEfRA1cr (phac), значительно больше, чем клетки, трансформированные с помощью pEfRA1c (данные не показаны). На Фигуре 9 показан Вестерн-блот анализ с антителом против His образцов колоний, взятых (полоски) с планшетов и инокулированных в LB/Carb, примерно, на 14-16 часов. Как показано на средней дорожке блота, экспрессия RA1c при использовании неиндуцированного phoA промотора (pEfRA1c) наводит на мысль об утечке (рассеивании) при использовании phoA промотора, когда фосфат не лимитирован. Напротив, никакой обнаруживаемой RA1c экспрессии не наблюдается при использовании строго контролируемого phac промотора (pEfRA1cr).
Экспрессия на базальном уровне многопроходного трансмембранного белка RA1c под контролем phoA промотора токсична для клетки, вызывает слабый рост в LB и приводит в результате к общей низкой плотности клеток. OD600 считывание инокулята в LB после стояния в течение ночи показывает увеличение с 1.55 в pEfRA1c до 3.57 в pEfRA1cr, что указывает на более жизнеспособные клетки с промотором phoA.
Результаты цикла экспрессии pEfRA1c (phoA промотор) показаны на Фигуре 10. Полагают, что фосфат в культуре истощается, примерно, до 2.0 OD600. Экспрессию детектируют уже в 10-м часу, показатель OD600 составляет 1.73. Очень узкий интервал продуцирования, около двух часов, наблюдается с 12го до 14го после индукции, но после этого продуцирование медленно "исчезает". К 24му часу после индукции полоса мономера экспрессируемого белка полностью исчезает, кроме того, так как "мономерная" полоса исчезает со временем, в пятне аккумулируется невосстанавливаемый агрегированный белок. Наивысшая плотность клеток при сохранении наибольшего количества белка в "мономерной" полосе составляет 2.51 OD600 через 14 часов (* на Фигуре 10).
Оптимально phac промотор должен быть полностью выключен до момента индукции истощением фосфата в средах C.R.A.P. Цикл экспрессии RA1c в культурах с индуцируемым IPTG промотором phac (pEfRA1cr) показан на Фигуре 11. Максимальное продуцирование белка достигается в течение двух часов после добавления IPTG и наблюдается аналогичный паттерн экспрессии мономерной полосы и пятна. Культура, индуцированная через 12 часов, демонстрирует лучшую экспрессию, нежели индуцированная через 10 часов, что указывает на большое значение времени индукции. Более важно, что phac промотор сообщает преимущество сохранения более жизнеспособных клеток, так что они могут расти до более высокой плотности, при этом промотор остается полностью выключенным до нужного момента. При индукции клетки претерпевают стресс и прекращают рост и, в конечном счете, плотность клеток падает. Культура, выращиваемая в течение 12 часов ("12-часовая культура") до момента добавления IPTG, уже начала экспрессировать RA1c, предположительно, вследствие частичной индукции промотора phac в результате истощения фосфата. Если бы IPTG нужно было добавлять несколько позже 12 часов, промотор phac мог бы оказаться менее полезным. Следовательно, в условиях таких культур добавление IPTG через 12 часов является самым поздним временем, когда можно воспользоваться преимуществами промотора phac. Максимальное продуцирование белка достигается через два часа после индукции с помощью IPTG (через 12 часов), через 14 часов. OD600 через 14 часов равна 2.97 при использовании промотора phac, что выше 2.51, значения, полученного с промотором phoA.
Сравнение общей экспрессии RA1c при использовании промотора phoA и промотора phac показано на Фигуре 12. Наилучшие результаты экспрессии при использовании промоторов phoA и phac представлены на том же блоте с целью непосредственного сравнения. Ясно, двухчасовая индукция RA1c с промотором phac после 12 часов роста (правая дорожка) дает в результате более высокую экспрессию белка и более высокую плотность клеток, чем наивысшая экспрессия при использовании phoA промотора через 14 часов (левая дорожка).
Пример 5
phoA vs. phac Экспрессия рецептора 73, связанного с человеческим G белком (GPR73)
Для последующего анализа влияния phoA мутантного промотора phac на экспрессию и выход EG-VEGF рецептора (GPR73) осуществляют индукцию с помощью встряхивания колбы при использовании плазмид pST239.EGVEGFr1. Flag.H8.1270 (phoA промотор) и pR1FHr (phac промотор). Обе плазмиды экспрессируют белок рецептор 73, связанный с человеческим белком G (hGPR73) (Lin, et al., 2002, J. Biol. Chem. 277: 19276-19280) с N-концевой лидерной последовательностью (MKHQHQQ. SEQ ID NO: 7) для эффективной инициации трансляции и двумя С-концевыми метками, Flag и окта-His. Плазмиды трансформируют в Е.coli штамм 58F3 (W3110-fhuAΔ(tonAΔ)phoAΔE15 lonΔ galE rpoHts (htpRts) ΔclpP lacIq ΔompTΔ (nmpc-fepE) ΔslyD). Выбранные трансформанты инокулируют в 5 мл среды Луриа-Бертани (Luria-Bertani), дополненной карбенициллином (50 мкг/мл) и выращивают при 30°С на культуре (culture wheel) в течение 14-16 часов. Оптическую плотность (OD600) культуры после выращивания в течение ночи с pST239.EGVEGFr1.Flag.H8.1270, содержащей промотор phoA, равна 0.84, а культуры с pR1FHr, содержащей промотор phac, равна 3.19.
Отбирают образец, 1 мл OD600 стоявшего в течение ночи ("ночного") LB инокулята. Затем каждую культуру разводят (1:100) в C.R.A.P. фосфат-лимитирующих средах (Simmons et al., 2002, J. Immunol. Methods, 263: 133-147) с 50 мкг/мл карбенициллина. Все культуры выращивают при 25°С в двухлитровой колбе с разделительной перегородкой при конечном объеме индукции 500 мл на шейкере со скоростью 200 об/мин. OD600 образец 1 мл отбирают через 14, 15, 16, 17 и 24 часа после инокуляции в среды C.R.A.P. для pST239.EGVEGFr1 .FlagHS. 1270 (phoA) культуры. Для культур, экспрессирующих pR1FHr (phac), 1 мМ IPTG добавляют в среды C.R.A.P. через 14 и 15 часов после инокуляции. Соответствующие значения OD600 культур при индукции IPTG равны 2.37 (14 часов) и 3.21 (15 часов).
Образец 1 мл OD600 отбирают перед добавлением IPTG, через два, три и десять часов после добавления IPTG для индукции в течение 14 часов. Для индукции в течение 15 часов образец 1 мл OD600 отбирают перед добавлением IPTG, через два и через девять часов после добавления IPTG.
Восстановленные клеточные лизаты образцов по 1 мл OD600 готовят, как описано выше для RA1c в Примере 4. После приготовления SDS образца, 8 мкл восстановленных лизатов цельных клеток наносят 10 лунок геля 1.0 мм 16% Трис-Глицин SDS-PAGE фирмы NOVEX и подвергают электрофорезу при примерном напряжении 120 вольт в течение, примерно, 1.5 часов. Полученные гели используют затем для Вестерн-блот анализа, как описано для RA1 в Примере 3.
Результаты Вестерн-блот анализа экспрессии GPR73 при использовании промотора phoA и промотора phac с антителом против His показаны на Фигуре 13. Человеческий GPR73 экспрессирует при использовании промотора phoA (pST239. EGVEGFr1.Flag.H8.1270) перед индукцией (средняя дорожка), что указывает на утечку промотора phoA, когда фосфат не является лимитирующим. Напротив, никакой экспрессии hGPR73 не детектируется с строго контролируемым промотором phac (правая дорожка). Как отмечается для экспрессии RA1c в Примере 3, экспрессия на базальном уровне многопроходного трансмембранного белка с промотором phoA токсична для клетки, что вызывает слабый рост в LB и приводит к общей низкой плотности клеток. Результат OD600 Дяя "ночного" LB инокулята повышается от 0.84 в pST239. EGVEGFr1.Flag.H8.1270 (phoA промотор) до 3.19 в pR1 FHr (phac промотор), что указывает на более жизнеспособные клетки и более здоровый рост клеток, содержащих phac промотор.
Результаты цикла экспрессии GPR73 при использовании промотора phoA показаны на Фигуре 14. Экспрессию определяют по полосе димера через 15 часов с показателем OD600 3.14, а белок полностью исчезает через 24 часа. Напротив, клетки продолжают расти, что видно по повышению величины OD600 reading. Этот непрекращающийся рост является примером изменчивых результатов, наблюдаемых при экспрессии некоторых многопроходных трансмембранных белков при использовании промотора phoA, и наводит на мысль, что многие клетки какими-либо методами подавляют (модулируют по типу отрицательной связи) экспрессию. Вне зависимости от этого паттерн экспрессии GPR73 аналогичен паттерну экспрессии в Примере 3 для RA1c или для других многопроходных трансмембранных белков. Экспрессия достигает пика через 16 часов и соответствующее значение OD600 равно 3.46.
Оптимально промотор phac должен полностью выключиться до момента индукции истощением фосфата в средах C.R.A.P. Индуцированная с помощью IPTG экспрессия GPR73 показана на Фигуре 15. Никакой детектируемой экспрессии hGPR73 не наблюдается в отсутствие индукции с помощью IPTG, что говорит о строгом контроле промотора phac. Максимальное продуцирование белка достигается в течение двух часов после добавления IPTG. Аналогичный паттерн экспрессии наблюдается, если смотреть по рано появляющимся мономерной и димерной полосам (полосам мономера и димера), тогда как пятно появляется поздно при индукции. При индукции клетки претерпевают стресс и перестают расти, и в конце концов плотность клеток падает. Культура, индуцированная через 15 часов, обладает лучшей (более высокой) экспрессией, чем культура, индуцированная через 14 часов. Максимального продуцирования достигают через два часа после индукции с помощью IPTG (через 15 часов) в час 17, и величина OD600 равна 2.41.
Сравнение общей экспрессии hGPR73 при использовании промоторов phoA и phac дано на Фигуре 16. Наилучшие результаты для каждого промотора представлены на одном и том же блоте для непосредственного сравнения. Никакой детектируемой экспрессии hGPR73 не наблюдается с применением промотора phoA (средняя дорожка) по сравнению с промотором phac (правая дорожка), за исключением случая с более продолжительной экспозиции с пленкой. Ясно, что двухчасовая индукция hGPR73 промотором phac через 15 часов дает значительно более высокую экспрессию белка. Результаты экспрессии с применением промотора phoA также имеют тенденцию меняться, тогда как результаты экспрессии с применением phac промотора сравнительно постоянны.
Пример 6
phoA vs. tphac для мембранного белка MS4A4A
Для анализа влияния мутантного промотора phoA, tphac, на экспрессию и выход полипептидов семейства MS4A осуществляют индукцию встряхиванием колбы с применением плазмид pMS4A4A.8His.32 с помощью промотора phoA и pMS4A4ArT с помощью промотора tphac. Обе плазмиды экспрессируют рецептор человеческого иммуноглобулина Е-подобный белок, (h1GERB) (hMS4A4A) (Liang, et al., 2001, Genomics 72: 119-127) с N-концевым лидером, MKHQHQQ, для эффективной инициации трансляции, и С-концевой 8x-his меткой. Плазмиды трансформируют в Е.coli штамм 58F3 (W3110-fhuAΔ (tonAΔ) phoAΔE15 lonΔ ga1E rpoHts(htpRts) ΔclpP laclq ΔompTΔ(nmpc-fepE) ΔslyD) и выбранные трансформанты инокулируют в 5 мл среды Луриа-Бертани, дополненной карбенициллином (50 мкг/мл) и выращивают при 30°С на культуре (culture wheel) в течение 14-16 часов. Величина OD600 культуры с pMS4A4A.8His.32 (phoA) равна 2.4, а культура с pMS4A4ArT (tphac) равна 2.5. Отбирают 1 мл OD600 образца из LB инокулята.
Каждую культуру разводят (1:100) C.R.A.P. фосфат-лимитирующими средами (Simmons et al., 2002, J. Immunol. Methods 263: 133-147) с 50 мкг/мл карбенициллина. Все культуры выращивают при 30°С в двухлитровой колбе с разделительной перегородкой при конечном объеме индукции 500 мл на шейкере со скоростью около 200 об/мин. OD600 образец 1 мл отбирают через 10, 11, 12, 14 и 15 часов после инокуляции в среды C.R.A.P. для pMS4A4A.8His.32 культуры (промотор phoA). Для культур, экспрессирующих pMS4A4ArT (tphac), 1 мМ IPTG добавляют в среды C.R.A.P. через 10 и 10.5 часов после инокуляции. Соответствующие значения OD600 культуры при индукции IPTG равны 2.1 и 2.6.
Образец OD600 (1 мл) отбирают перед добавлением IPTG и через один, два, четыре и пять часов после добавления IPTG для культуры pMS4A4ArT (через 10 часов после добавления IPTG). Для культуры pMS4A4ArT, полученной в результате добавления IPTG через 10.5 часов, отбирают 1 мл OD600 образца перед добавлением IPTG, через один и четыре часа после добавления IPTG.
Восстановленные клеточные лизаты образцов по 1 мл OD600 готовят, как описано в Примере 4. После приготовления SDS образца, 8 мкл образцов pMS4A4A.8His.32 (phoA) и pMS4A4ArT (tphac) наносят на 10 лунок геля 1.0 мм 16% Трис-Глицин SDS-PAGE фирмы NOVEX и подвергают электрофорезу при примерном напряжении 120 вольт в течение, примерно, 1.5 часов. Индуцированные образцы в средах C.R.P.A. наносят на 15 лунок геля. Полученные гели используют затем для Вестерн-блот анализа, как описано в Примере 3.
Результаты Вестерн-блот анализа с антителом против His, показывающие экспрессию MS4A4A, векторы pMS4A4A.8His.32 (phoA) и pMS4A4ArT (tphac) в жидких средах LB, представлены на Фигуре 17. Рецептор человеческого иммуноглобулина Е-подобный белок, (h1GERB) экспрессируют при использовании промотора phoA (pMS4A4A.8His.32) в отсутствие индукции, что указывает на утечку промотора phoA, когда фосфат не является лимитирующим. Напротив, никакой экспрессии hMS4A4A не детектируется с строго контролируемым промотором tphac. Результат OD600 для "ночного" LB инокулята немного повышается от 2.4 в pMS4A4A.8His.32 (phoA промотор) до 2.5 (tphac промотор).
Результаты цикла экспрессии MS4A4A при использовании промотора phoA показаны на Фигуре 18. Экспрессию определяют через 11 часов с помощью показателя OD600 2.24. Белок является относительно устойчивым во времени; однако экспрессия не увеличивается во времени. Максимальное продуцирование достигается в течение одного часа после 11го часа. С этого момента падает плотность клеток.
Результаты индуцированной IPTG экспрессии MS4A4A при использовании pMS4A4ArT (tphac) культур показаны на Фигуре 19. Никакой детектируемой экспрессии hIGERB без добавления IPTG не наблюдается, что наводит на мысль о строгости промотора tphac. Экспрессия обнаруживается в течение часа после индукции с помощью IPTG. Культура, индуцированная через 10 часов, с OD600 2.1 проявляет слегка повышенную экспрессию по сравнению и культурой, индуцированной через 10.5 часов, с OD600 2.6. Максимального продуцирования достигают через 2 часа после индукции с помощью IPTG, всего через 12 часов (IPTG добавляют через 10 часов), и OD600 равна 2.36.
Сравнение общей экспрессии hMS4A4A сравнивают для промотора phoA и для промотора tphac, дано на Фигуре 20. Наилучшие данные для экспрессии при использовании промотора phoA и промотора tphac представлены на том же блоте для непосредственного сравнения. "12-Часовой" образец культуры pMS4A4ArT, индуцированной с помощью IPTG (IPTG добавляют через 10 часов), показывает значительное увеличение экспрессии по сравнению с "11-часовым" образцом pMS4A4A.8His.32 с промотором phoA. Величина OD600 культуры, контролируемой tphac, равна 2.19, тогда как OD600 в случае максимальной экспрессии с помощью phoA промотора равна 2.24.
Пример 7
Повышение экспрессии CD20 при использовании лидеров trpLE
Последовательность trp LE уже много лет используют в качестве партнера слияния по N-концу для экспрессии особенно проблематичных белков, которые плохо аккумулируются (см., например, Yansura, 1990, Methods in Enzymology, Vol.185: 161-166, Academic Press Inc., San Diego, CA). Белки trpLE обычно представляют собой делеции внутри рамки считывания trp лидера на одном конце и дистальные участки trp Е гена. Сообщалось о двух вариантах, ΔtrpLE1417 и ΔtrpLE14l3 (см., например, Bertrand et al., 1976, J. Mol. Biol. 103: 319-337 и Miozzari et al., 1978, J. Bacterial. 133: 1457-1466). ΔtrpLE1413 используют для конструкции векторов экспрессии некоторых человеческих белков (см., например, патент США 5888808). Однако, как сообщалось, попытки экспрессировать трансмембранный белок, поверхностный антиген вируса гепатита В при использовании аналогичного вектора, pHS94, дали отрицательные результаты (см., например, патенты США 4803164 и 4741901).
Высокоуровневую экспрессию белков, слитых с trpLE, обычно относят за счет сильного trp промотора, сильной области инициации трансляции, которая включает первые несколько кодонов trp лидера и результирующего "проталкивания" гетерологичных белков в протеолитически устойчивые преломляющие тельца. Для определения, действительно ли trpLE лидер может направлять многопроходные трансмембранные белки в преломляющие тельца и/или иным способом повышать экспрессию и выход гетерологичных многопроходных трансмембранных белков в Е.coli, созданы несколько конструкций с использованием нескольких вариантов лидерной последовательности trpLE, обозначенной LE (SEQ ID NO: 25), и sLE (SEQ ID NO: 26). Эти конструкции создают с более строго регулируемыми промоторами phoA и phac/tphac, описанными в вышеприведенных Примерах.
Более протяженный из двух лидеров, LE, содержит всего лишь первые девять аминокислот ((M)KAIFVLKGS, SEQ ID NO: 27) Е.coli белка trp Е, описанного, например, в экспрессирующем векторе pNCV (ΔtrpLE1413) (Maniatis et al., В Molecular Cloning: A Laboratory Manual, p 426, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1982), с последующими аминокислотами 339-408 полипептида trpE (SEQ ID NO: 25), показанного на Фигуре 22. Лидер с меньшей trpLE последовательностью (sLE), содержащий те же первые девять аминокислот лидера trp, связанных с последовательностью непрерывных аминокислот полипептида trpЈ (SEQ ID NO: 26), также показан на Фигуре 22. Эти два trpLE лидера ведут себя аналогично с точки зрения проникновения в мембрану, хотя уровень экспрессии, примерно, в два раза выше в случае более протяженного LE лидера.
Клонирование и экспрессия LE CD20
Мутантный CD20, в котором Cys111 и Cys220, каждый, заменен на серин, a Cys81 заменен на аланин, субклонируют стандартными методами молекулярной биологии (Ausubel et al., (eds.), 2003, Current Protocols in Molecular Biology, 4 Vols., John Wiley & Sons) в образованную из pBR322 плазмиду, содержащую ген бета-лактамазы и гены tPHK трех редких кодонов Е.coli (argU, glyT и pro2). На N-конце добавляют trpLE спейсерную последовательность из 79 аминокислот (SEQ ID NO: 25) и GS линкерную последовательность. Сайт распознавания и расщепления тромбина добавляют после девяти аминокислот CD20, для отщепления trpLE лидера, и после 236ой аминокислоты CD20, для отщепления внутриклеточного гидрофильного хвоста. Для кодирования последовательности метки на С-конце добавляют окта-His (SEQ ID NO: 8), чтобы содействовать обнаружению и очистке экспрессированного белка. Полученная плазмида обозначена pLErGKiSArT. Транскрипцию гена осуществляют под контролем phoA промотора, а экспрессию индуцируют, лимитируя фосфат в Е.coli штамме 58F3, как описано выше в Примере 4. Насыщенную LB карбенициллином культуру разводят в C.R.A.P. фосфат-лимитирующих средах (Simmons et al., 2002, J. Immunol. Methods, 263:133-147). Затем культуры выращивают при 30°С в течение 24 часов.
Для анализа экспрессии готовят 1 мл OD600 образцов восстановленных лизатов цельных клеток следующим образом:
(5) образцы 1 мл OD600 центрифугируют в микроцентрифужной пробирке.
(6) каждый клеточный осадок ресуспендируют в 70 мкл ТЕ (10 мМ Tris pH 7.6, 1 мМ EDTA).
(7) Для восстановления дисульфидных связей к каждому образцу добавляют 10 мкл 1 М дитиотреитола (Sigma D-5545).
(8) В каждый образец добавляют 20 мкл 20% SDS.
Образцы встряхивают, нагревают при 90°С в течение 5 минут, а затем снова встряхивают. По охлаждении образцов до комнатной температуры прибавляют 100 мкл 2Х NOVEX SDS. Затем образцы нагревают в течение 5 минут при 90°С, хорошо встряхивают и оставляют охлаждаться до комнатной температуры. Затем центрифугируют последний раз в течение 5 минут и супернатанты переносят в чистые пробирки.
После приготовления SDS образца образцы инокулята, 16 мкл "индукционных" образцов, наносят на 10 лунок геля 1.0 мм NOVEX 16% Трис-Глицин SDS-PAGE и подвергают электрофорезу при примерном напряжении 120 вольт в течение, примерно, 1.5 часов. Полученный гель окрашивают Кумасси синим с добавлением 10% уксусной кислоты. Для Вестерн-блот анализа наносят 1 мкл лизата цельных клеток и полученный в результате гель подвергают электроблоттингу на нитроцеллюлозную мембрану (NOVEX) в 1X буфера Трис-глицин (Invitrogen, CA), 0.01% SDS, 5% метанола. Затем мембраны блокируют в растворе 1X NET (150 мМ NaCl, 5 мМ EDTA, 50 мМ Tris (Трис), pH 7.4, 0.05% Тритон Х-100) и 0.5% желатина в течение, примерно, от 30 до одного часа на шейкере при комнатной температуре. После стадии блокирования мембраны помещают в раствор IX NET, 0.5% желатина, содержащий антитело к His (конъюгированное с пероксидазой моноклональное антитело, специфичное к His6, от Roche Diagnostics) для анти-His Вестерн-блот анализа. Разведение антитела против His составляет 1:5000, и мембраны отмывают минимум 3×10 минут в IX NET, а затем 1×15 минут в TBS (20 мМ Tris pH 7.5, 500 мМ NaCl). Полосы белка, связанного антителом против His, визуализуют, используя метод Amersham Pharmacia Biotech ECL и экспонируя мембраны на рентгеновскую пленку.
На Фигуре 23 показана конструктивная схема результатов экспрессии LE.CD20 и CD20 при использовании LE.CD20 по сравнению с экспрессией при использовании промотора PhoA без trpLE лидера, но под контролем phoA промотора с лидерной последовательностью MKHQHOQ (SEQ ID NO: 7), описанной в Примере 1. Экспрессия и продуцирование белка CD20 в клетках Е.coli при использовании trpLE значительно повышена до уровня, обнаруживаемого на геле, окрашенном Кумасси синим. Вестерн-блот анализ показывает, примерно, в десять раз более высокий выход экспрессии CD20 при использовании trpLE лидера, чем в отсутствие trpLE лидера (данные не показаны). Примерный уровень экспрессии около 30 мг CD20, слитого с trpLE, на 1 л культуры.
Экстракция мембранного белка LE CD20
Для анализа растворимости слияния trpLE с CD20 клеточный осадок ресуспендируют, используя Polytron (Brinkmann, Westbury, NY), в 10 мл нативного буфера для лизиса (20 мМ Tris pH 7.5, 300 мМ NaCl) на каждый 1 г пасты, лизируют дезинтеграцией клеток в микрофлюидизаторе (Microfluidics Corp, Newton, MA) и центрифугируют при 391000 × g в течение 1 часа на ультрацентрифужном роторе TLA-100.3 (Beckman, Fullerton, CA). Супернатант, содержащий растворимые белки (Sup 1), отделяют от осадка, содержащего мембранные белки и нерастворимые белки (осадок 1). Осадок 1 затем ресуспендируют, используя Polytron в нативном буфере для лизиса с неденатурирующим нейтральным детергентом, 1% Fos-Choline 12, и экстрагируют в течение ночи при 4°С. На следующий день образец снова центрифугируют на ультрацентрифуге и супернатант, содержащий мембраносвязанные белки в мицеллярной форме (Sup 2), отделяют от нерастворимых белков (осадок 2). Клеточные осадки и супернатанты ресуспендируют в восстанавливающем SDS лодинг-буфере и анализируют с помощью SDS-PAGE, а иммуноблот на нитроцеллюлозной мембране анализируют с помощью антител к His, конъюгированных с пероксидазой хрена (Roche Applied Science, Indianapolis, IN).
Результаты демонстрируют почти полную экстракцию trpUE.CD20 из мембран, что видно на окрашенном Кумасси синим геле и на иммуноблоте (Фигура 24).
Центрифугирование LE CD20 в градиенте плотности
Непрерывный градиент плотности сахарозы получают, послойно помещая в центрифужные пробирки 1.9 М и 1.4 М растворы сахарозы, забуференные 150 мМ NaCl и 20 мМ HEPES, рН 7.2. Клетки, экспрессирующие белок LE CD20, лизируют в нативном буфере для лизиса. Фракцию мембранных и растворимых белков выделяют ультрацентрифугированием при 391000 × g (85K об/мин) в течение 1 часа на ультрацентрифужном роторе TLA100.3 (Beckman, Fullerton, СА). Супернатант отбрасывают и осадок ресуспендируют в 1.9 М растворе сахарозы. Аликвоту 100 мкл повторной суспензии (ресуспензии) снова смешивают с 0.9 мл 1.9 М раствора сахарозы. Затем эту смесь помещают на дно центрифужной пробирки и сверху помещают слой 1 мл 1.4 М раствора сахарозы. Образец загружают в ротор SWTi55 и центрифугируют в течение 1 часа при 128,356 × g (32.5K об/мин). Затем фракционированный образец осторожно выгружают в аликвотах 200 мкл сверху пробирки и каждую фракцию (10 сверху, 1 внизу) анализируют методом SDS-PAGE, переносят на нитроцеллюлозную мембрану и анализируют с помощью конъюгированного с пероксидазой хрена антитела к His. Вестерн-блот анализ, демонстрирующий слияние trp LE с CD20 полипептидом, обнаруживают в мембранном слое, расположенном между (интерфейс) растворами сахарозы 1.4 М и 1.9 М (фракции 5 и 6), причем основная часть полипептида находится во фракции 5 (данные не показаны), это указывает, что, по-видимому, основная часть слитого с trpLE полипептида CD20 включена в Е.coli мембрану.
Расщепление LE CD20 тромбином
Тромбин с 1 мМ хлорида кальция добавляют к растворимым мембранным экстрактам 1% Fos-Choline 12 из анализа на растворимость в разведении 1:1000 и оставляют при комнатной температуре в течение ночи. Восстановленные SDS приготовленные образцы анализируют методом SDS-PAGE. Гель, окрашенный Кумасси синим, демонстрирует оптимальное расщепление двух сайтов расщепления тромбином на слитом белке, что дает полосы трех белков, включая трансмембранный домен усеченного CD20 (26 кДа), trpLE (11 кДа) и гидрофильный С-концевой хвост CD20 (8 кДа) (данные не показаны). N-концы пептидов из каждой из трех полос секвенируют для подтверждения идентичности.
Пример 8
Повышение экспрессии RA1c с помощью trpLE лидера
Клонирование и экспрессия LERA1c
ДНК, кодирующую RA1c, субклонируют стандартными способами молекулярной биологии, как описано выше, в плазмиду, образованную из pBR322, содержащую ген бета-лактамазы и гены tPHK трех редких для Е.coli кодонов (argU, glyT и pro2). На N-конце RA1c добавляют trpLE лидер, а затем метку flag (DYKDDDDK, SEQ ID NO: 32) и сайт распознавания тромбином (thrX) (LVPRGS, SEQ ID NO: 31) для гарантии инициации высокого уровня трансляции, детектирования и расщепления соответственно. На С-конце добавляют десять остатков гистидина, чтобы содействовать детектированию и очистке экспрессированного белка. Полученную плазмиду обозначают pLEfRA1CnFcHrT. Транскрипция гена осуществляется под контролем tphac промотора, а экспрессию индуцируют с помощью лимитирующего фосфата и добавления 1 мМ IPTG до примерной плотности клеток 2-3 OD600, как описано в вышеприведенных Примерах. Насыщенную LB карбенициллином культуру разводят в C.R.A.P. фосфат-лимитирующих средах (Simmons et al., 2002, J. Immunol. Methods, 263: 133-147). После этого культуру выращивают при 30°С в течение 24 часов после добавления IPTG.
Для анализа экспрессии готовят 1 мл OD600 образцов восстановленных лизатов цельных клеток, как описано ранее в Пример 7. После приготовления SDS образца 5 мкл "индукционных" образцов наносят на 10 лунок геля 1.0 мм NOVEX 16% Трис-Глицин SDS-PAGE и подвергают электрофорезу при примерном напряжении 120 вольт в течение, примерно, 1.5 часов. Для Вестерн-блот анализа полученный в результате гель подвергают электроблоттингу на нитроцеллюлозную мембрану (NOVEX) в 1X буфера Трис-глицин (Invitrogen, CA), 0.01% SDS, 5% метанола. Затем мембраны блокируют в растворе 1X NET (150 мМ NaCl, 5 мМ EDTA, 50 мМ Tris (Трис), pH 7.4, 0.05% Тритон Х-100) и 0.5% желатина в течение, примерно, от 30 до одного часа на шейкере при комнатной температуре. После стадии блокирования мембраны помещают в раствор IX NET, 0.5% желатина, содержащий антитело к His (конъюгированное с пероксидазой моноклональное антитело, специфичное к His6, от Roche Diagnostics) для анти-His Вестерн-блот анализа. Разведение антитела против His составляет 1:5000, и мембраны отмывают минимум 3×10 минут в IX NET, а затем 1×15 минут в TBS (20 мМ Tris pH 7.5, 500 мМ NaCl). Полосы белка, связанного антителом против His, визуализуют, используя метод Amersham Pharmacia Biotech ECL и экспонируя мембраны на рентгеновскую пленку.
На Фигуре 25 показана конструктивная схема результатов экспрессии LE.RA1c (47.6 кДа) и дано сравнение экспрессии RA1 при использовании LE. RA1c экспрессией RA1 при использовании конструкции, не содержащей trpLE лидера, но с лидерной последовательностью MKHQHOQ (SEQ ID NO: 7), обе экспрессии под контролем промотора tphac, как описано в Примере 4. Вестерн-блот анализ с применением антитела к HIS показывает значительно более высокий уровень экспрессии в отношении LE.RA1c, содержащего LE лидерную последовательность, по сравнению с RA1c без trpLE лидера (см. Фигуру 25).
Экстракция мембранного белка RA1c
Для анализа растворимости слияния trpLE с RA1c клеточный осадок ресуспендируют, используя Polytron (Brinkmainn, Westbury, NY), в 10 мл нативного буфера для лизиса (20 мМ Tris pH 7.5, 300 мМ NaCl) на каждый 1 г пасты, лизируют дезинтеграцией клеток в микрофлюидизаторе (Microfluidics Corp, Newton, MA) и центрифугируют и при 391000×g в течение 1 часа на ультрацентрифужном роторе TLA-100.3 (Beckman, Fullerton, CA). Супернатант, содержащий растворимые белки (sup 1), отделяют от осадка, содержащего мембранные белки и нерастворимые белки (осадок 1). Осадок 1 затем ресуспендируют, используя Polytron в нативном буфере для лизиса с неденатурирующим нейтральным детергентом, 1% Fos-Choline 12, и экстрагируют в течение ночи при 4°С. На следующий день образец снова центрифугируют на ультрацентрифуге и супернатант, содержащий мембраносвязанные белки в мицеллярной форме (sup 2), отделяют от нерастворимых белков (осадок 2). Клеточные осадки и супернатанты ресуспендируют в восстанавливающем SDS лодинг-буфере до равных объемов и анализируют с помощью SDS-PAGE, а иммуноблот на нитроцеллюлозной мембране анализируют с помощью антител к His, конъюгированных с пероксидазой хрена (Roche Applied Science, Indianapolis, IN). Как показано на Фигуре 26, наблюдается почти полная экстракция LE.RA1c из мембраны, что видно на окрашенном Кумасси синим геле и на иммуноблоте. Выход белка RA1c достаточен для визуализации окрашенным Кумасси синим гелем после экстракции с помощью 1% Fos-Choline 12.
Центрифугирование LE RA1c в градиенте плотности
LE RA1c подвергают центрифугированию в градиенте плотности по методу, описанному для CD20 в Примере 7. Коротко говоря, непрерывный градиент плотности сахарозы получают, послойно помещая 1.9 М и 1.4 М растворы сахарозы, как описано в Примере 7. Аналогично описанному в Примере 7 результаты Вестерн-блот анализа показывают, что, по-видимому, основная часть LE.RAc полипептида включена в Е.coli мембрану (данные не показаны).
Выделение белка LE RA1c
Для выделения белка LE.RA1c, описанного выше, в анализе растворимости растворимый в 1% Fos-Choline 12-мебранный экстракт наносят на колонку Ni-NTA PhyTip (Phynexus, San Jose, CA), отмывают с помощью 50 мМ имидазола и 0.5% Fos-Choline 12 в нативном буфере для лизиса и элюируют с помощью 250 мМ имидазола и 0.75% Fos-Choline 12 в нативном буфере для лизиса. Фракции элюата ресуспендируют в восстанавливающем SDS лодинг-буфере и анализируют с помощью SDS-PAGE. Гель SDS PAGE показывает очищенный LE.RA1c с молекулярной массой 47.6 кДа (данные не показаны).
Известно, что связанные с G-белком рецепторы олигомеризуются, например, димеризуются, даже в присутствии восстановителя и SDS (Bouvier, 2001, Nature Reviews Neuroscience. 1: 274-286). Верхние полосы в геле, по-видимому, являются такими димерами и олигомерами RA1c, судя по молекулярным массам (данные не показаны). Секвенирование N-концевых аминокислот подтверждает, что очищенный белок является LE.RA1c.
Около 10 миллиграмм trpLE RA1c можно выделить из экстракта цельных клеток из 1 литра культуры во встряхиваемой колбе, если исходить из интенсивности белковых полос после окрашивания Кумасси синим.
Расщепление LE RA1c тромбином
Для выделения RA1c полипептида тромбин с 1 мМ хлорида кальция добавляют к элюату RA1c в разведении 1:1000 и оставляют при комнатной температуре в течение ночи для обеспечения ферментативной активности тромбина. Образцы восстанавливают, готовят и анализируют методом SDS-PAGE. Полученный в результате гель, окрашенный Кумасси синим, демонстрирует оптимальное расщепление слитого белка во встроенных сайтах расщепления тромбином на слитом белке, что дает две белковые полосы, слияние усеченного RA1c (37 кДа) и trpLE с меткой flag (10.6 кДа) (данные не показаны). Идентичность полос расщепленного полипептида подтверждают секвенированием по N-концу.
Пример 9
Повышенная экспрессия GPR 73 с trp LE лидером
Клонирование и экспрессия белка LE рецептор 73, связанный с человеческим G-белком (hGPR 73)
ДНК, кодирующую связанный с человеческим G-белком рецептор 73 (GPR 73), субклонируют стандартными методами молекулярной биологии (Ausubel et al., (eds.), 2003, Current Protocols in Molecular Biology, 4 Vols., John Wiley & Sons) в плазмиду, образованную из pBR322, содержащую ген бета-лактамазы и гены tPHK трех редких для Е. coli кодонов (argU, glyT и pro2). Как показано на Фигуре 25, на N-конце GPR73 добавляют trpLE лидер, а затем метку flag (DYKDDDDK, SEQ ID NO: 32) и сайт распознавания тромбином (ThrX) (LVPRGS, SEQ ID NO: 31) для гарантии инициации высокого уровня трансляции, детектирования и расщепления соответственно. На С-конце добавляют метку окта-His (SEQ ID NO: 8), чтобы содействовать детектированию и очистке экспрессированного белка. Полученную плазмиду обозначают pLEfR1nFcHrT. Транскрипция гена осуществляется под контролем tphac промотора, как описано выше, в Примере 8, а экспрессию индуцируют с помощью лимитирующего фосфата и добавления 1 мМ IPTG до примерной плотности клеток 2-3 OD600. Насыщенную LB карбенициллином культуру разводят в C.R.A.P. фосфат-лимитирующих средах (Simmons et al., 2002, J. Immunol. Methods, 263: 133-147). После этого культуру выращивают при 30°С в течение 6 часов после добавления IPTG.
Для анализа экспрессии готовят 1 мл OD600 образцов восстановленных лизатов цельных клеток, как описано ранее в Пример 7. SDS "индукционные" образцы 5 мкл наносят на 10 лунок геля 1.0 мм NOVEX 16% Трис-Глицин SDS-PAGE и подвергают электрофорезу при примерном напряжении 120 вольт в течение, примерно, 1.5 часов. Для Вестерн-блот анализа полученный в результате гель подвергают, электроблоттингу на нитроцеллюлозную мембрану (NOVEX) в 1X буфера для переноса (Invitrogen, СА), 0.01% SDS, 5% метанола. Затем мембраны блокируют в растворе 1X NET (150 мМ NaCl, 5 мМ EDTA, 50 мМ Tris (Трис), pH 7.4, 0.05% Тритон Х-100) и 0.5% желатина в течение, примерно, от 30 до одного часа на шейкере при комнатной температуре. После стадии блокирования мембраны помещают в раствор 1X NET, 0.5% желатина и антитело к His (конъюгированное с пероксидазой моноклональное антитело, специфичное к His6, от Roche Diagnostics) для анти-His Вестерн-блот анализа. Разведение антитела против His составляет 1:5000, и мембраны отмывают минимум 3×10 минут в IX NET, а затем 1×15 минут в TBS (20 мМ Tris pH 7.5, 500 мМ NaCl). Полосы белка, связанного антителом против His, визуализуют, используя метод Amersham Pharmacia Biotech ECL и экспонируя мембраны на рентгеновскую пленку.
На Фигуре 25 показана конструктивная схема результатов экспрессии в сравнении с экспрессией GPR73 без trpLE лидера. Вестерн-блот анализ показывает значительно более высокий уровень экспрессии и выход LE.GPR73, чем без trpLE лидера.
Экстракция мембранного белка LE рецептор 73, связанный с человеческим G-белком
Для анализа растворимости слияния trpLE с GPR73 белок экстрагируют из клеточных мембран и анализируют, как описано выше, для аналогичного анализа белков LE.CD20 и LE.RA1c в Примерах 7 и 8. Мембранные белки (осадок 1) отделяют от супернатанта, содержащего растворимые белки (sup 1). Затем клеточный осадок 1 экстрагируют с помощью Fos-Choline 12, и мембранные белки в мицеллярной форме (sup 2) отделяют от нерастворимых белков (осадок 2).
Клеточные осадки и супернатанты ресуспендируют в восстанавливающем SDS лодинг-буфере до равных объемов и анализируют с помощью SDS-PAGE, а иммуноблот на нитроцеллюлозной мембране анализируют с помощью антител к His, конъюгированных с пероксидазой хрена (Roche Applied Science, Indianapolis, IN). Наблюдается почти полная экстракция GPR73 из Е.coli клеточной мембраны, что видно на иммуноблоте (Фигура 27).
Центрифугирование в градиенте плотности белка LE рецептор 73,
связанный с человеческим G-белком
Непрерывный градиент плотности сахарозы получают, послойно помещая в центрифужные пробирки 1.9 М и 1.4 М растворы сахарозы, забуференные 150 мМ NaCl и 20 мМ HEPES, рН 7.2, и применяют для разделения Е.coli мембранной фракции, как описано выше в Примерах 7 и 8. Клетки, экспрессирующие белок LE GPR73, лизируют в нативном буфере для лизиса. Фракцию мембранных и растворимых белков выделяют ультрацентрифугированием при 391000×g в течение 1 часа на ультрацентрифужном роторе TLA100.3 (Beckman, Fullerton, CA). Супернатант отбрасывают и осадок ресуспендируют в 1.9 М растворе сахарозы. Аликвоту 100 мкл повторной суспензии (ресуспензии) снова смешивают с 0.9 мл 1.9 М раствора сахарозы. Затем эту смесь помещают на дно центрифужной пробирки и сверху помещают слой 1 мл 1.4 М раствора сахарозы. Образец загружают в ротор SW55 и центрифугируют в течение 1 часа при 12835 × g (32.5К об/мин). Затем фракционированный образец осторожно выгружают в аликвотах 200 мкл сверху пробирки и анализируют методом SDS-PAGE, переносят на нитроцеллюлозную мембрану и анализируют с помощью конъюгированного с пероксидазой хрена антитела к His. Результаты демонстрируют, что, по-видимому, основная часть слитого с trpLE белка GPR73 включена в Е.coli мембрану (данные не показаны).
Выделение белка LE рецептор 73 связанный с человеческим G-белком
Для выделения белка LE.GPR73 часть растворимого в 1% Fos-Choline 12 мембранного экстракта, описанного выше, в анализе растворимости, наносят на колонку Ni-NTA PhyTip (Phynexus, San Jose, CA), отмывают с помощью 50 мМ имидазола и 0.5% Fos-Choline 12 в нативном буфере для лизиса и элюируют с помощью 250 мМ имидазола и 0.75% Fos-Choline 12 в нативном буфере для лизиса. Образцы фракций элюата ресуспендируют в восстанавливающем SDS лодинг-буфере и анализируют с помощью SDS-PAGE. Полученный гель показывает очищенный GPR73 белок с молекулярной массой 56.5 кДа (данные не показаны). Идентичность очищенного белка подтверждена N-концевым секвенированием. Расчеты выхода белка показывают, что около 2-3 миллиграмм trpLE LE.GPR73 можно выделить из экстракта цельных клеток из 1 литра культуры во встряхиваемой колбе.
Расщепление тромбином белка LE рецептор 73, связанный с человеческим G-белком
Белок GPR73 отщепляют от слитого белка во встроенных сайтах расщепления тромбином. Тромбин с 1 мМ хлорида кальция добавляют к элюату очищенного GPR73 в разведении 1:1000 и оставляют при комнатной температуре в течение ночи для обеспечения ферментативной активности. Образец отщепленного белка восстанавливают с помощью SDS и анализируют методом SDS-PAGE. Окрашенный Кумасси синим гель демонстрирует оптимальное расщепление слитого белка во встроенных сайтах, что дает две белковые полосы, включая отщепленный GPR73 (45.9 кДа) и trpLE с меткой flag (10.6 кДа) (данные не показаны). Идентичность отщепленного белка подтверждают секвенированием по N-концу.
Пример 10
Очистка мембранного белка
Очистка человеческого CD20, меченного LE и sLE
Для выделения LE и sLE белков, продуцированных в Е. coli, как описано выше в Примерах 7, 8 и 9, мембранные фракции готовят ресуспендированием цельных клеток на приборе Polytron (Brinkmann, Westbury, NY), в 1:10 вес./об. буфере для лизиса (20 мМ Tris, pH 7.5, 300 мМ NaCl и 1 мМ EDTA). Затем клетки лизируют дезинтеграцией на микрофлюидизаторе (Microfluidics Corp., Newton, MA) и смесь центрифугируют при 12000×g в течение 1 часа. Затем клеточный осадок (Р1) ресуспендируют в буфере для лизиса без EDTA, и в присутствии или в отсутствие бета-меркаптоэтанола, на гомогенизаторе Polytron. Добавляют додецилфосфохолин (DDPC, Fos-Choline® 12) до концентрации 1% и образцы пропускают через микрофлюидизатор от одного до трех раз. Затем раствор центрифугируют при 125000×g, примерно, от 45 минут до 1 часа. Супернатант наносят на колонку Ni-NTA Superflow (Qiagen Inc. Valencia, CA), предварительно уравновешенную в буфере (20 мМ Tris, pH 7.5, 250-300 мМ NaCl и 5 мМ DDPC или 0.1% н-додецил-N,N-диметиламин-N-оксида (LDAO)). Колонку отмывают 10 CV (объемами колонки) того же буфера с 50 мМ имидазола и элюируют тем же буфером с 250-300 мМ имидазола. Все стадии очистки вплоть до загрузки колонки осуществляют при 4°С.
Белок, очищенный в присутствии восстановителя, выделяют из мембран только в виде мономера (данные не показаны). Белок, выделенный в отсутствие восстановителя, присутствует как в виде мономера, так и в виде связанной дисульфидной связью димерной формы.
Для превращения димерного белка в мономерную форму CD20 восстанавливают, добавляя бета-меркаптоэтанол или DTT. Восстановитель либо удаляют с помощью нескольких циклов диализа против буфера, содержащего 20 мМ Tris pH 8.0, 300 мМ NaCl, и детергент присутствует без восстановителя, либо восстановитель нейтрализуют, добавляя окисленный восстановитель, такой как окисленный DTT или окисленный глутатион.
Метку His и лидер LE или sLE отщепляют от белка инкубацией с бычьим тромбином. Для отделения мономерного CD20 от некоторого любого остающегося димера (LE или sLE) и His-меток отщепленный белок наносят на колонку Superdex 200 (Amersham Biosciences, Piscataway, NJ), предварительно уравновешенную в 20 мМ Tris, pH 7.5, 300 мМ NaCl и 5 мМ DDPC или 0.1% н-додецил-N,N-диметиламин-N-оксидом (LDAO). Фракции, полученные на колонке для эксклюзионной хроматографии по размеру, собирают и анализируют гель-хроматографией для демонстрации локализации очищенного белка. Очищенный белок используют в описанных выше анализах связывания.
Анализ активности методом ELISA
Функциональную целостность белков CD20 оценивают способностью человеческих CD20 полипептидов, содержащих или не содержащих LE и sLE последовательности, связывать антитело ритуксимаб. Ритуксимаб распознает только скрученную конформацию человеческого CD20, в которой образуется нативная дисульфидная связь между цистеиновыми остатками 167 и 183. Для анализа нативного рефолдинга CD20 используют анализ связывания человеческого CD20 с ритуксимабом методом ELISA.
Очищенный белок CD20 анализируют методом ELISA. 96-луночные планшеты покрывают в течение ночи при 4°С, используя 100 мкл CD20 при концентрации 1 мкг/мл в PBS, содержащем солюбилизирующий детергент, разведенный до концентрации мицелл ниже критической. Затем планшеты трижды отмывают PBS, содержащим 0.05% Твин-20 (PBST), и в течение 45 при комнатной температуре блокируют с помощью 200 мкл PBST, содержащего 0.5% BSA (блокирующий и аналитический буфер). Планшеты снова трижды отмывают PBST, а затем анализируют с применением первичного антитела. Объем 150 мкл ритуксимаба при концентрации 60 мкг/мл в буфере для анализа добавляют в соответствующие лунки и делают трехкратные серийные разведения в последующих лунках, отбирая 50 мкл из первой лунки и смешивая со 100 мкл аналитического буфера в следующей и в дальнейших лунках до конечной концентрации около 2 нг/мл. После инкубации при комнатной температуре в течение 90 минут планшеты отмывают PBST и связанный ритуксимаб обнаруживают, добавляя 100 мкл конъюгата пероксидазы хрена с антителом козы к человеческому F(ab′)2 (Jackson ImmunoResearch Laboratories Inc, West Grove, PA), разведенного 1:2000 в аналитическом буфере, отмывают шесть раз с помощью PBST и проявляют, добавляя 100 мкл/лунка системы субстрата ТМВ Microwell Peroxidase Substrate System (KPL, Gaithersburg, MD), смешанной в соответствии с инструкциями производителя. Реакцию останавливают, добавляя 100 мкл/лунка 1.0 М фосфорную кислоту (100 мкл/лунка), и измеряют оптическую плотность при 450 нм на планшет-ридере.
На Фигуре 28 показаны результаты анализа связывания, в котором сравниваются ответы связывания ритуксимаба с человеческим белком CD20, экспрессированным без LE или sLE последовательности, и с человеческим белком CD20, экспрессированным с LE или sLE лидером и выделенным в различных условиях. Каждый из экспрессированных человеческих LE.CD20 и sLE.CD20 полипептидов, аналогично контрольному человеческому CD20, демонстрирует связывание со специфическим к конформации антителом, ритуксимабом.
Реферат
Изобретение относится к области биотехнологии и может быть использовано в биохимии, протеомике и фармакологии для экспрессии и получения сложных трансмембранных белков из бактериальных штаммов-продуцентов. Предложена экспрессирующая конструкция для экспрессии одно- или многопроходных трансмембранных полипептидов в бактериальной клетке-хозяине. Данная конструкция содержит белок-кодирующий полинуклеотид, строго регулируемый промотор, имеющий низкую базальную активность в клетке-хозяине, и лидерную последовательность, содержащую энхансер инициации трансляции. Строго регулируемый промотор содержит, по меньшей мере, один позитивный регуляторный элемент контроля и, по меньшей мере, один негативный регуляторный элемент контроля, причем один или более из позитивных и негативных регуляторных элементов контроля представляет собой гетерологичный регуляторный элемент контроля. Кроме того, предлагается способ получения экспрессированного трансмембранного полипептида и способ его выделения из клетки-хозяина. Предложенные способы обеспечивают получение трансмембранных полипептидов с высокими выходами, с нативной конформацией и/или в растворимой форме. 7 н. и 46 з.п. ф-лы, 28 ил., 6 табл.
Формула
а) полинуклеотид, кодирующий одно- или многопроходный трансмембранный полипептид;
б) строго регулируемый промотор, имеющий низкую базальную активность в клетке-хозяине; причем промотор содержит по меньшей мере один позитивный регуляторный элемент контроля и по меньшей мере один негативный регуляторный элемент контроля; где один или более из позитивных и негативных регуляторных элементов контроля представляет собой гетерологичный регуляторный элемент контроля, и
в) лидерную последовательность, содержащую энхансер инициации трансляции.
а) промотор phoA, phac или tphac;
б) промотор Т7, metE или лямбда PL; или
в) промотор бета-лактамазы, лактозный или триптофановый промотор и по меньшей мере один гетерологичный регуляторный элемент контроля для снижения базальной активности.
а) введение экспрессирующей конструкции по любому из пп.1-33 в клетку-хозяина;
б) экспрессирование конструкции в клетке-хозяине с продуцированием трансмембранного полипептида.
а) дезинтеграцию клеток-хозяев, экспрессирующих полипептид;
б) разделение растворимой и нерастворимой фракций дезинтегрированных клеток; и
в) суспендирование нерастворимой фракции в неионном или цвиттер-ионном детергенте для солюбилизации и, тем самым, выделения полипептида из клеток-хозяев.
а) цвиттергент® 3-08, 3-10, 3-12, 3-16, ASB-14, ASB-16 или ASBC80;
б) производное фосфохолина;
в) додецилфосфохолин (DDPC), децилмальтозид (DDM), децилмальтозид (DM), н-додецил-N,N-диметиламин N-оксид (LADO), н-децил-N,N-диметиламин N-оксид, DHPC, DLPC, MPC, LOPC, LLPG; или
г) NP40.
а) дезинтеграцию клеток-хозяев, экспрессирующих полипептид;
б) разделение растворимой и нерастворимой фракций дезинтегрированных клеток; и
в) суспендирование нерастворимой фракции в неионном или цвиттер-ионном детергенте для солюбилизации и, тем самым, выделения полипептида из клеток-хозяев.
Комментарии