L-фукоза α1→6 специфичный лектин - RU2524425C2
Код документа: RU2524425C2
Чертежи






































Описание
Область техники
[0001]
Настоящее изобретение относится к новому L-фукоза α1→6-специфическому лектину, методу его получения и его применению. В частности, настоящее изобретение относится к новому лектину, получаемому из базидиального гриба или сумчатого гриба, методу его получения и методу обнаружения и фракционирования сахарной цепи с помощью лектина.
Уровень техники
[0002]
Известно, что ген α1→6 фукозилтрансферазы (FUT8), который переносит L-фукозный остаток к восстанавливающему концу N-ацетилглюкозамина N-гликана через α1→6-связь с фукозилированием ядра, проявляется в связи с появлением признаков злокачественности гепатоцитов. Злокачественная гепатома в настоящее время обнаруживается с помощью электрофореза по сродству к лектину с использованием агглютинина из Lens culinaris (LCA), обладающего сродством к фукозилированным моно- и биантенным N-гликанам.
[0003]
Антителозависимая клеточная цитотоксичность (далее ADCC-активность) представляет собой одну из иммунных функций человека. ADCC-активность - активность, благодаря которой лейкоциты, такие как естественные клетки-убийцы и моноциты, убивают клетки-мишени, такие как раковые клетки, с помощью антител. ADCC-активность связана с противоопухолевым механизмом действия медицинских средств на основе антител, таких как Гернептин в качестве гуманизированного антитела (терапевтическое средство для лечения метастатического рака молочной железы) и Ритуксан в качестве химерного антитела (терапевтическое средство для лечения неходжкинской лимфомы) (беспатентная публикация 1).
В том случае, если эти медицинские средства на основе антител обладают низкой ADCC-активностью, возникает необходимость в применении большой дозы медицинского средства на основе антител, что, в свою очередь, создает ряд проблем, таких как увеличение стоимости и побочные эффекты (например, инфицирование вследствие снижения иммунитета).
[0004]
ADCC-активность антитела, к которому переносится α1→6 L-фукоза, и антитела без α1→6 L-фукозы различается в 50-100 раз (Беспатентная публикация 2). В случае получения антитела, к которому α1→6 L-фукозы не переносится, возможно достижение высокой ADCC-активности.
[0005]
Традиционно наряду с LCA известны и другие фукозилированные лектины, такие как, например, агглютинин из Pisum sativum (PSA), лектип из Aleuria aurantia (AAL), агглютинин из Narcissus pseudonarcissus (NPA), агглютинин из Vicia faba (VFA) и лектин из Aspergillus oryzae (AOL) (Патентные публикации 1-5).
Беспатентная публикация 1: Clynes RA et at, Inhibitory Fc receptors modulate in vivo cytoxicity against tumor targets/Клайнс P.A. и др., Ингибирующие Fc-рецепторы модулируют in vivo цитотоксичность против опухолевых целей NATURE MED 2000 APR; 6(4):443-446
Беспатентная публикация 2: Toyohide Shinkawa et at., The absence of L-fucose but not the presence of galactose or bisecting N-acetyl glucosamine of human IgGl complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. (Тоехидэ Синкава и др., Отсутствие L-фукозы, а не присутствие галактозы или деление N-ацетилглюкозамина сложных олигосахаридов IgGI человека свидетельствует о критически важной роли усиления антителозависимой цитотоксичности) J Biol Chem. 2003 Jan 31; 278(5):3466-73. Epub 2002 Nov 8.
Патентная публикация 1: WO 2002/030954
Патентная публикация 2: WO 2003/084569
Патентная публикация 3: Пример из публикации нерассмотренной патентной заявки Японии No. H02-083337
Патентная публикация 4: Пример 5 из публикации нерассмотренной патентной заявки Японии No. 2002-112786
Патентная публикация 5: Публикация нерассмотренной патентной заявки Японии No. 2007-161633
Раскрытие изобретения
[0006]
Известные лектины, используемые для обнаружения L-фукоза α1→6 сахарной цепи, обладают также сродством не только к L-фукоза α1→6 сахарной цепи, но и к гликолипидной сахарной цепи, включающей L-фукозу, присоединенную не по связи α1→6, и сахарную цепь с высоким содержанием манноза, не содержащую L-фукозу. В частности, AAL и AOL обладают, например, сродством к α1→2 L-фукозе и α1→3 L-фукозе. Маннозоспецифичные LCA, PSA и VFA также обладают сродством к нефукозилиронным моно- и биантенным N-гликанам. Таким образом, не представлялось возможным точно идентифицировать L-фукоза α1→6 сахарную цепь, а также выделить L-фукоза α1→6 сахарную цепь. До настоящего времени не был известен лектин, способный связывать только L-фукоза α1→6 сахарную цепь,
[0007]
В свете вышеизложенного целью настоящего изобретения является создание нового лектина, способного к специфическому связыванию L-фукоза α1→6 сахарной цепи. Это изобретение позволяет создать более точный метод обнаружения L-фукоза α1→6 сахарной цепи с помощью нового лектина, в отличие от традиционного случая, и метод фракционирования L-фукоза α1→6 сахарной цепи и не содержащей L-фукоза α1→6 сахаридной цепи на основе обнаружения L-фукоза α1→6 сахарной цепи.
[0008]
Авторы настоящего изобретения обнаружили новый лектин, обладающий очень высоким сродством к сахарной цепи, включающей α1→6 связанную L-фукозу. Авторы настоящего изобретения обнаружили, что новый лектин дает возможность специфического обнаружения L-фукоза α1→6 сахарной цепи, а также что этот лектин можно использовать для очистки L-фукоза α1→6 сахарной цепи или сахарной цепи, не содержащей L-фукозу α1→6 (L-фукоза α1→2, 1→3,1→4 сахарная цепь). Термин "L-фукоза α1→6 сахарная цепь" означает структуру, в которой L-фукоза присоединена к восстанавливающему концу N-ацетилглюкозамина N-гликана по связи α1→6. Термин "сахарная цепь, не содержащая L-фукозу α1→6" означает сахарную цепь, в молекулах которой не содержится L-фукоза, присоединенная по связи α1→6.
[0009]
В частности, настоящее изобретение позволяет получить L-фукоза α1→6-специфичный лектин, который: (1) экстрагируется из базидиального гриба или сумчатого гриба, (2) имеет молекулярную массу в соответствии с методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE), равную 4000-40000, и (3) обладает сродством к L-фукоза α1→6 сахарной цепи, определяемым константой ассоциации 1,0×104M-1 или более (при 25 градусов C). Константа ассоциации здесь означает числовое значение, измеряемое с помощью фронтальной аффинной хроматографии (FAC) при температуре 25 градусов C.
[0010]
L-фукоза α1→6 сахарная цепь может включать сиало N-гликаны.
[0011]
Кроме того, желательно, чтобы L-фукоза α1→6-специфичный лектин (4) в значительной степени не связывался с высокоманнозными сахарными цепями и/или гликолипидами, не содержащими L-фукоза α1→6 сахарную цепь. [0012]
Кроме того, желательно, чтобы L-фукоза α1→6-специфичный лектин (5) обладал сродством к α1→6-фукозилированным моно-, ди-, три-, и тетраантенным N-гликанам с константой ассоциацией 1.0×104M-1 или более (при 25 градусов C).
[0013]
Базидиальный гриб принадлежит, в частности, к семействам Strophariaceae, Tricholomataceae, Amanitaceae или Polyporaceae.
[0014]
Базидиальными грибами являются, например, Pholiota terrestris, Pholiota squarrosa, Pholiota adiposa, Stropharia rugosoannulata, Naematoloma sublateritium, Lepista sordid и Amanita muscaria.
[0015]
В частности, аминокислотная последовательность L-фукоза α1→6-специфичного лектина (6) отображается номером 1.
[0016]
Настоящее изобретение позволяет также получить L-фукоза α1→6-специфичный лектин, который представляет собой (a) белок или пептид, включающий аминокислотную последовательность, описываемую одним из номеров от 2 до 5 или (b) белок или пептид, в котором одна или несколько аминокислот удалены, добавлены или замещены в аминокислотной последовательности, отображаемой одним из номеров от 2 до 5, и который функционально эквивалентен белку или пептиду с аминокислотной последовательностью, описываемой одним из номеров от 2 до 5. Термин "функционально эквивалентный" означает сродство к L-фукоза α1→6 сахарной цепи, характеризуемое константой ассоциации 1,0×104M-1 и более (при 25 градусах C).
[0017]
Белок или пептид, описываемый в пункте (b), имеет аминокислотную последовательность, описываемую, например, номером 6.
[0018]
Настоящее изобретение позволяет также получить L-фукоза α1→6-специфичный лектин, который представляет собой белок или пептид, по меньшей мере, с 37% сходством с аминокислотной последовательностью, описываемой одним из номеров от 2 до 6, и является функционально эквивалентным по отношению белку или пептиду с аминокислотной последовательностью, описываемой одним из номеров от 2 до 6.
[0019]
Настоящее изобретение позволяет также создать также метод получения L-фукоза α1→6-специфичного лектина, согласно которому водный экстракт (водорастворимый экстракт) базидиального и/или сумчатого гриба подвергается (i) гидрофобной хроматографии и обращенно-фазовой хроматографии, (ii) аффинной хроматографии или (iii) ионообменной хроматографии и гель-фильтрации с получением лектина, который (vi) имеет молекулярную массу, определяемую электрофорезом в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE) и равную 4000-40000, и который (v) характеризуется сродством, характеризуемым константой ассоциации L-фукоза α1→6 сахарной цепи 1,0×104M-1 или более (при 25 градусах C).
[0020]
Желательно, чтобы, по крайней мере, один базидиальный гриб выбирался из семейств Strophariaceae, Tricholomataceae, Amanitaceae и Polyporaceae.
[0021]
Желательно, чтобы, по крайней мере, один базидиальный гриб выбирался из Pholiota terrestris, Pholiota squarrosa, Pholiota adiposa, Stropharia rugosoanmilata, Naematoloma sublateritium, Lepista sordida и Amanita muscaria.
[0022]
Желательно, чтобы базидиальный и/или сумчатый гриб, используемый при получении L-фукоза α1→6-специфичного лектина, представлял собой карпофоры (плодовые тела).
[0023]
Настоящее изобретение позволяет также создать метод обнаружения L-фукоза α1→6-сахарной цепи, включающий процесс, приводящий к воздействию сахарной цепи на L-фукоза α1→6-специфичный лектин.
[0024]
Сахарная цепь является, в частности, онкомаркером.
[0025]
Настоящее изобретения позволяет также создать метод фракционирования L-фукоза α1→6 сахарной цепи, включающий процесс, приводящий к воздействию сахарной цепи на L-фукоза α1→6-специфичный лектин. В частности, изобретение позволяет создать метод фракционирования L-фукоза α1→6 сахарной цепи и не содержащей L-фукозы α1→6 сахарной цепи путем применения иммобилизованного L-фукоза α1→6-специфичного лектина.
[0026]
Сахарная цепь, используемая для метода фракционирования, связывается, например, с антителом.
[0027]
Настоящее изобретение позволяет также создать диагностическое средство и диагностический комплект для обнаружения L-фукоза α1→6 сахарной цепи. Диагностическое средство включает в качестве активного компонента L-фукоза α1→6-специфичный лектин.
[0028]
Новый лектин, описываемый в настоящем изобретении, обладает гораздо более высоким сродством к сахаридной цепи, гликопептидам и гликопротеину с α1→6 связанной L-фукозой, чем обычный лектин, и имеет константу ассоциации 1,0×104M-1 или более. В частности, возможно специфическое распознавание только сахарной цепи с L-фукоза α1→6 структурой цепи.
Обладающий такой специфичностью, L-фукоза α1→6-специфичный лектин может использоваться в различных областях применения, как показано ниже.
[0029]
В отличие от обычного лектина, обладающего сродством к L-фукоза α1→6 сахарной цепи, настоящее изобретение позволяет обнаруживать α1→6 L-фукоза сахарную цепь с большей специфичностью.
[0030]
Метод фракционирования L-фукоза α1→6 сахарной цепи настоящего изобретения дает возможность обеспечить на основе точной идентификации α1→6 L-фукозы более строгое фракционирование между L-фукоза α1→6 сахарной цепью и не содержащей L-фукозы α1→6 сахарной цепью. В результате L-фукоза α1→6 сахарная цепь или не содержащая L-фукозы α1→6 сахарная цепь очищаются с высокой степенью чистоты. В частности, путем использования описанного здесь метода фракционирования для удаления L-фукоза α1→6 сахарной цепи из препарата на основе антитела, включающего смесь L-фукоза α1→6 сахарной цепи и не содержащей L-фукозы α1→6 сахарной цепи, можно получить лекарства на основе антител с повышенной ADCC-активностью. В результате препарат на основе антитела можно вводить в пониженной дозе, тем самым удается с успехом добиться, например, снижения стоимости препарата и уменьшить влияние побочных эффектов. Кроме того, препарат на основе антитела можно прописывать в соответствии с симптомом или побочным эффектом.
Перечень рисунков
[0031]
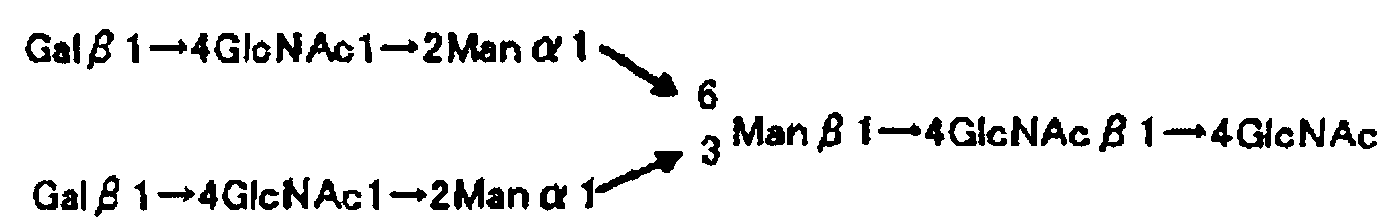
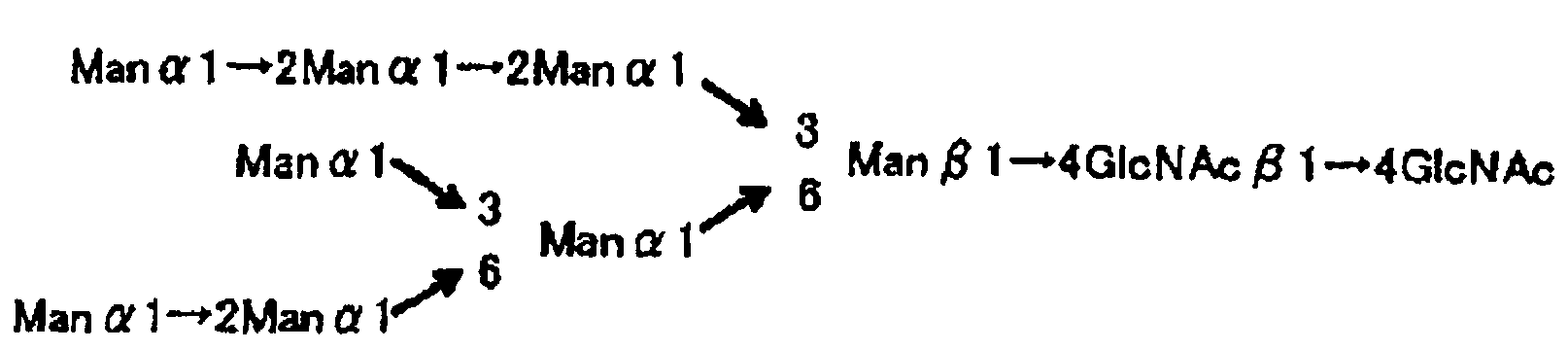
Рис.1 представляет собой структурную схему α1→6 L-фукоза олигосахарида и не содержащего α1→6 L-фукозы олигосахарида, используемую в рабочем примере и сравнительном примере настоящего изобретения.
Рис.2 представляет собой структурную схему L-фукоза α1→6 олигосахарида и не содержащего L-фукозу α1→6 олигосахарида, используемую в рабочем примере и сравнительном примере настоящего изобретения.
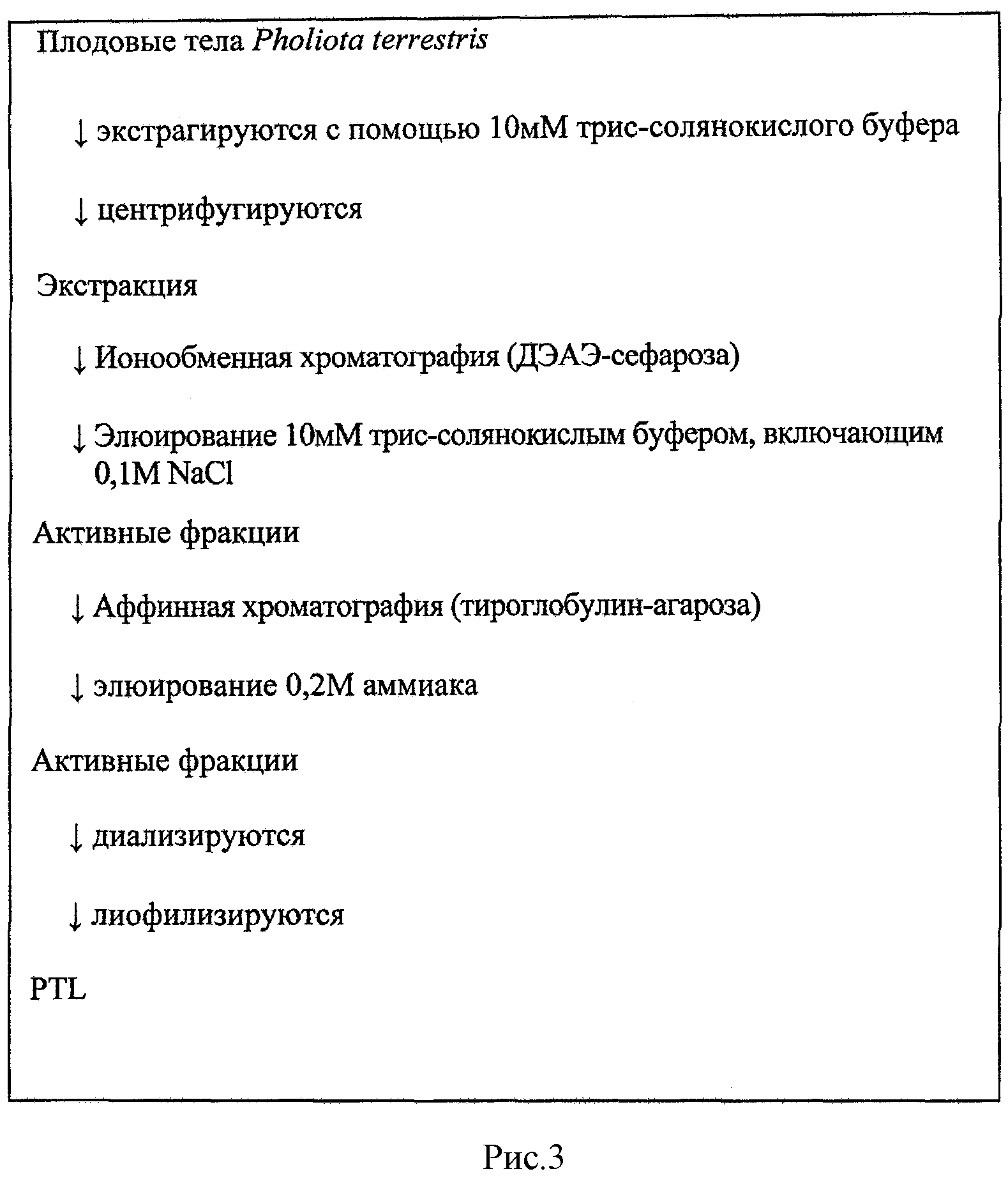
Рис.3 представляет процесс очистки PTL из примера 1.
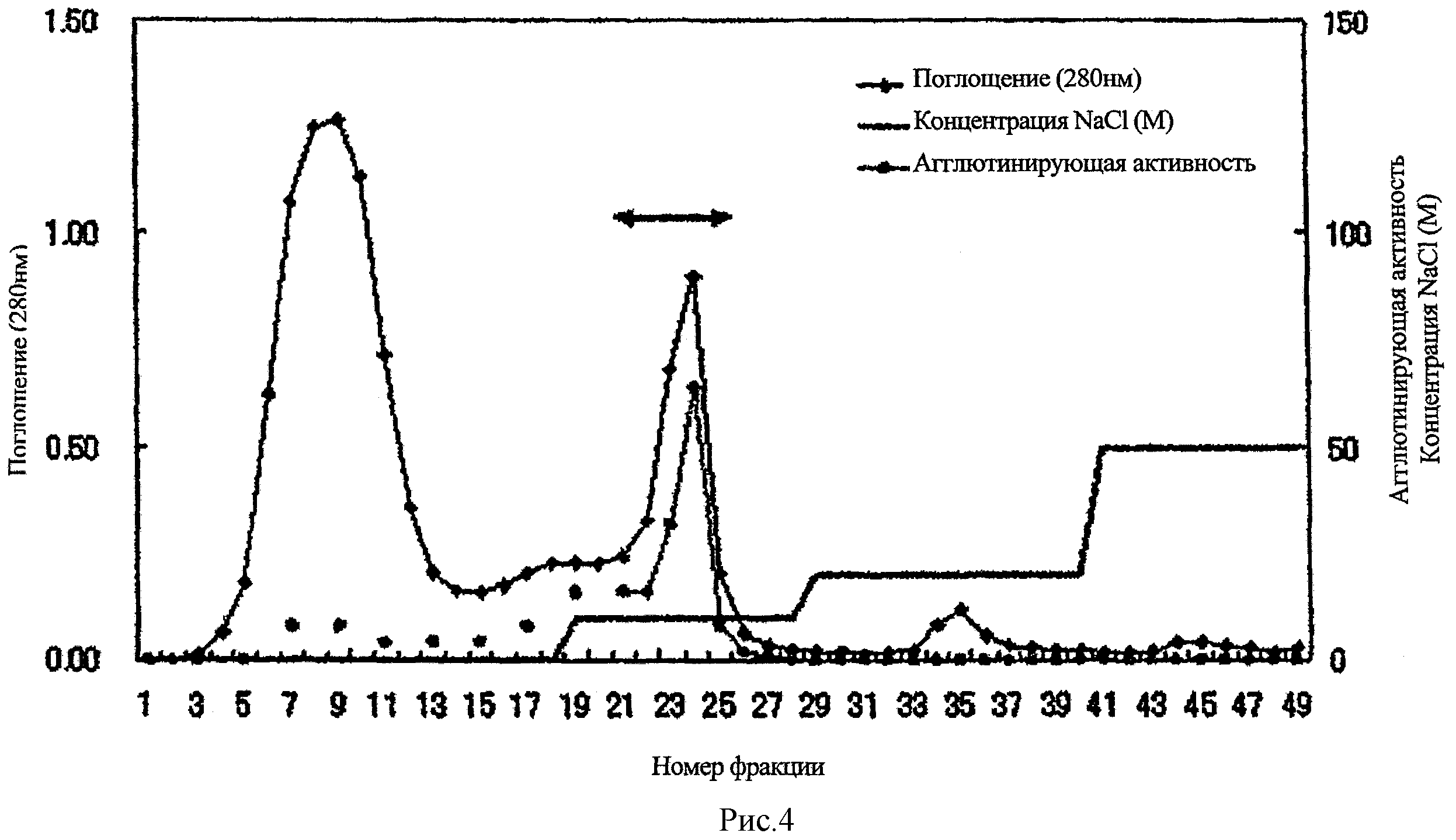
Рис.4 представляет собой схему элюирования при ионообменной хроматографии PTL из примера 1.
Рис.5 представляет собой схему элюирования при аффинной хроматографии PTL из примера 1.
На рис.6 показан результат электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE) PTL из примера 1 (фотография вместо рисунка).
На рис.7 показан процесс очистки SRL из примера 2.
На рис.8 представлена схема элюирования при гидрофобной хроматографии SRL из примера 2.
На рис.9 представлена схема элюирования при обращенно-фазовой хроматографии SRL из примера 2.
На рис.10 показан результат электрофореза SDS-PAGE SRL из примера 2 (фотография вместо рисунка).
На рис.11 показан результат масс-спектроскопии PTL из примера 1.
На рис.12 показан результат масс-спектроскопии SRL из примера 2.
На рис.13 показан результат Вестерн-блоттинга с использованием PTL из примера 1 (фотография вместо рисунка).
На рис.14 показан результат Вестерн-блоттинга с использованием SRL из примера 2 (фотография вместо рисунка).
На рис.15 показан результат Вестерн-блоттинга с использованием AAL из сравнительного примера Comparison Example 1 (фотография вместо рисунка).
На рис.16 показан результат Вестерн-блоттинга с использованием AOL из сравнительного примера 2 (фотография вместо рисунка).
На рис.17 показан результат Вестерн-блоттинга с использованием LCA из сравнительного примера 3 (фотография вместо рисунка).
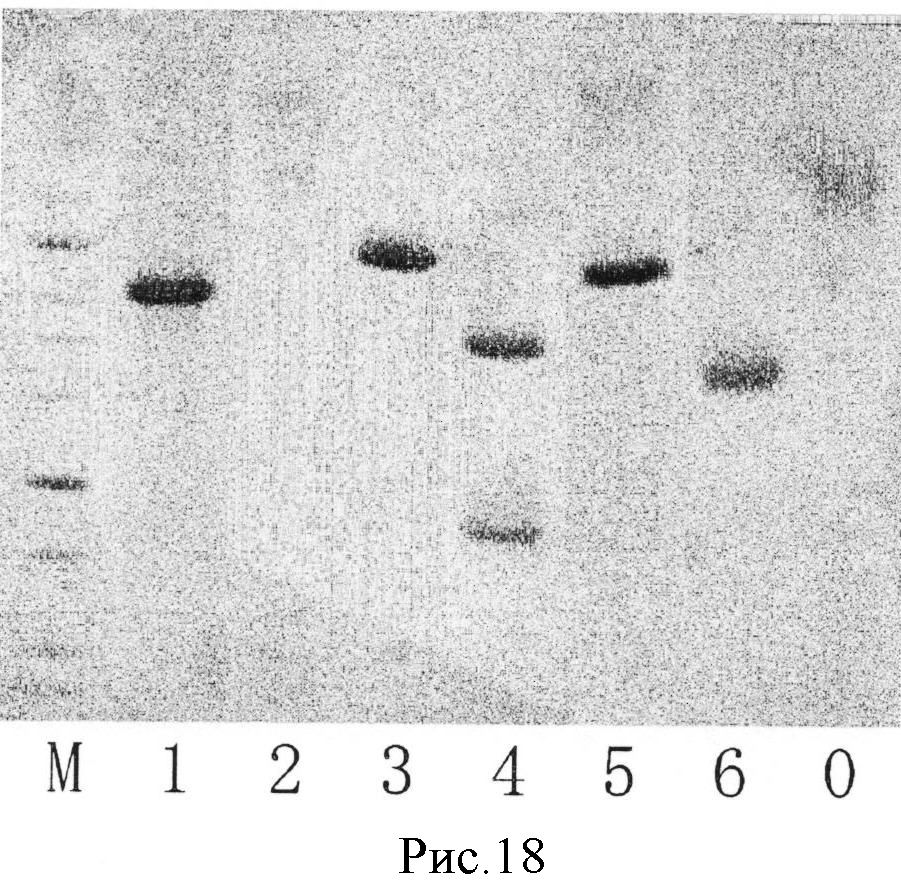
На рис.18 показан результат, когда только белки окрашиваются СВВ (кумасси ярко-синий) для контроля (фотография вместо рисунка).
На рис.19 показан результат обнаружения гликопротеинов методом твердофазного иммуноферментного анализа ELISA с использованием PTL из примера 1.
На рис.20 показан результат обнаружения гликопротеинов методом ELISA с использованием SRL из примера 2.
На рис.21 показан результат обнаружения гликопротеинов методом ELISA с использованием AAL из сравнительного примера 1.
На рис.22 показан результат обнаружения гликопротеинов методом ELISA с использованием AOL из сравнительного примера 2.
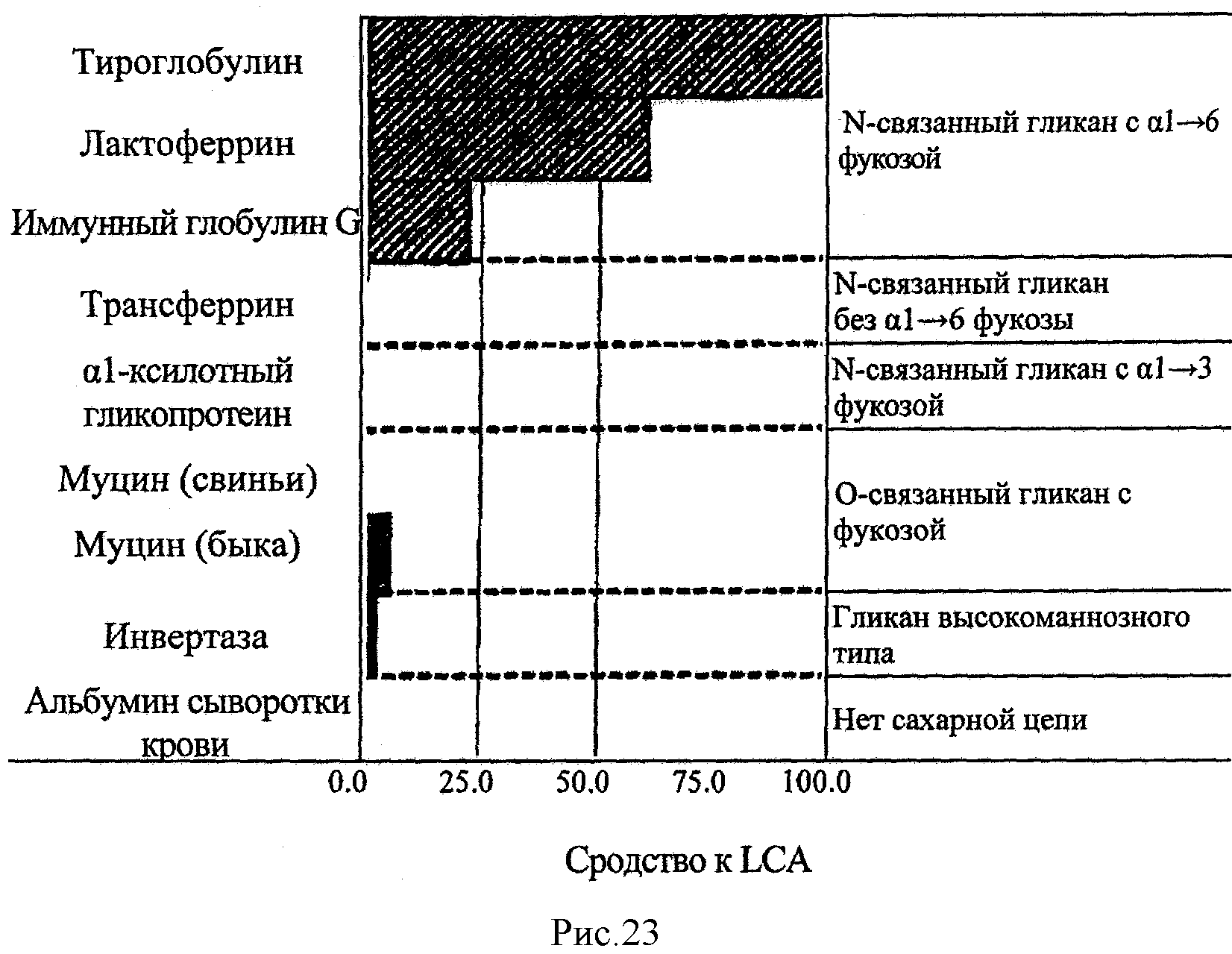
На рис.23 показан результат обнаружения гликопротеинов методом ELISA с использованием LCA из сравнительного примера 3.
На рис.24 показан результат обнаружения разницы в сахарной цепи между α-фетопротеинами L1 и L3 методом ELISA с использованием PTL из примера 1.
На рис.25 показан результат обнаружения разницы в сахарной цепи между α-фетопротеинами L1 и L3 методом ELISA с использованием SRL из примера 2.
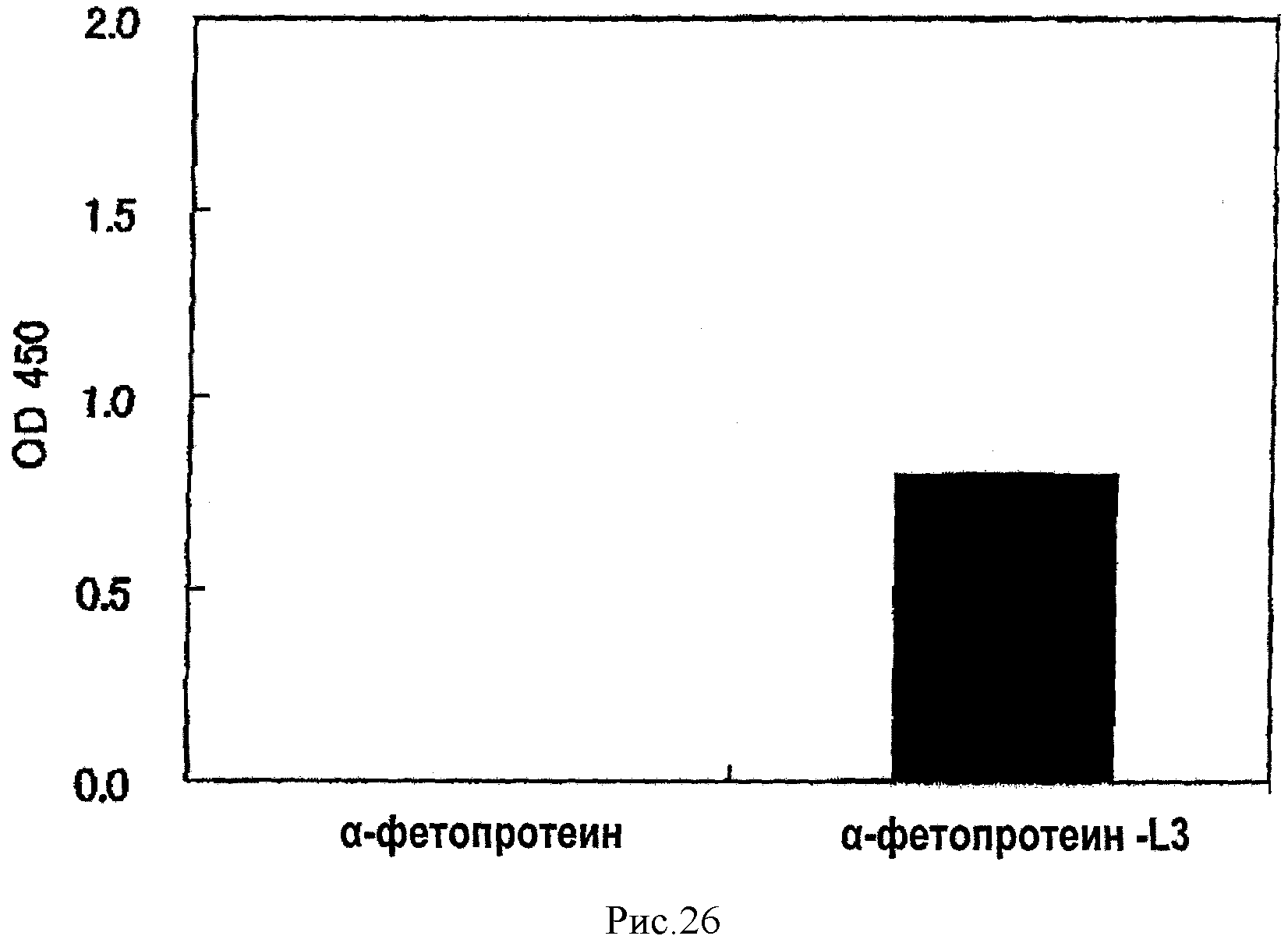
На рис.26 показан результат обнаружения разницы в сахарной цепи между α-фетопротеинами L1 и L3 методом ELISA с использованием AAL из сравнительного примера 1.
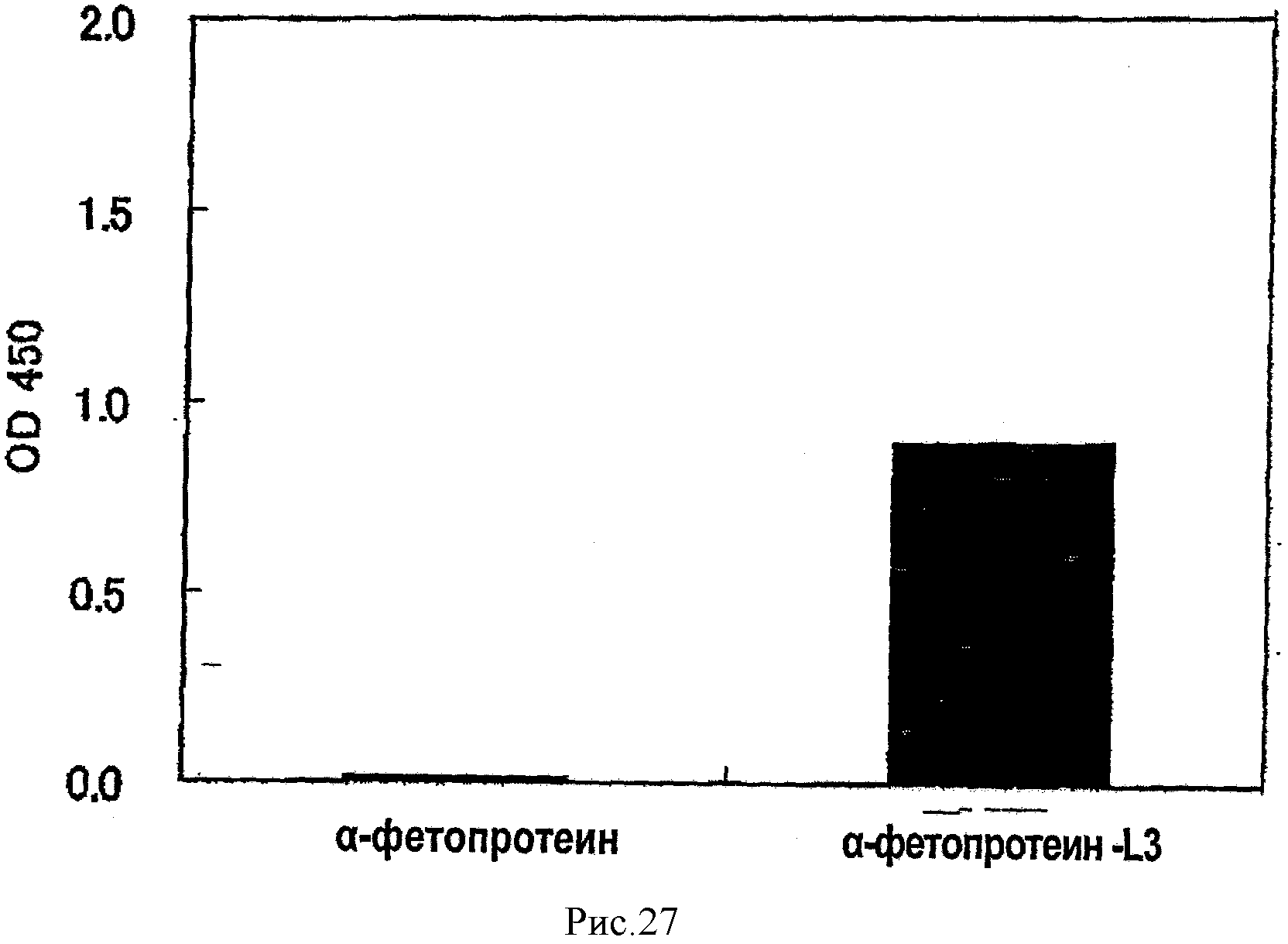
На рис.27 показан результат обнаружения разницы в сахарной цепи между α-фетопротеинами L1 и L3 методом ELISA с использованием AOL из сравнительного примера 2.
На рис.28 показан результат обнаружения разницы в сахарной цепи между α-фетопротеинами L1 и L3 методом ELISA с использованием LCA из сравнительного примера 3.
На рис.29 показан процесс очистки NSL из примера 3.
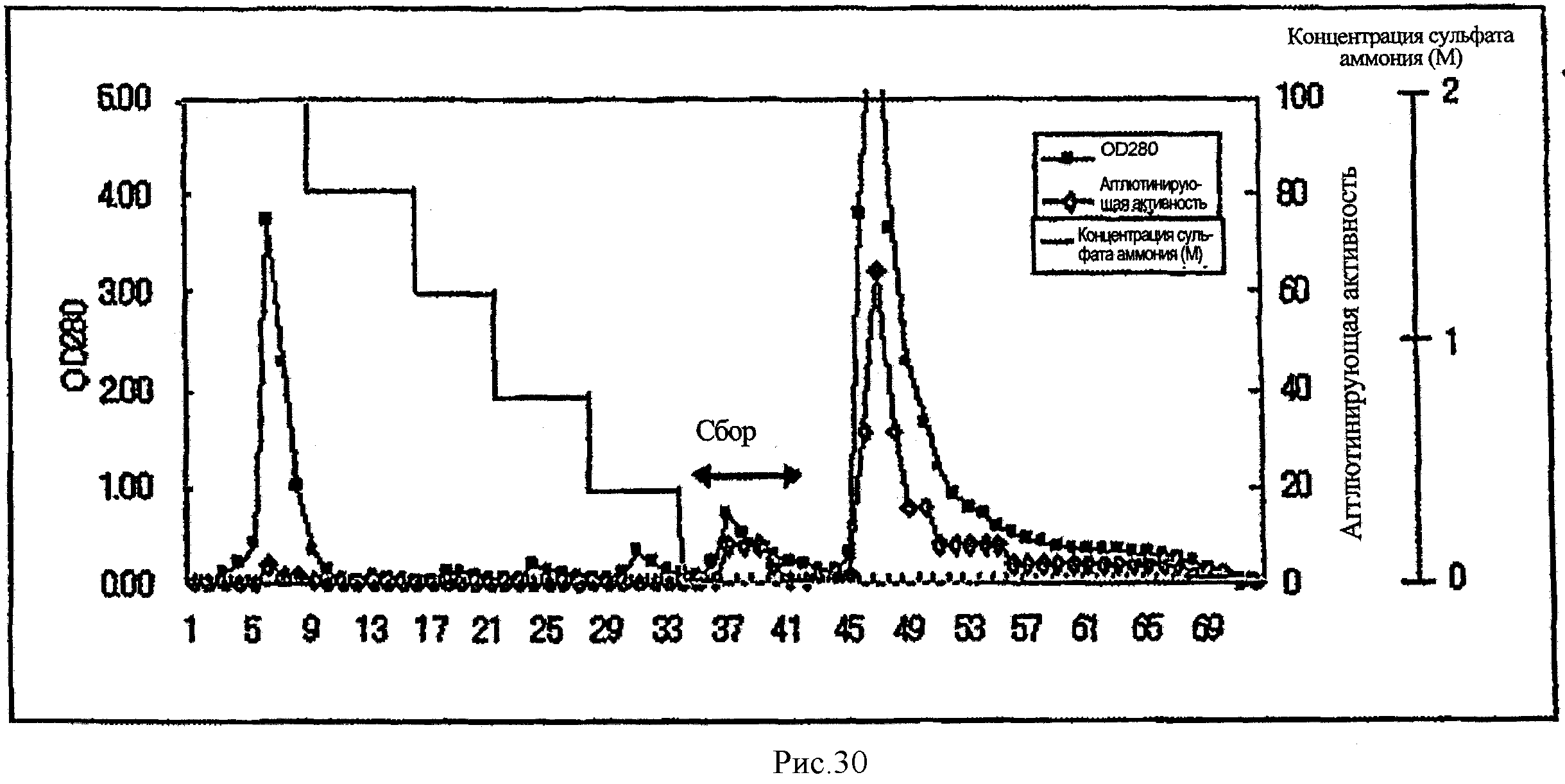
На рис.30 показана диаграмма элюирования при гидрофобной хроматографии NSL из примера 3.
На рис.31 показана диаграмма элюирования при обращенно-фазовой хроматографии NSL из примера 3.
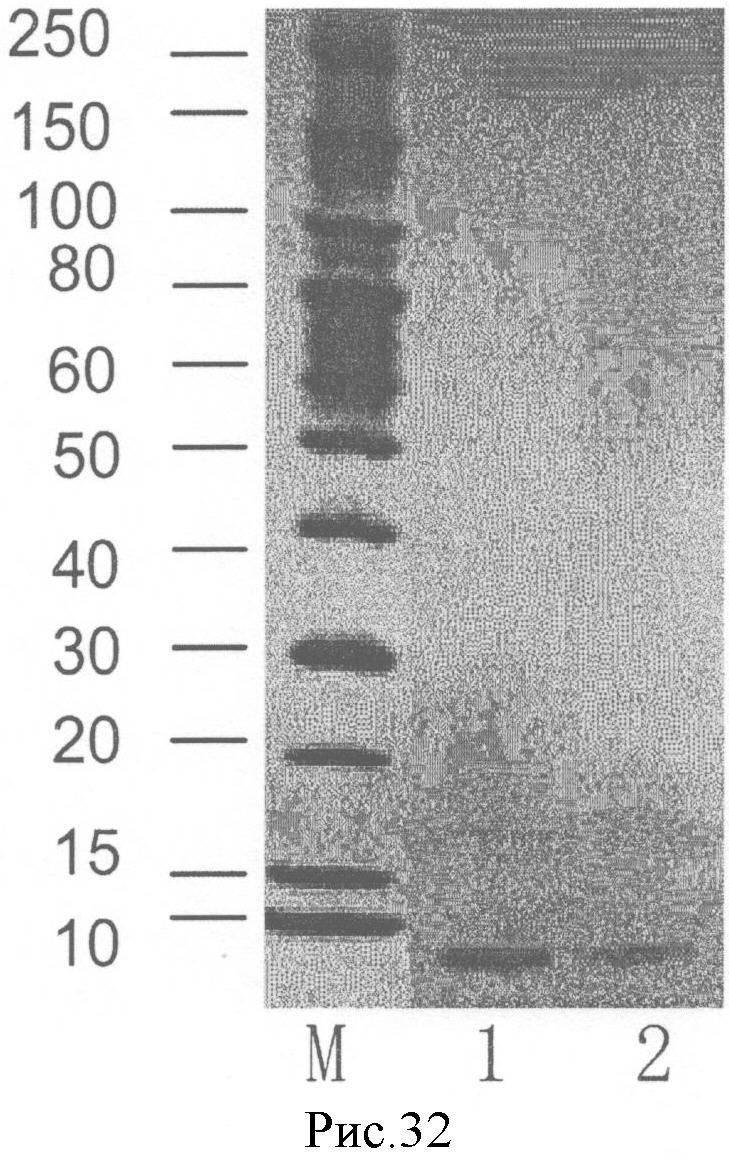
На рис.32 показан результат электрофореза SDS-PAGE NSL из примера 3 (фотография вместо рисунка).
На рис.33 показан процесс очистки LSL из примера 4.
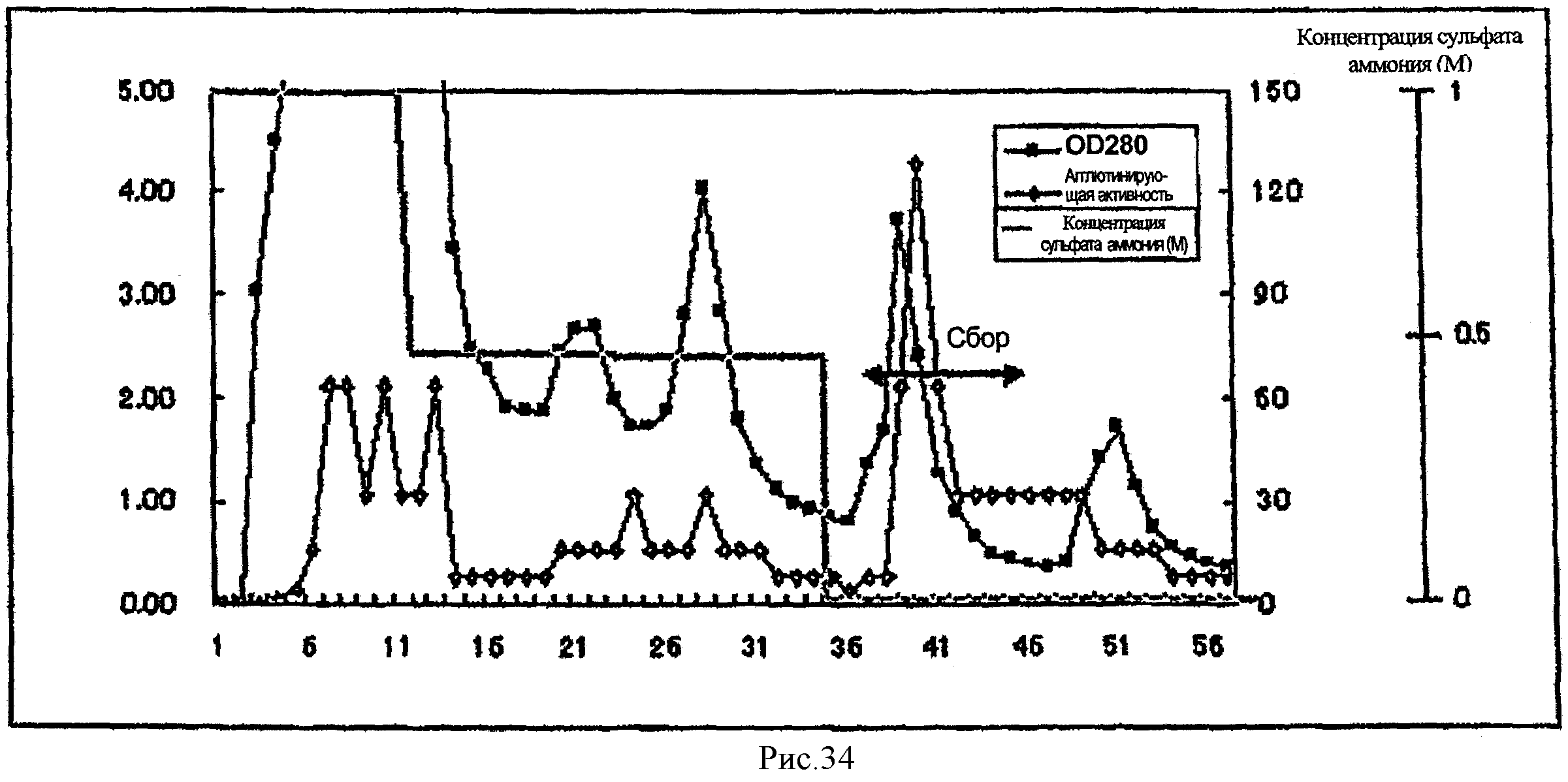
На рис.34 показана диаграмма элюирования при гидрофобной хроматографии LSL из примера 4.
На рис.35 показана диаграмма элюирования при обращенно-фазовой хроматографии LSL из примера 4.
На рис.36 показан результат SDS-PAGE LSL из примера 4 (фотография вместо рисунка).
На рис.37 показан результат масс-спектроскопии NSL из примера 3.
На рис.38 показан результат масс-спектроскопии LSL из примера 4.
Наилучший способ применения изобретения
[0032]
В следующем разделе представлен пример L-фукоза α1→6 сахарной цепи, с которой связан L-фукоза α1→6-специфичный лектин.
[0033]
[Химическая формула 1]

[В формуле Man означает маннозу, GlcNAc означает N-ацетилглюкозамин, a Fuc означает L-фукозу.]
[0034]
Кроме того, L-фукоза α1→6 сахарные цепи включают, например, свободный олигосахарид, гликоаминовую кислоту, гликопептид, гликолипид, гликопротеин, протеогликан и клетки. Кроме того, L-фукоза α1→6 сахарные цепи могут флуоресцентно окрашиваться, например, с помощью CyDye, 4-этиламинобензоата (ABEE) и аминопиридина. N-сахарная цепь включает также, например, высокоаммозную цепь, цепь комплексного и гибридного типа. Кроме того, N-сахарная цепь также может быть получена частичным разложением сахарной цепи химическим путем с помощью, например, кислоты или гидразина или с помощью одновременного или последовательного применения ферментов сиалидазы, галактозидазы, N-ацетилгексозаминидазы, фукозидазы и маннозидазы для частичного разложения сахарной цепи. Или N-сахарная цепь может быть также получена добавлением сахара, например глюкозы, или функциональной группы, например ацетильной группы, сульфатной группы или фосфатной группы, к сахарной цепи.
[0035]
(1) Базидиальный или сумчатый гриб, из которых получают L-фукоза α1→6-специфичный лектин относятся, например, к семействам Strophariaceae, Tricholomataceae, Polyporaceae и Amanitaceae. Strophariaceae включает, в частности, Pholiota terrestris, Stropharia rugosoannulata, Naematoloma sublateritium, Pholiota squarrosa, Pholiota adipose и Pholiota adiposa. Tricholomataceae включает, в частности, Lepista sordida. Polyporaceae включает, в частности, Trichaptum elongatum и Microporus affinis. Amanitaceae включает, в частности, Amanita muscaria. Среди этих базидиальных или сумчатых грибов особенно предпочтительны в плане эффективности извлечения лектина и специфичности к связыванию сахара, семейства Strophariaceae, Tricholomataceae и Amanitaceae.
Другими предпочтительными базидиальными и сумчатыми грибами являются Pholiota terrestris, Pholiota squarrosa, Pholiota adiposa, Stropharia rugosoannulata, Naematoloma sublateritium, Lepista sordid и Amanita muscaria.
[0036]
L-фукоза α1→6-специфичный лектин имеет (2) молекулярную массу, определенную с помощью электрофреза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE), 4000-40000, предпочтительно 4000-20000. Молекулярная масса SDS-PAGE измеряется, например, методом
Лемми (Laemmi) (Nature, том 227, страница 680, 1976).
[0037]
L-фукоза α1→6-специфичный лектин обладает (3) константой ассоциации (Ka) с L-фукоза α1→6 сахарной цепью, равной 1,0×104M-1 и более, предпочтительно 1,0×105M-1 и более или более предпочтительно 1,0×106M-1 и более. В частности, по сравнению с AAL, AOL, LCA, NPA и PSA, которые, как известно, обладают сродством к α1→6 связанной L-фукозе, L-фукоза α1→6-специфичный лектин имеет исключительно высокую константу ассоциации. Это означает, что L-фукоза α1→6 специфичный лектин связывается с L-фукоза α1→6 сахарной цепью с гораздо более высокой селективностью, чем обычный лектин.
[0038]
L-фукоза α1→6 сахарная цепь может иметь сиаловую кислоту на нередуцирующем конечном участке цепи. Обычный специфичный лектин с фукозой в ядре (например, LCA, NPA и PSA) имеет низкое сродство к L-фукоза α1→6 сахарной цепи с сиаловой кислотой на невосстанавливающем конце. С другой стороны, L-фукоза α1→6-специфичный лектин превосходит обычный с точки зрения высокого сродства также и к сахарной цепи, как говорилось выше.
[0039]
В следующем разделе описывается метод расчета константы ассоциации с помощью фронтальной аффинной хроматографии (FАС). Этот метод основан на следующем принципе. Когда разбавитель с определенной концентрацией флуоресцентно окрашенной сахарной цепи (например, показанной на рис.1 и 2) подается в колонку, где находится иммобилизованный лектин, сахарная цепь через короткое время выходит из колонки, если между лектином и сахарной цепью не происходит взаимодействия. Затем немедленно наблюдают фронт элюирования. Если существует сродство к лектину, элюирование сахарной цепи происходит с запаздыванием.
[0040]
Подготовка колонки с лектином, используемой в приборе, выполняется следующим образом. 1. Очищенный лектин растворяют в 0,1-0,2 M буферном растворе NaHCO3 (pH 8,3-8,5). 2. Лектин иммобилизуется по первичной аминогруппе, носителем является, например, активированная NHS-сефароза. 3. Далее, лектин блокируется раствором трис-буфера, включающим, например, первичный амин или этаноламин. 4. Лектин-сефароза суспендируются в 10 м молях трис-буфера, включающего 0.8% NaCl (pH 7.4, TBS). Далее смола с иммобилизованным лектином загружается в миниатюрную колонку (φ2 мм* 10 мм, 31.4 мкл). 5. Миниатюрная колонка, в которой находится смола с иммобилизованным лектином, фиксируется держателем, после чего колонка с лектином подключается к автоматическому анализатору FAC (FAC-1, SHIMADZU CORPORATION).
[0041]
В уравновешенную колонку с лектином заливают при скорости потока 0,125 мл/мин в количестве 300 мкл пиридиламинированную сахарную цепь (PA-сахарная цепь), разбавленную аналитическим буферным раствором (10 ммолей трис-буфера, включающего 0.8% NaCl (pH 7,4, TBS)) до концентрации (2,5 наномолей) в достаточной степени более низкой, чем константа диссоциации (Kd) лектина. Далее с помощью детектора флуоресцещтого излучения идентифицируется PA-сахарная цепь, выходящая из колонки (длина волны возбуждения/длина волны флуоресцентного излучения: 310 нм/380 им).
[0042]
На основе данных детектора, используя фронт элюирования (V0) сахарной цепи (PA-рамноза), не взаимодействующей с лектином как эталон, в качестве показателя интенсивности взаимодействия рассчитывается запаздывание (V-V0) фронта элюирования (V) сахарной цепи, взаимодействующей с лектином. Далее, на основе следующих критериальных уравнений FAC, исходя из V-V0 и Bt рассчитывается константа ассоциации (Ka) между сахарной цепью и лектином. Если интенсивность взаимодействия (значение V-V0) и константа ассоциации более высокие, наблюдается более высокое сродство между лектином и L-фукоза α1→6 сахарной цепью,
[0043]
[Уравнение 1]



[В этом уравнении A означает вещество, используемое для элюирования, A0 означает начальную концентрацию вещества A, B - иммобилизованный лиганд, V - элюирующий объем, V0 - элюирующий «фронтальный» объем для вещества, не взаимодействующего с иммобилизованным лигандом B, Bt - эффективное количество лиганда, Kd - константа диссоциации (величина, обратная константе ассоциации)].
[0044]
Специфичность лектина к связыванию сахара подтверждается также путем использования эритроцитов, которые подвергаются специфической агглютинации лектином с целью исследования типа сахара, который может подавлять агглютинацию эритроцитов и его концентрации.
[0045]
Кроме того, желательно, чтобы L-фукоза α1→6-специфичный лектин (4) практически не связывался с высокоманнозной сахарной цепью и/или гликолипидом, не включающим L-фукоза α1→6 сахарную цепь. Таким образом, L-фукоза α1→6-специфичный лектин обладает более высокой связывающей специфичностью. Выражение "практически не связывался" здесь означает константу ассоциации 1,0×103M-1 или меньше, предпочтительно константу ассоциации 1,0×102M-1 или меньше и, в частности, константу ассоциации, равную 0.
[0046]
Кроме того, желательно, чтобы L-фукоза α1→6-специфичный лектин (5) имел сродство к α1→6 фукозилированным, моно-, ди-, три- и тетраантенным N-гликанам. Это сродство характеризуется константой ассоциации 1,0×104M-1 и более (при 25 градусах C), предпочтительно - константой ассоциации 1.0×105M-1 и более.
[0047]
Примеры структур α1→6 фукозилированных моно-, ди-, три- и тетраантенных N-гликанов, обладающих сродством к описанному здесь лектину, представлены ниже.
[Химическая формула 2]


[0048]
L-фукоза α1→6-специфичный лектин, описываемый в настоящем изобретении, имеет обычную аминокислотную последовательность, в частности, представленную номером 1. 4ая, 5ая, 6ая и 7ая Xaas в последовательности 1 означает Asp/Asn/Glu/Thr, Thr/Ser/Ala, Tyr/Phe и Gln/Lys/Glu соответственно, причем диагональные разделительные линии означают "или".
[0049]
Конкретный пример L-фукоза α1→6-специфичного лектина, описываемого в настоящем изобретении, - протеин или пептид, представленный номерами 1-6.
[0050]
Лектин, описываемый номером последовательности 2, представляет собой новый лектин, экстрагируемый из Pholiota terrestris (далее PTL). 10ый и 17ый Xaas номера 2 могут представлять собой любой аминокислотный остаток, предпочтительно Cys. 20ый, 23ий, 27*, 33ий, 35ый и 39ый Xaas представляют собой Tyr/Ser, Phe/Tyr, Arg/Lys/Asn, Asp/Gly/Ser, Asn/Ala и Thr/Gln соответственно.
[0051]
Лектин, представленный номером 3, представляет собой новый лектин, экстрагируемый из Stropharia rugosoannulata (далее SRL). 10ый и 17ый Xaas номера 3 могут представлять собой любой аминокислотный остаток, предпочтительно Cys. 4ый, 7ой, 9ый, 13ый, 20ый, 27ой, 29ый, 33ий, 34ый и 39ый Xaas представляют собой Pro/Gly, Glu/Lys, Val/Asp, Asn/Asp/Glu, His/Ser, Lys/His, Val/Ile, Gly/Asn/Ser, Ala/Thr и Arg/Thr соответственно.
[0052]
Лектин, представленный порядковым номером 4, представляет собой новый лектин, экстрагируемый из Lepista sordida (далее LSL). 10ый и 17ый Xaas номера 4 могут представлять собой любой аминокислотный остаток, предпочтительно Cys. 1ый, 4ый, 7ой, 8ой, 9ый, 13ый, 16ый, 20ый, 22ой, 25ый, 27ый, 31ый и 34tый Xaas представляют собой Ala/Gin, Pro/Lys, Ala/Ser, Met/Ile/Val, Tyr/Thr, Asp/Asn, Lys/Glu, Ala/Asn, Val/Asp/Asn, Asp/Asn, Arg/His/Asn, Gin/Arg и Thr/Val соответственно.
[0053]
Лектин, представленный порядковым номером 5, представляет собой новый лектин, экстрагируемый из Naematoloma sublateritium (далее NSL). 10ый и 17ый Xaas номера 5 могут представлять собой любой аминокислотный остаток, предпочтительно Cys. 13ый, 14ый и 16tый Xaas представляют собой Asp/Thr, Ser/Ala, и Gln/Lys соответственно.
[0054]
Лектин, представленный порядковым номером 6, представляет собой новый лектин, экстрагируемый из Naematoloma sublateritium (далее NSL). Номер последовательности 6 описывает вариант, в котором одна Asn присоединена к пептиду с номером 5. Таким образом, 10ый и 18ый Xaas номера 6 могут представлять собой любой аминокислотный остаток, но предпочтительно Cys. 14ый, 15ый и 17ый Xaas представляют собой Asp/Thr, Ser/Ala и Gln/Lys соответственно.
[0055]
Поскольку белки или пептиды, имеющие аминокислотные последовательности SEQ ID NО: 2-6 являются новыми, настоящее изобретение посвящено L-фукоза α1→6-специфичному лектину, который является (a) белком или пептидом с аминокислотной последовательностью, выбранной из SEQ ID NO: 2-5 или (b) белком или пептидом, в котором одна или несколько аминокислот удалены, добавлены или замещены в одной из последовательностей SEQ ID NO: 2-5, который функционально эквивалентен белку или пептиду с аминокислотной последовательностью SEQ ID NO: 2-5. Термин «функционально эквивалентен» здесь означает сродство с константой ассоциации 1,0×104M-1 или более к L-фукоза α1→6 сахарной цепи, предпочтительно 1.0×105M-1 или более и более предпочтительно - 1.0×106M-1 и более. Примером варианта, показанного в пункте (b), является белок или пептид, имеющий аминокислотную последовательность SEQ ID NO: 6.
[0056]
Настоящее изобретение посвящено также гену, определяющему (a) белок или пептид с аминокислотной последовательностью, выбранную из SEQ ID NO: 2-5 или (b) белок или пептид, в котором одна или несколько аминокислот удалены, добавлены или замещены в одной из последовательностей SEQ ID NO: 2-5 и который функционально эквивалентен белку или пептиду с аминокислотной последовательностью, выбранную из SEQ ID NO: 2-5. Выражение "функционально эквивалентен" имеет то же значение, что и выше.
[0057]
Степень сходства между белками и пептидами, SEQ ID NO: 2-6. составляет, по меньшей мере, 37% (см. таблицу 14). Таким образом, настоящее изобретение позволяет также получить L-фукоза α1→6-специфичный лектин, который является белком или пептидом со степенью сходства как минимум 37% и более с аминокислотной последовательностью, выбранной из SEQ ID NO: 2-6, и который функционально эквивалентен белку или пептиду с аминокислотной последовательностью, выбранной из SEQ ID NO: 2-5. Выражение "функционально эквивалентен" имеет то же значение, что и выше.
[0058]
L-фукоза α1→6-специфичный лектин может быть выделен из базидиального и/или сумчатого гриба, например, с помощью подходящего сочетания известного метода экстракции, метода разделения и метода очистки. В частности, возможен процесс с использованием водного растворителя с целью получения водорастворимого экстракта базидиального и/или сумчатого гриба. Из этого экстракта получают лектин с (vi) молекулярной массой, определяемой методом SDS-PAGE и равной 4000-40000, предпочтительно 4000-20000 и со (v) сродством к L-фукоза α1→6 сахарной цепи, описываемым константой ассоциации, равной 1,0×104M-1 и более, предпочтительно 1,0×105M-1 и более и более предпочтительно 1,0×106M-1 и более (при 25 градусах C).
[0059]
Желательно, чтобы базидиальный гриб был выбран из, по крайней мере, одного из семейств Strophariaceae, Tricholomataceae, Polyporaceae и Amanitaceae. В частности, к Strophariaceae относятся, например, Pholiota terrestris (Pholiota terrestris Overholts), Pholiota squarrosa (Pholiota squarrosa (Fr.) Kummer), Pholiota adiposa (Pholiota adiposa (Fr.) Kummer), Stropharia rugosoannulata (Stropharia rugosoannulata Farlow in Murr.), Naematoloma sublateritium (Naematoloma sublateritium (Fr.) Karst или Hypholoma sublateritium (Fr.) Quel), к Tricholomataceae относится, например, Lepista sordida (Lepista sordida (Schum.: Fr.) Sing.), к Polyporaceae относятся, например, Trichaptum elongatum (Trichaptum elongatum), Microporus affmis (Microporus vemicipes), к Amanitaceae относится, например, Amanita muscaria (Amanita muscaria). Для всех указанных базидиальных и/или сумчатых грибов предпочтительно использование карпофора (плодового тела).
[0060]
Метод получения экстракта базидиального гриба не имеет ограничений, до тез пор пока метод обеспечивает, например, контакт водного растворителя с карпофором базидиального гриба. С точки зрения эффективности экстракции предпочтителен метод, при котором карпофор базидиального гриба распыляется в водной среде с получением суспензии. В качестве метода распыления может применяться традиционный метод с использованием, например, миксера или гомогенизатора.
[0061]
Водный растворитель может представлять собой буферный раствор или смесь воды или буферного раствора и органического растворителя, который может смешиваться, например, с водой и предпочтительно представляет собой буферный раствор или смесь органического растворителя и буферного раствора.
[0062]
Вид буферного раствора не ограничивается, он может представлять собой известные буферные растворы, в числе которых особенно предпочтительно применение буферного раствора с интервалом буферирования pH 3-10 и еще более предпочтительно - буферного раствора с интервалом буферирования pH 6-8. В частности, возможно применение фосфатного буфера, нитратного буфера, уксуснокислого буфера и трис-буфера, из которых с точки зрения эффективности экстракции предпочтителен фосфатный буфер.
[0063]
Буферный раствор не ограничивается по конкретной предельной концентрации соли. С точки зрения эффективности экстракции и интервала буферирования желательна концентрация соли 1-100 ммоль и более, предпочтительна концентрация соли 5-20 ммоль.
[0064]
В буферный раствор можно добавлять соль. Например, изотонический раствор хлорида натрия, забуференный фосфорной кислотой и полученный добавлением пищевой соли к фосфатному буферу, желательно использовать в качестве водного растворителя для целей настоящего изобретения.
[0065]
Органический растворитель может представлять собой любой органический растворитель, смешивающийся с водой без каких-либо ограничений, в частности, предпочтительно применение ацетона, метанола, этанола, 2-пропанола и ацетонитрила. Предпочтительно смешать органический растворитель с водой или буферным раствором в концентрации 10-40% (массовых).
[0066]
Желательно, чтобы процесс экстракции дополнительно включал, в частности, процесс удаления из смеси водного растворителя и карпофора базидиального гриба веществ, нерастворимых в водной среде. Метод удаления нерастворимых веществ может представлять собой, например, фильтрацию или центрифугирование, причем с точки зрения эффективности удаления предпочтительно центрифугирование.
[0067]
Желательно, чтобы процесс экстракции обеспечивал распыление карпофора базидиального гриба, например, в фосфатно забуференной соли, для удаления нерастворимых веществ с помощью центрифугирования с целью получения экстракта в водном растворителе.
[0068]
Метод получения L-фукоза α1→6-специфического лектина может предусматривать дополнительную эффективную очистку с применением любого из следующих методов очистки.
[0069]
(Метод очистки 1)
Экстракт в водном растворителе, полученный в результате осуществления данного процесса, подвергается методу осаждения сульфатом аммония с целью получения лектинсодержащей фракции. Далее полученная лектинсодержащая фракция очищается с помощью гидрофобной хроматографии и обращенно-фазовой хроматографии.
[0070]
(Метод очистки 2)
Экстракт в водном растворителе, полученный в результате осуществления данного процесса, подвергается аффинной хроматографии с использованием носителя, представляющего собой тироглобулин, иммобилизованный на агарозе.
[0071]
(Метод очистки 3)
Экстракт в водном растворителе, полученный в результате осуществления данного процесса, обрабатывается методом осаждения сульфатом аммония с целью получения лектинсодержащей фракции, которая подвергается диализу в отличие от методов дистилляции и лиофильной сушки. Далее грубая фракция лектина растворяется в растворе трис-буфера и затем подвергается ионообменной хроматографии. Далее, оставшаяся активная фракция концентрируется и в дальнейшем отделяется путем гель-фильтрации.
[0072]
Метод получения, описанный в настоящем изобретении, может включать этап обработки диализом фракции, включающей лектин, полученный путем очистки, и этап лиофилизации раствора лектина, полученного в результате диализной обработки. В итоге обеспечивается возможность простого выделения лектина. Этап обработки диализом и этап лиофилизации могут выполняться с применением традиционно используемых известных методов.
[0073]
L-фукоза α1→6-специфичный лектин, который представляет собой (a) белок или пептид с аминокислотной последовательностью, выбранной из SEQ ID NO: 2-5 или (b) белок или пептид, в котором одна или несколько аминокислот удалены, добавлены или замещены в одной из последовательностей, выбранной из SEQ ID NO: 2-5 и который функционально эквивалентен белку или пептиду с аминокислотной последовательностью, выбранной из SEQ ID NO: 2-5, может быть получен не только экстракцией из натуральных растений, но также и путем искусственной экспрессии в носителе неприродного происхождения или химического синтеза. Описанное выше вещество также находится в рамках технического объема настоящего изобретения. Экспрессия в носителе и химический синтез могут осуществляться с помощью традиционно используемых известных методов.
[0074] Настоящее изобретение позволяет также создать метод обнаружения L-фукоза α1→6 сахарной цепи с помощью L-фукоза α1→6-специфичного лектина. L-фукоза α1→6 специфичный лектин распознает L-фукоза α1→6 сахарную цепь с большей специфичностью, чем в обычном случае и может связываться с ней. Таким образом, L-фукоза α1→6-специфичный лектин желательно использовать для специфического обнаружения соединения с сахарной цепью, включая L-фукоза α1→6 сахарную цепь, например, полисахаридов, гликолипидов или гликопротеинов.
[0075] В L-фукоза α1→6-специфичном лектине, применяемом для обнаружения, предпочтительно использовать меченый лектин. Меченый лектин, описываемый в настоящем изобретении, включает, как минимум, L-фукоза α1→6-специфичный лектин и средство мечения и помечен так, что меченый лектин поддается обнаружению.
[0076] Набор средств мечения не ограничен, они могут относиться к любому известному методу мечения, в том числе, например, мечению радиоизотопом или, например, связыванию соединения-метки.
[0077] Набор соединений-меток практически не ограничен и может включать соединение, традиционно применяемое для этой цели, в том числе прямое и непрямое соединение-метку, фермент или флуоресцирующее соединение. Среди конкретных примеров соединения-метки биотин, дигоксигенин, пероксидаза из хрена, флуоресцеинтиоизоцианат и CyDye. Эти соединения-метки связываются с лектином с помощью традиционного метода.
[0078] Желательно, чтобы L-фукоза α1→6-специфичный лектин представлял собой лектин из базидиального гриба, предпочтительно PTL, SRL, NSL, LSL и AML и более предпочтительно PTL и SRL. Как показано в примере, PTL и SRL отличаются от обычных L-фукоза-специфичных лектинов тем, что PTL и SRL не связываются с L-фукозой, отличной от α1→6 связанной L-фукозы, и высокоманнозной сахарной цепью, не содержащей L-фукозы. Таким образом, PTL и SRL - оптимальный вариант при использовании L-фукоза α1→6-специфичного лектина в методе определения, описываемом в настоящем изобретении.
[0079] Обнаружение L-фукоза α1→6 сахарной цепи может быть выполнено, например, с помощью лектиновой хроматографии при использовании иммобилизованного L-фукоза α1→6-специфичного лектина. Лектиновая хроматография представляет собой аффинную хроматографию, в которой используется свойство лектина специфически связываться с сахарной цепью. В случае сочетания лектиновой хроматографии с ВЭЖХ (HPLAC) можно ожидать анализа с высокой пропускной способностью.
[0080]
Носитель, на котором иммобилизуется L-фукоза α1→6-специфичный лектин, представляет собой гелеобразующий материал, например, агарозу, декстран, целлюлозу, крахмал или полиакриламид. Эти гелеобразующие материалы могут представлять собой коммерческие материалы без каких-либо специфических ограничений, включая, например, сефарозу 4B и сефарозу 6B (GE Healthcare Bioscience).
[0081]
Колонка, используемая в лектиновой хроматографии, представляет собой колонку, в которой лектин иммобилизуется, например, на микропланшете или наномембране.
[0082]
Иммобилизованный L-фукоза α1→6-специфичный лектин имеет концентрацию, как правило, в пределах от 0,001 до 100 мг/мл и предпочтительно от 0,01 до 20 мг/мл. Когда носителем является агарозный гель, носитель активируется, например, CNBr и в дальнейшем связывается с лектином. Лектин также можно иммобилизовать на геле с помощью активированного спейсера. Или лектин можно также иммобилизовать на геле с помощью формильной группы с последующим восстановлением NaCNBH3. Или возможно также использование коммерческого активированного геля, например, NHS-сефарозы (GE Healthcare Bioscience).
[0083]
Образец L-фукоза α1→6 сахарной цепи заливается в колонку, через которую затем пропускают буферный раствор для очистки и установления равновесия. Один из видов буферного раствора имеет концентрацию от 5 до 500 ммоль, предпочтительно от 10 до 500 ммоль, и pH от 4,0 до 10,0, предпочтительно от 6,0 до 9,0, и содержит NaCl от 0 до 0,5 моль, предпочтительно от 0,1 до 0,2 моль, и CaCl2, MgCl2 или MnCl2 от 0 до 10 ммоль, предпочтительно от 0 до 5 ммоль.
[0084]
После промывки несвязанных материалов буфером L-фукоза α1→6 сахарная цепь элюируется нейтральным неденатурирующим буферным раствором, который способствует эффективному элюированию сахарной цепи десорбирующим агентом, таким, например, как хлорид натрия или гаптен-сахар. Этот буферный раствор также может представлять собой буферный раствор, указанный выше. Десорбирующий агент имеет концентрацию предпочтительно от 1 до 500 ммоль и более предпочтительно от 10 до 200 ммоль.
[0085]
Наряду с вышеописанным методом сахарная цепь обнаруживается также с помощью хроматографии, лектинового чипа, иммуноферментного анализа (ELISA), аггрегации, метода поверхностного плазменного резонанса, например, с помощью системы Biacore®, или электрофореза, например, метода, хорошо известного специалистам в данной области техники.
[0086]
На образец, включающий сахарную цепь, не накладываются какие-либо специфические ограничения. Образец с сахарной цепью может включать, например, кровь, плазму крови, сыворотку крови, слезы, жидкость организма, жидкость груди, мочу, кондиционированную среду в клеточной культуре и секрецию генетически измененных животных.
[0087]
Конкретным примером L-фукоза α1→6 сахарной цепи как объекта обнаружения данным методом является сахарная цепь, синтезированная с помощью α1→6 фукозилтрансферазы (FUT8). L-фукоза α1→6 сахарная цепь может находиться, например, в α-фетопротеине, α5β1-интегрине, TGFβ-рецепторе или EGF-рецепторе. Предпочтительно, чтобы сахарная цепь, используемая в методе обнаружения, представляла собой онкомаркер.
[0088]
Точное обнаружение α1→6 фукозилированного α-фетопротеина полезно, например, для ранней диагностики рака клеток печени, который клинически осложняет цирроз, гистологического наблюдения за раком клеток печени, точного определения терапевтического эффекта, раннего обнаружения эмбриональной опухоли и определения показателя восстановления печени при фульминантном гепатите. α5β1-интегрин, к которому переносится α1→6 связанная L-фукоза, также рассматривается как показатель при диагностике рака печени.
[0089]
Объектом метода обнаружения, описываемого в настоящем изобретении, являются в том числе, в дополнение к раку клеток печени, диагностика опухолей (например, рак предстательной железы, рак груди, рак желудка, рак тонкого кишечника, рак толстой кишки, колоректальный рак, рак почки, рак поджелудочной железы, мелкоклеточный рак легкого, немелкоклеточный рак легкого, рак матки, рак яичника, саркома мягких тканей, рак костей, меланома, глиобластомы, астроцитома, медуллобластома, острая лимфома, злокачественная лимфома, болезнь Ходжкинса, неходжкинская лимфома, острый миелолейкоз, хронический лимфолейкоз), аллергия, аутоиммунные нарушения и сердечно-сосудистые заболевания, например, эмфизема.
[0090]
L-фукоза α1→6-специфичный лектин - новый лектин, отличающийся от известного традиционного лектина своими физико-химическими и биохимическими свойствами, такими как, например, специфическое связывание с сахарной цепью. В частности, поскольку L-фукоза α1→6-специфичный лектин обладает способностью специфического распознавания L-фукоза α1→6 связи, L-фукоза α1→6-специфичный лектин может использоваться в качестве диагностического средства, тест-реагента, адсорбирующего агента для анализа с разделением карбогидратов и иммунорегулирующего средства. Таким образом, настоящее изобретение позволяет получить диагностическое средство, включающее L-фукоза α1-6-специфичный лектин в качестве активного компонента и используемое для обнаружения L-фукоза α1→6 сахарной цепи, синтезированной с помощью L-фукоза α1→6 трансферазы, и диагностический комплект, включающий диагностическое средство. Диагностическое средство или диагностический комплект используются, например, для диагностики рака клеток печени.
[0091]
Настоящее изобретение позволяет также создать метод фракционирования L-фукоза α1→6 сахарной цепи, включающий этап применения L-фукоза α1→6-специфичного лектина в качестве связывающей среды для L-фукоза α1→6 сахарной цепи с целью фракционирования L-фукоза α1→6 сахарной цепи и не содержащей L-фукозы α1→6 сахарной цепи.
[0092]
Предпочтительно, чтобы в качестве L-фукоза α1→6-специфичного лектина, используемого в методе фракционирования L-фукоза α1→6 сахарной цепи, применялся лектин из базидиального гриба, более предпочтительно PTL, SRL, NSL, LSL и AML, и более предпочтительно PTL и SRL.
[0093]
Метод фракционирования, описываемый в настоящем изобретении, представляет собой, например, лектиновую хроматографию с использованием иммобилизованного L-фукоза α1→6-специфичного лектина. Подробности метода совпадают с подробностями, описанными для метода обнаружения. Если L-фукоза α1→6 сахарная цепь очищена, L-фукоза α1→6-специфичный лектин связывается с носителем в колонке, представляющим собой агарозу или целлюлозу, с помощью функциональной группы, после чего заливается образец с сахарной цепью. После пропускания образца через колонку собирается адсорбированная L-фукоза α1→6 сахарная цепь. Если очищена не содержащая L-фукозы α1→6 сахарная цепь, собирается такой образец сахарной цепи, который не адсорбируется во время пропускания образца с сахарной цепью через колонку.
[0094]
Объект, очищенный методом фракционирования, описываемым в настоящем изобретении, может быть двух типов - L-фукоза α1→6 сахарная цепь и не содержащая L-фукозы α1→6 сахарная цепь. Конкретным примером является сахарная цепь антитела и предпочтительно сахарная цепь IgG человека.
[0095]
Чистота сахарной цепи, фракционированной с помощью метода фракционирования (т.е. соотношение в случае L-фукоза α1→6 сахарной цепи, L-фукоза α1→6 сахарной цепи и общего количества L-фукоза α1→6 сахарной цепи и не содержащей L-фукозы α1→6 сахарной цепи и соотношение, в случае не содержащей L-фукозы α1→6 сахарной цепи, не содержащей L-фукозы α1→6 сахарной цепи и общего количества L-фукоза α1→6 сахарной цепи и не содержащей L-фукозы α1→6 сахарной цепи), составляет, как правило, от 90 до 100%, предпочтительно от 95 до 100% и особенно предпочтительно от 99 до 100%.
[0096]
Настоящее изобретение позволяет также получить препарат на основе антитела, включающий в качестве активного компонента L-фукоза α1→6 сахарную цепь или не содержащую L-фукозы α1→6 сахарную цепь с чистотой, как правило, от 90 до 100%, предпочтительно от 95 до 100% и особенно предпочтительно - от 99 до 100%. В частности, препарат на основе антитела, включающий антитело, удаляемое при переносе к нему α1→6 L-фукозы, как ожидают, обеспечит повышенную ADCC-активность. Среди вариантов: Ритуксан (химерное антитело, неходжкинская лимфома), Герцептин (гуманизированное антитело, рак груди), Эрбитукс (химерное антитело, рак толстой кишки, рак головы и шеи), Зевалин (мышиное антитело, неходжкинская лимфома), Кампат (гуманизированное антитело, B-клеточный хронический лимфолейкоз), Бексар (мышиное антитело, неходжкинская лимфома) и Авастин (гуманизированное антитело, метастатический рак толстой кишки).
[0097]
Медицинское средство на основе антитела, полученное методом фракционирования, описываемым в настоящем изобретении, может использоваться так же, как это предусматривается традиционным методом (в том, что касается, например, фармакологически одобренного носителя, вспомогательного вещества, способа введения лекарства или лекарственной формы), за исключением того факта, что медицинское средство на основе антитела, полученное методом фракционирования, описываемым в настоящем изобретении, может использоваться в низкодозовом режиме и в малой дозировке, поскольку это средство обладает более высокой специфической активностью, чем традиционное средство.
[0098]
Настоящее изобретение позволяет также получить L-фукоза α1→6 сахарную цепь или не содержащую L-фукозы α1→6 сахарную цепь с чистотой от 90 до 100%, фракционированную предложенным методом, а также медицинское средство, включающее в качестве активного компонента L-фукоза α1→6 сахарную цепь или не содержащую L-фукозы α1→6 сахарную цепь с чистотой от 90 до 100%. Чистота означает, что в случае L-фукоза α1→6 сахарной цепи, соотношение L-фукоза α1→6 сахарной цепи и общего количества L-фукоза α1→6 сахарной цепи и не содержащей L-фукозы α1→6 сахарной цепи и, в случае не содержащей L-фукозы α1→6 сахарной цепи, соотношение не содержащей L-фукозы α1→6 сахарной цепи и общего количества L-фукоза α1→6 сахарной цепи и не содержащей L-фукозы α1→6 сахарной цепи. Предпочтительно, чтобы медицинское средство представляло собой препарат на основе антитела.
[0099]
Настоящее изобретение позволяет также создать метод отбора L-фукозы α1→6 сахарной цепи. Этот метод включает этап взаимодействия жидкости, содержащей сахарную цепь, с L-фукоза α1→6-специфичным лектином и сбор L-фукоза α1→6 сахарной цепи, адсорбированной L-фукоза α1→6-специфичным лектином. Этот метод отбора полезен для поиска нового онкомаркера с L-фукоза α1→6 сахарной цепью. Можно также использовать L-фукоза α1→6-специфичный лектин, который может применяться в целях метода обнаружения, описываемого в настоящем изобретении, для простого отбора маркера, включающего L-фукоза α1→6 сахарную цепь.
[0100]
Предпочтительно, чтобы L-фукоза α1→6-специфичный лектин, используемый в методе отбора, представлял собой лектин из базидиального гриба, особенно предпочтительно - одного из семейства Strophariaceae, Tricholomataceae, Polyporaceae и Amanitaceae и более предпочтительно PTL, SRL, NSL, LSL, и/или AML, среди которых самыми предпочтительными являются PTL и SRL.
[0101]
Настоящее изобретение позволяет также создать метод отбора L-фукоза α1→6 сахарной цепи специфичного лектина. В этом методе используется, в частности, иммобилизованный L-фукоза α1→6-специфичный лектин. Обеспечивается воздействие жидкости, содержащей несколько сахарных цепей, на иммобилизованный лектин. Далее тестовый профиль сахарной цепи, адсорбированной на иммобилизованном лектине, (например, гель-электрофрез) сравнивается с эталонным профилем сахарной цепи, адсорбированной лектином, в том случае, когда жидкость действует на иммобилизованные PTL или SRL. Таким образом, экстрагируется L-фукоза α1→6 специфичный лектин с профилем, совпадающим с эталонным. Поскольку идентифицировать сахарную цепь, адсорбированную на каждом образце лектина, не требуется, целевой лектин можно отбирать путем очень простой процедуры. Экстрагированный таким образом L-фукоза α1→6-специфичный лектин может использоваться в качестве эталона для следующего метода отбора. Возможно и применение другого метода для использования cDNA с целью полного или частичного кодирования аминокислотной последовательности, SEQ ID NO: 1. в качестве праймера для улавливания cDNA L-фукоза α1→6-специфичного лектина из образца лектина.
[0102]
Сахарная цепь включает, по меньшей мере, одну из L-фукоза α1→6 сахарных цепей, как показано на рис.1 и рис.2, и предпочтительно включает, по меньшей мере, одну из не содержащих L-фукозы α1→6 сахарных цепей, к которым переносится L-фукоза, отличная от присоединяемой по связи α1→6, и, по меньшей мере, одну сахарную цепь, не содержащую L-фукозы (например, высокоманнозная сахарная цепь). Путем ее добавления можно подтвердить отсутствие сродства к сахарной цепи, отличной от L-фукоза α1→6 сахарной цепи.
[0103]
Профиль адсорбированной сахарной цепи можно измерить с помощью метода, хорошо известного специалистам в данной области техники, например, с помощью различных видов хроматографии, масс-анализа, гель-электрофореза, лектинового чипа, иммуноферменного анализа (ELISA), поверхностного плазменного резонанса, например, системы Biacore®, или электрофореза.
Пример
[0104]
В следующем разделе настоящее изобретение описывается более подробно на примерах и сравнительных примерах. Однако оно не ограничивается представленными примерами.
[0105]
[Примеры 1 и 2] (Получение, измерение характеристик и описание PTL и SRL) (1)
Получение PTL (пример 1)
На основе процесса очистки, показанного на рис.3, лектин из Pholiota terrestris (PTL) был очищен из гриба Pholiota terrestris.
[0106]
(Экстракция)
Все процедуры выполнялись при 4°C. Высушенные сублимацией порошки Pholiota terrestris подвергали экстракции с помощью 50 мл 10 мМ трис-буфера (pH 7,2) при 4 градусах C в течение 2 часов. Полученную жидкость центрифугировали (15000 об./мин, 20 мин, 4 градуса C). Далее надосадочную жидкость фильтровали через марлю с получением первого экстракта. Остаток подвергался повторной экстракции 50 мл 10 мМ трис-буфера (pH 7,2) при 4 градусах C в течение ночи. Затем жидкость центрифугировали (15000 об./мин, 20 мин, 4 градуса C) и надосадочную жидкость фильтровали через марлю с получением второго экстракта. Затем эти экстракты фильтровали совместно через фильтровальную бумагу с получением экстракта Pholiota terrestris.
[0107]
(Ионообменная хроматография)
Экстракт (87 мл) заливали в колонку с ДЭАЭ-сефарозой (GE Healthcare Bioscience) и устанавливали равновесие с помощью 10 мМ трис-буфера (pH 7,2). После промывки колонки буфером, связанную фракцию десорбировали 0,1 M NaCl в буфере. Далее фракции с гемагглютинирующей активностью (показанные ←→ на рис.4) подвергались диализу против дистиллированной воды и лиофилизации.
[0108]
(Аффинная хроматография)
Лиофилизированный диализат снова растворяли в 10 мМ натрий-фосфатном буфере (pH 7,4, в дальнейшем просто PBS). Затем раствор экстракта заливали в колонку с тиреоглобулином, иммобилизованным на агарозе, причем равновесие достигалось с помощью того же буфера. После промывки колонки PBS связанная фракция десорбировалась с помощью 0,2 М аммиака. Затем фракции с гемагглютинирующей активностью (показанные ←→ на рис.5) собирали, подвергали ультрафильтрации и лиофилизации с получением 1,07 мг PTL.
[0109]
(Электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE))
Процедура SDS-PAGE выполнялась с помощью прибора для электрофореза Phastsystem (GE Healthcare Bioscience) и геля с градиентом 8-25 (GE Healthcare Bioscience). Исследуемый раствор и маркер молекулярного веса использовались в количестве 1 мкл.
Электрофорез проводили на основе протокола продукта с помощью традиционного метода. На рис.6 показаны результаты SDS-PAGE для PTL. На рис.6 полоса M, полосы 1 и 2 означают следующее. Полоса M: маркер молекулярного веса (APRO), полоса 1: PTL, 2-меркаптоэтанол (-), полоса 2: PTL, 2-меркаптоэтанол (+), гель: Градиент 8-25, Образец: 1 мкл/полоса, краситель: куммасси бриллиантовый голубой (CBB)
[0110]
При проведении SDS-PAGE с 8-25% гелем подтверждено, что основным компонентом является PTL.
[0111]
(2) Получение SRL (пример 2)
На основе процесса очистки, показанного на рис.7, лектин из Stropharia rugosoannulata (SRL) был очищен из гриба Stropharia rugosoannulata.
[0112]
(Экстракция)
Все процедуры выполнялись при 4°C. Высушенные сублимацией порошки Stropharia rugosoannulata (400 г) подвергали экстракции с помощью 1,6 л PBS при 4 градусах C в течение 2 часов. Полученную жидкость центрифугировали (15000 об./мин, 20 мин, 4 градуса C). Далее надосадочную жидкость фильтровали через марлю с получением первого экстракта. Остаток подвергался повторной экстракции 0.8 л PBS при 4 градусах C в течение ночи. Затем жидкость центрифугировали (10000 об./мин, 20 мин, 4 градуса C) и надосадочную жидкость фильтровали через марлю с получением второго экстракта. Затем эти экстракты перемешивали с получением жидкого экстракта Stropharia rugosoannulata.
[0113]
(Осаждение сульфатом аммония)
Твердый (NH4)2SO4 (1,3 кг) добавляли к полученной надосадочной жидкости (2,4 л) до 80% насыщения. После оставления на ночь при 4 градусах C осадок собирали центрифугированием (10000 об./мин, 20 мин, 4 градуса C) и подвергали диализу против дистиллированной воды и лиофилизации с получением фракции Stropharia rugosoannulata, полученной фракционированием 80% сульфатом аммония.
[0114]
(Гидрофобная хроматография)
Stropharia rugosoannulata, полученная фракционированием 80% сульфатом аммония, заливали в Butyl-TOYOPEARL 650M (TOSOH CORPORATION), равновесие в котором устанавливали с помощью 2M сульфата аммония-PBS, для очистки путем гидрофобной хроматографии. Этот вид хроматографии предусматривает сбор, ультрафильтрацию и лиофилизацию фракций, элюированных дистиллированной водой, с получением грубой фракции лектина из Stropharia rugosoannulata (показана ←→ на рис.8).
[0115]
(Обращенно-фазовая хроматография)
Грубую фракцию лектина из Stropharia rugosoarmulata заливали в колонку с C8 (Wako Pure Chemical Industries, Ltd.), равновесие в которой достигалось с помощью 0,05% трифторуксусной кислоты (TFA)/ацетонитрила (100/0). При использовании этой разновидности хроматографии собирали фракцию, элюированную 0,05% TFA/ацетонитрилом (70/30) (показана ←→ на рис.9). Далее удаляли растворитель путем испарения при комнатной температуре, а образовавшиеся сухие порошки собирали с получением 7,5 мг SRL.
[0116]
SDS-PAGE (PhastGel, градиент 8-25) проводили с помощью системы Phastsystem (GE Healthcare Bio-Sciences). Исследуемый раствор и маркер молекулярного веса брали в количестве 1 мкл. Электрофорез проводили на основе протокола продукта с помощью традиционного метода. На рис.10 показаны результаты SDS-PAGE для SRL. На рис.10 полоса M, полосы 1 и 2 означают следующее. Полоса M: маркер молекулярного веса (APRO), полоса 1: SRL, 2-меркаптоэтанол (+), полоса 2: SRL, 2-меркаптоэтанол (-), гель: градиент 8-25, Образец: 1 мкл/полоса, краситель: серебряный.
[0117]
При проведении SDS-PAGE с 8-25% гелем подтверждено, что основным компонентом является SRL.
[0118]
(3) Свойства PTL и SRL
(Масс-спектрометрический анализ MALDI-TOF)
PTL из примера 1 и SRL из примера 2 в количестве 10 мкг соответственно по отдельности растворяли в TA (смесь с объемным соотношением 0,1%-TFA и ацетонитрила 2:1). Далее насыщенный образец, растворенный в TA- и лектиновом TA-растворах, смешивали в объемном соотношении 4:1 и образовавшуюся смесь добавляли по каплям в количестве 1,0 мкл на подложку с получением образца. Для измерения молекулярного веса PTL и SRL в режиме LP использовали прибор Autoflex (Broker Daltonics K.K.). Как показывает результат, молекулярный вес составляет около 4500 m/z (рис.11 и рис.12).
[0119]
(Анализ аминокислотной последовательности)
Для PTL из примера 1 и SRL из примера 2 анализировали аминокислотную последовательность с помощью прибора Protein Peptide Sequencer PPSQ-21 (SHIMADZU CORPORATION). Результаты отмечены номерами последовательностей 2 и 3 соответственно. Все последовательности оказались новыми.
[0120]
PTL и SRL были подвергнуты тесту на агглютинирующую активность для эритроцитов кролика, лошади, свиньи, овцы, человека (A, B, O) и эритроцитов кролика, обработанных актиназой E. Результат в виде гемагглютинирующей активности показан в таблице 1.
[0121]
NT: Испытания не проводились
[0122]
Как видно из представленных выше результатов анализа аминокислотного состава и теста на гемагглютинирующую активность, PTL из примера 1 и SRL из примера 2 представляли собой новый лектин.
[0123]
(4) Оценка сахарсвязывающей специфичности PTL и SRL
Различные моносахариды, олигосахариды и полисахариды, представленные в таблице 2, и гликопротеины, представленные в таблице 3, тестировали на подавление гемагглютинации для оценки сахарсвязывающей специфичности PTL из примера 1 и SRL из примера 2.
[0124]
В 96-луночном титрационном микропланшете с U-образным дном приготовили серию двойных разведении 10 мкл растворов моносахарида, олигосахарида, полисахарида и гликопротеина. Затем раствор лектина с предварительно установленным титром 4 добавляли в количестве 10 мкл в соответствующие лунки. Далее планшет выдерживали для сенсибилизации при комнатной температуре в течение одного часа. После этого 10 мкл 4% суспензии эритроцитов добавляли в соответствующие лунки и выдерживали планшет при комнатной температуре в течение часа. Далее визуально оценивался фактор разведения исследуемого раствора, при котором полностью подавлялась гемагглютинация. Минимальная концентрация, при которой наблюдалось подавление, получила название минимальной подавляющей концентрации. Чем ниже минимальная подавляющая концентрация, тем выше специфичность к лектину. Результаты представлены в таблицах 2 и 3.
[0125]
Для сравнения оценивалась сахарсвязывающая специфичность с использованием следующих коммерчески доступных лектинов: сравнительный пример 1: AAL (SEIKAGAKU BIOBUSINESS CORPORATION - J-OIL MILLS, Inc.), сравнительный пример 2: AOL (TOKYO CHEMICAL INDUSTRY CO., LTD. - Gekkeikan Sake Company, Ltd.), сравнительный пример 3: LCA (SEIKAGAKU BIOBUSINESS CORPORATION - J-OIL MILLS, Inc.) и сравнительный пример 4: PSA (SEIKAGAKU BIOBUSINESS CORPORATION - J-OIL MILLS, Inc.). Результаты представлены в таблицах 2 и 3.
[0126]
[0127]
[0128]
Как видно из таблиц 2 и 3, PTL из примера 1 и SRL из примера 2 связывались только с тиреоглобулином с α1→6 связанной L-фукозой. С другой стороны, AAL из сравнительного примера 1 связывался не только с тиреоглобулином, но также с сахарами, такими как L-фукоза, фруктоза, а также с гликопротеином, например, муцином с L-фукозой в O-сцепленной сахарной цепи. AOL из сравнительного примера 2 связывался с сахарами, такими как L-фукоза и фруктоза, и с гликопротеином, например, муцином с L-фукозой в O-сцепленной сахарной цепи. LCA из сравнительного примера 3 и PSA из сравнительного примера 4 связывались не только с тиреоглобулином, но также с сахарами, такими как манноза и метил α-маннозид. Можно сказать, что PTL из примера 1 и SRL из примера 2 представляют собой L-фукоза α1→6 специфичный лектин, который не связывается с L-фукозой и маннозой, а связывается только с α1→6 связанной L-фукозой.
[0129]
(5) Измерение констант ассоциации PTL и SRL с L-фукоза α1→6 сахарной цепью. Константы ассоциации PTL из примера 1 и SRL из примера 2 с L-фукоза α1→6 сахарной цепью измерялись с помощью следующей процедуры.
[0130]
(Приготовление олигосахарида)
Пиридиламинированные (PA) сахарные цепи, показанные на рис.1 и 2, использовались для анализа методом фронтальной аффинной хроматографии (FAC). PA-сахара были приобретены у компаний TAKARA BIO INC., SEIKAGAKU BIOBUSINESS CORPORATION и Masuda ChemicalIndustries co., LTD. PA-caxap получали также путем пиридиламинирования с помощью GlyeoTAG® (TAKARA BIO INC.) немаркированной сахарной цепи или сахарной цепи, полученной, например, ферментативным расщеплением.
[0131]
(Подготовка колонки с лектином)
Лектин растворяли в 0.2 M буферном растворе NaHCO3 (pH 8,3), включающем 0,5 М NaCl, и связывали с NHS-активированной сефарозой (GE Healthcare Bioscience) согласно указаниям производителя. Затем сефарозу с иммобилизованным лектином суспендировали в 10 мМ трис-буфере (pH 7,4, TBS), включающем 0,8%-NaCl, и полученным веществом заполняли миниатюрную колонку (φ2 мм× 10 мм, 31,4 мкл).
[0132]
(Фронтальная аффинная хроматография)
Фронтальная аффинная хроматография выполнялась с помощью автоматического прибора FAC (FAC-1, SHIMADZU CORPORATION). В частности, подготовленная вышеописанным способом колонка с лектином устанавливалась в держателе из нержавеющей стали, а держатель прикрепляли к прибору FAC-1. Скорость потока и температура колонки поддерживались на уровне 0,125 мл/мин и 25 градусов C соответственно. После установления равновесия в колонке с помощью TBS излишний объем (от 0,5 мл до 4 мл) PA-сахарной цепи (3,75 нМ или 7,5 нМ) непрерывно подавали в колонку с помощью автоматического прибора для взятия проб.
[0133]
Интенсивность флуоресценции элюата PA-сахара (длина волны возбуждения 310 нм и длина волны флуоресцентного излучения 380 нм) регистрировалась для измерения интенсивности взаимодействия [разница между передним краем элюата и стандартным полисахаридом (PA-рамноза): V-V0]. Исходя из интенсивности взаимодействия и эффективного количества лиганда рассчитывалась константа ассоциации Ka. Результат представлен в таблицах 4-9.
[0134]
Для сравнения, с помощью процедуры, аналогичной вышеописанной, рассчитывались также константы ассоциации для AAL (сравнительный пример 1), AOL (сравнительный пример 2), LCA (сравнительный пример 3) и PSA (сравнительный пример 4), которые считаются лектинами, специфичными к коровой фукозе. Результаты показаны в таблицах 4-7.
[0135]
[0136]
[0137]
[0138]
[0139]
Как видно из таблиц 4-7, AAL из сравнительного примера 1 и AOL из сравнительного примера 2 связываются с не содержащими α1→6 гликолипидную L-фукозу сахарными цепями (сахарные цепи 718, 722, 723, 727, 909, 910 и 933), а также с L-фукоза α1→6 сахарными цепями (сахарные цепи 15, 201-203 и 401-418). LCA из сравнительного примера 3 и PSA из сравнительного примера 4 связываются с большим количеством сахарных цепей, не содержащих L-фукоза α1→6 сахарных цепей (сахарные цепи 003, 005-014). С другой стороны, PTL из примера 1 и SRL из примера 2 крепко связываются с L-фукоза α1→6 сахарной цепью и не связываются с не содержащей L-фукозы α1→6 сахарной цепью и сахарной цепью, вообще не содержащей L-фукозы. Кроме того, PTL из примера 1 имеет более высокую константу ассоциации, чем обычный лектин (константа ассоциации Ka=1.0×105M-1или более). Более того, PTL из примера 1 и SRL из примера 2 также крепко связываются с корово фукозилированными триантенными N-гликанами (сахарные цепи 407-413) и тетраантенным N-гликаном (сахарная цепь 418). Даже в случае добавления сиаловой кислоты (сахарные цепи 601 и 602) видно, что константа ассоциации L-фукоза α1→6 сахарной цепи не понижается.
[0140]
(6) Обнаружение гликопротеина с помощью PTL и SRL
(i) Приготовление гликопротеина
Были приготовлены гликопротеины (1)-(9) со следующими структурами главной сахарной цепи и (10) альбумин бычьей сыворотки, не содержащий сахара.
[0141]
(1) В качестве гликопротеина, содержащего L-фукоза α1→6 N-связанные гликаны, тиреоглобулин (свиной), имеющий следующую формулу:
[Химическая формула 3]
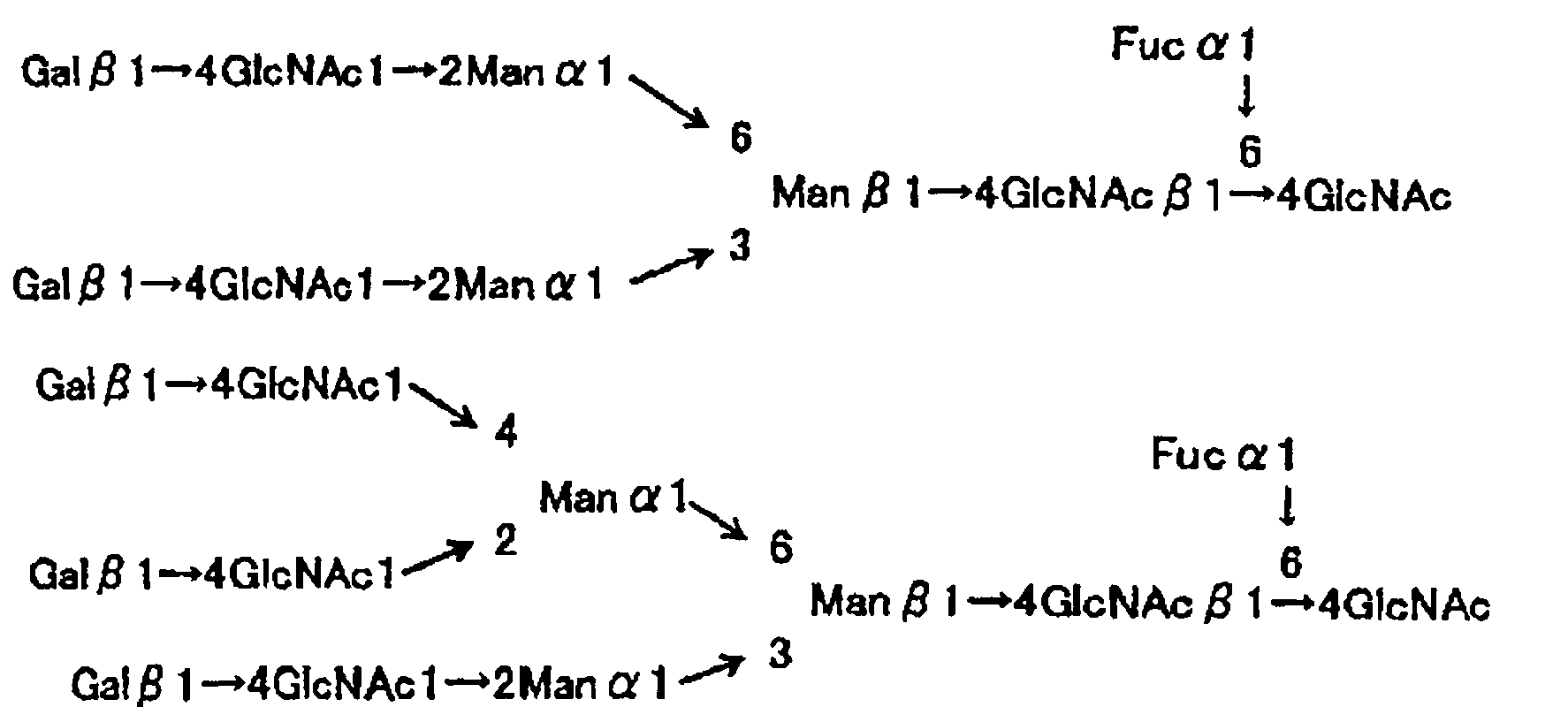
[0142]
(2) В качестве гликопротеина, содержащего L-фукоза α1→6 N-связанные гликаны, лактоферрин (бычий), имеющий следующую формулу:
[Химическая формула 4]

[0143]
(3) В качестве гликопротеина, содержащего L-фукоза α1→6 N-связанные гликаны, иммунный глобулин G (человеческий), имеющий следующую формулу:
[Химическая формула 5]
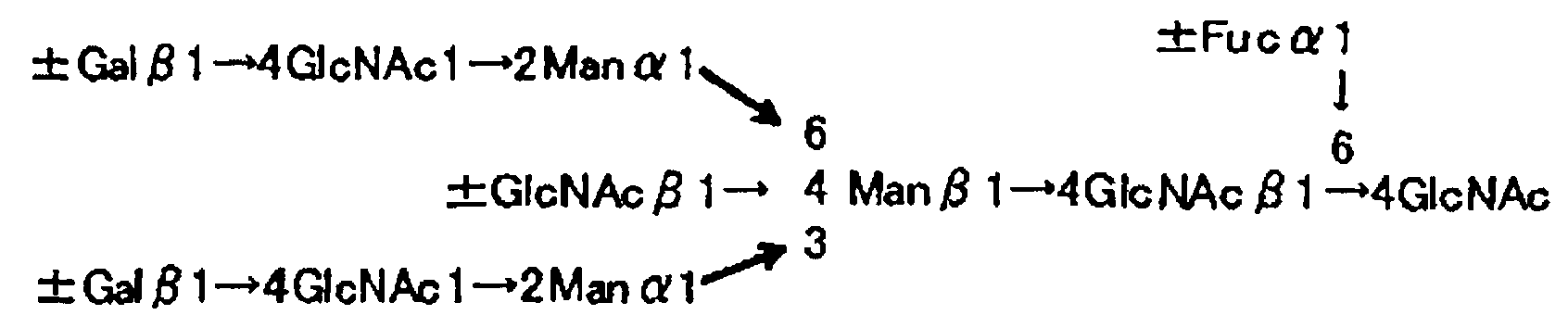
[0144]
(4) В качестве гликопротеина, не содержащего L-фукоза α1→6 N-связанных гликанов, трансферрин (человеческий), имеющий следующую формулу:
[Химическая формула 6]
[0145]
(5) В качестве гликопротеина, не содержащего L-фукоза α1→6 N-связанных гликанов, α1-кислотный гликопротеин (человеческий), имеющий следующую формулу:
[Химическая формула 7]


[0146]
(6) В качестве гликопротеина, содержащего высокоманнозные сахарные цепи, инвертаза (дрожжевая), имеющая следующую формулу:
[Химическая формула 8]

[0147]
(7) В качестве гликопротеина, содержащего L-фукоза O-сцепленные сахарные цепи, муцин (свиной), имеющий следующую формулу:
[Химическая формула 9]
[0148]
(8) В качестве гликопротеина, содержащего L-фукоза O-сцепленные сахарные цепи, муцин (бычий), имеющий следующую формулу:
[Химическая формула 10]
[0149]
(ii) Обнаружение гликопротеина с помощью вестерн-блоттинга
PTL из примера 1, SRL из примера 2, AAL из сравнительного примера 1, AOL из сравнительного примера 2 и LCA из сравнительного примера 3 подвергались биотиновому мечению.
[0150]
(Биотинилирование лектина)
Лектин измеряли и растворяли в 0,1 M растворе бикарбоната натрия (5 мг/мл). Затем реагент для биотинилирования растворяли в диметилсульфоксиде, и полученный раствор добавляли к раствору лектина для проведения реакции между ними. Далее реактант дистиллировали и лиофилизировали для получения меченного биотином лектина.
[0151]
(SDS-PAGE и блоттинг гликопротеина)
Раствор, полученный растворением образца гликопротеина в 10 мМ натрий-фосфатном буфере (pH 7,4, PBS) при 2 мг/мл, подавался в микротрубку по 18 мкл. 6 мкл жидкого додецилсульфата натрия (SDS) (исследуемый буферный раствор (2МЕ-); Wako Pure Chemical Industries, Ltd.) и 1,25 мкл 2-меркаптоэтанола (Bio-Rad Laboratories, Inc.) добавляли к каждой жидкости и полученную смесь кипятили в течение 5 минут.
После электрофореза в полиакриламидном геле вещества переносили на ПВДФ-мембраны (Immobilon IPVH 304 F0, Millipore K.K.).
[0152]
(Окрашивание меченного биотином лектина)
Пленку погружали в 10 мМ раствор трис-буфера, содержащего 0,8% NaCl с 1% BSA (pH 7,4, 1% BSA + TBS), и встряхивали при комнатной температуре в течение одного часа. Затем пленку очищали три раза с помощью 10 мМ раствора трис-буфера (pH 7,4, TBS), содержащего 0,8% NaCl. Затем пленку погрузили в раствор меченного биотином лектина (2 мкг/мл) и встряхивали при комнатной температуре в течение одного часа. Затем пленку три раза очищали с помощью TBS. После этого пленку погружали в раствор меченного HRP (пероксидазой хрена) стрептавидина при 1 мкг/мл (Vector Laboratories) и встряхивали при комнатной температуре в течение 30 минут. После того, как пленка была очищена три раза с помощью TBS, для выполнения пробы с окрашиванием использовали набор для окраски с использованием иммунной метки POD (Wako Pure Chemical Industries, Ltd.). Проба с окрашиванием предназначена для обнаружения гликопротеина с помощью вестерн-блоттинга с использованием меченного биотином лектина. На рис.13-17 представлены фотографии, на которых изображены следующие гликопротеины, окрашенные с помощью PTL (рис.13), SRL (рис.14), AAL (рис.15), AOL (рис.16) или LCA (рис.17).
[0153]
На рис.13-17 полосы 0-6 означают следующее. Полоса 1: тиреоглобулин, полоса 2: лактоферрин, полоса 3: иммунный иммуноглобулин, полоса 4: трансферрин, полоса 5: α1-кислотный гликопротеин, полоса 6: инвертаза, полоса 0: альбумин бычьей сыворотки (станжарт).
[0154]
Для контроля выполнили также окрашивание белка СВВ. Фотография окрашивания представлена на рис.18. На рис.18 полосы M и 1-6 означают следующее. Полоса M: маркер молекулярного веса, полоса 1: тиреоглобулин, полоса 2: лактоферрин, полоса 3: иммуноглобулин G, полоса 4: трансферрин, полоса 5: α1-кислотный гликопротеин, полоса 6: инвертаза, полоса 0: альбумин бычьей сыворотки
[0155]
[0156]
Как видно из результатов, представленных в таблице 8 и на рис.13-18, в случае LCA из сравнительного примера
обнаруживался не только гликопротеин, содержащий L-фукоза α1→6 сахарные цепи, но и гликопротеин, содержащий высокоманнозную цепь (инвертаза). Напротив, в случае PTL из примера 1 и SRL из примера 2 обнаруживался только гликопротеин, содержащий sL-фукоза α1→6 сахарную цепь, а не содержащие L-фукозы α1→6 сахарные цепи и сахарные цепи, не содержащие L-фукозы, не обнаруживались совсем.
[0157]
(iii) Обнаружение гликопротеинов с помощью иммуноферментного анализа (ELISA)
PTL из примера 1, SRL из примера 2, AAL из сравнительного примера 1, AOL из сравнительного примера 2 и LCA из сравнительного примера 3, меченные биотином, использовались для обнаружения гликопротеинов методом иммуноферментного анализа (ELISA).
[0158]
Гликопротеин и альбумин в качестве белка, не содержащего сахарную цепь и выполняющего роль стандарта, растворяли в 0,1 М буферном растворе углекислоты (pH 9,5) при 1 мг/мл. Далее полученный раствор добавляли на титрационный микропланшет (Nunc 439454) и инкубировали при 4 градусах C в течение ночи. Затем раствор очищали три раза с помощью 0.05% Tween/PBS. После этого добавляли в лунки 1% BSA/PBS и инкубировали раствор при 37 градусах C в течение одного часа. Далее после трехкратной очистки раствора 0,05% Tween/PBS добавляли в лунки меченный биотином лектин, разведенный соответствующим образом с помощью 1% BSA/0.05% Tween/PBS и инкубировали раствор при 37 градусах C в течение одного часа. После трехкратной очистки раствора 0,05% Tween/PBS добавляли в лунки раствор меченного HRP (пероксидазой хрена) стрептавидина, разведенный 1% BSA/0.05% Tween/PBS и инкубировали раствор при 37 градусах C в течение 30 минут. После трехкратной очистки раствора 0,05% Tween/PBS добавляли субстрат ТМБ-Перокисдаза (KPL) и полученный раствор инкубировали при комнатной температуре и без доступа света в течение 10 минут.
[0159]
Реакции останавливали с помощью 1 M фосфорной кислоты. Затем с использованием считывающего устройства для микропланшетов (MPR-A4i, TOSOH CORPORATION) измеряли поглощение при 450 нм. На основе этого значения рассчитывали показатель для реакции ([поглощение при 450 нм для лунки, в которой находился твердофазный гликопротеин, участвовавший в реакции] - [поглощение при 450 нм для лунки, в которой не было твердофазного гликопротеина, участвовавшего в реакции]. Далее для каждого лектина рассчитывали интенсивность взаимодействия (относительное значение) с каждым гликопротеином на основе допущения о том, что значение для гликопротеина (тиреоглобулин) составляет 100%. Результаты расчетов представлены в таблице 9 и на рис.19-23.
[0160]

[0161]
Как видно из таблицы 9 и рис.19-23, в случае AAL из сравнительного примера 1 и AOL из сравнительного примера 2 обнаруживался не только гликопротеин, содержащий L-фукоза α1→6 сахарные цепи, но и, что является нежелательным, O-сцепленные сахарные цепи (муцин).
Напротив, в случае PTL из примера 1 и SRL из примера 2 обнаруживался только гликопротеин, содержащий L-фукоза α1→6 сахарные цепи, а не содержащие L-фукозы α1→6 сахарные цепи и сахарные цепи, не содержащие L-фукозы, не обнаруживались.
Кроме того, лектины из сравнительных примеров 1-3 демонстрируют интенсивность взаимодействия с лактоферрином и иммуноглобулином G (IgG), более низкую по сравнению с той, что наблюдается для тиреоглобулина. Напротив, лектины из примеров 1 и 2 продемонстрировали такую же интенсивность взаимодействия с лактоферрином, что и в случае тиреоглобулина.
[0162]
(iv) Обнаружение сахарной цепи онкомаркера с помощью метода ELISA
α-фетопротеин (далее "AFP") представляет собой гликопротеин, содержащийся в сыворотке крови с N-сшитой сахарной цепью. AFP практически отсутствует в сыворотке крови здорового взрослого человека. С другой стороны, сыворотка крови пациента с доброкачественным заболеванием печени характеризуется повышенным содержанием α-фетопротеин L1-сахарной цепи(AFP-L1).
Далее α-фетопротеин L3-сахарная цепь (AFP-L3) обнаружена у пациента, страдающего раком печени. Обычно различие в сахарных цепях измеряется с помощью LCA, и результаты измерений используются для диагностики заболеваний печени.
[0163]
[Химическая формула 12]
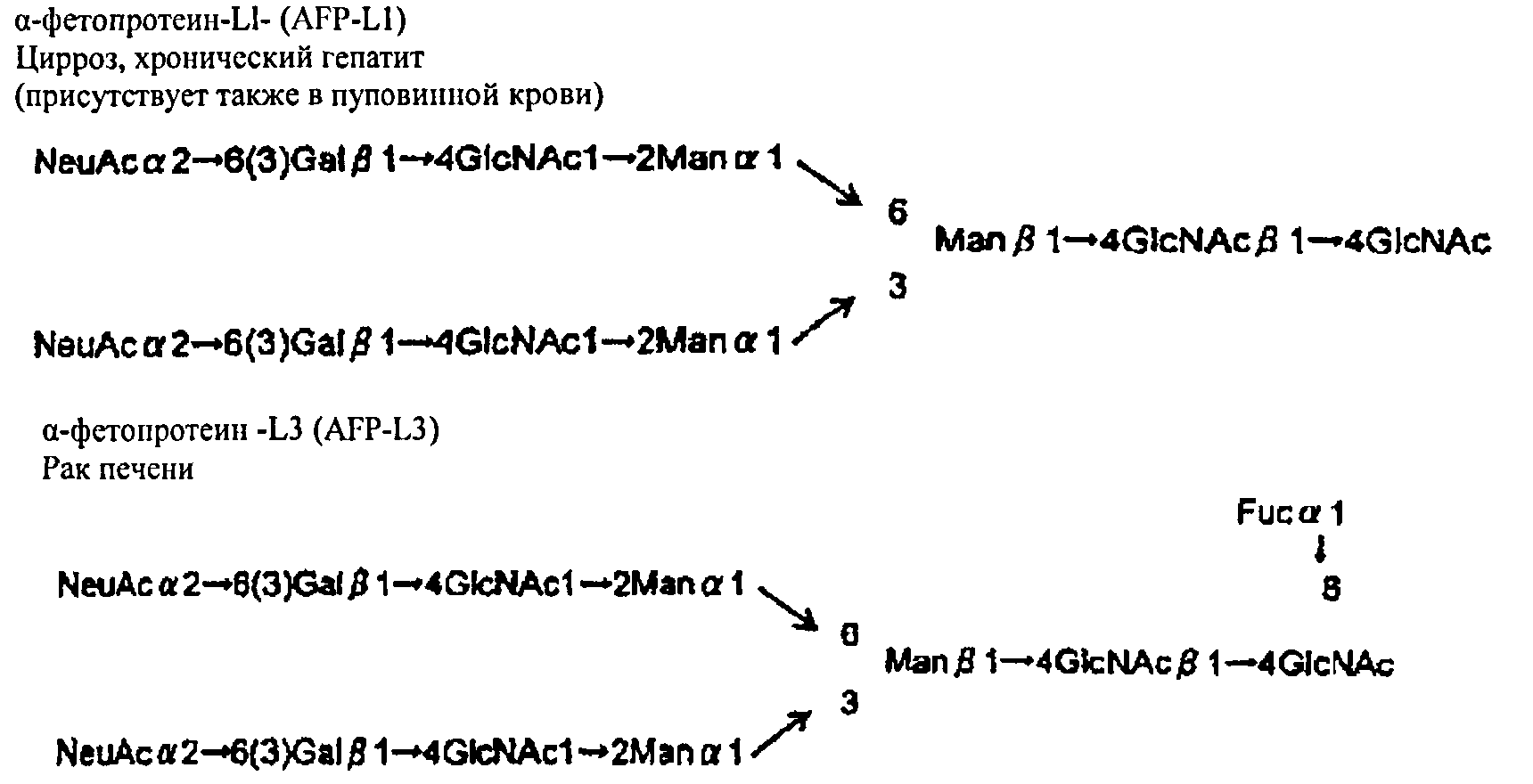
[0164]
Меченные биотином PTL из примера 1, SRL из примера 2, AAL из примера 1, AOL из сравнительного примера 2 и LCA из сравнительного примера 3 использовались для оценки связывающей способности α-фетопротеина по методу ELISA.
[0165]
0.1 M буферный раствор углекислоты (pH 9,5) использовали для разведения α-фетопротеина (полученного из сыворотки пуповинной крови человека, главным образом L1-сахарная цепь) и α-фетопротеина-L3 (приготовленного из кондиционированной среды клеток рака печени человека) до 0,01 мкг/мл. Далее полученный раствор добавляли на микротитрационный планшет (Nunc) и инкубировали при 4 градусах C в течение одной ночи. После трехкратной очистки раствора 0,05% Tween/PBS добавляли в лунки 1% BSA/PBS и инкубировали раствор при 37 градусах C в течение одного часа. После трехкратной промывки раствора 0,05% Tween/PBS в лунки добавляли раствор меченного биотином лектина, соответствующим образом разведенный 1% BSA/0.05% Tween/PBS, и инкубировали раствор при 37 градусах C в течение одного часа. После трехкратной очистки раствора 0.05% Tween/PBS добавляли раствор меченного HRP стрептавидина, разведенный 1% BSA/0.05% Tween/PBS, и инкубировали раствор при 37 градусах C в течение 30 минут. После трехкратной очистки раствора 0,05% Tween/PBS, добавляли субстрат ТМВ-Пероксидаза (KPL) и полученный раствор инкубировали при комнатной температуре и без доступа света в течение 10 минут. Реакцию останавливали 1 M фосфорной кислотой. Далее с помощью считывающего устройства для микропланшетов (MPR-A4i, TOSOH CORPORATION) измеряли поглощение при 450 нм.
[0166]
Измеряли поглощение при 450 нм для планшета, на котором происходила реакция α-фетопротеина и α-фетопротеина-L3 с лектином. Исходя из этого значения рассчитывали значение для реакции ([поглощение при 450 нм для лунки, в которой находился твердофазный гликопротеин, участвовавший в реакции] - [поглощение при 450 нм для лунки, в которой не было твердофазного гликопротеина, участвовавшего в реакции]. Далее для каждого лектина рассчитывали интенсивность взаимодействия. Результаты расчетов представлены в таблице 10 и на рис.24-28.
[0167]
[0168]
Как видно из таблицы 10, PTL из примера 1 и SRL из примера 2 позволяют обнаружить изменение в сахарной цепи онкомаркера (α-фетопротеин-L1 - сахарная цепь и L3-сахарная цепь) с уровнем точности, аналогичным или более высоким, чем в случае обнаружения по LCA из сравнительного примера 3. Таким образом, PTL из примера 1 и SRL из примера 2 могут использоваться в качестве диагностического агента или в составе диагностического комплекта для обнаружения изменений в сахарной цепи онкомаркера. Кроме того, лектин, описываемый в настоящем изобретении, обладает высокой специфичностью и позволяет обнаруживать соединения с L-фукоза α1→6 сахарной цепью среди группы соединений, содержащих сахарные цепи, отличные от L-фукоза α1→6 сахарной цепи, с более высокой точностью, чем другие лектины.
[0169]
[Примеры 3 и 4] (Получение и измерение свойств NSL и LSL)
(1) Получение NSL (пример 3)
С использованием процесса очистки, представленного на рис.29, лектин Naematoloma sublateritium (NSL) был очищен от Naematoloma sublateritium.
[0170]
(Экстракция)
Высушенные сублимацией порошки Naematoloma sublateritium (40 г) подвергали экстракции с помощью 0,8 л PBS при 4 градусах C в течение 2 часов. Полученную жидкость центрифугировали (10000 об./мин, 20 мин, 4 градуса C). Далее надосадочную жидкость фильтровали через марлю с получением первого экстракта. Остаток подвергался повторной экстракции 0,4 л PBS при 4 градусах C в течение ночи. Затем жидкость центрифугировали (10000 об./мин, 20 мин, 4 градуса C) и надосадочную жидкость фильтровали через марлю с получением второго экстракта. Затем экстракты перемешивали с получением жидкого экстракта Naematoloma sublateritium.
[0171]
(Осаждение сульфатом аммония)
Твердый (NH4)2SO4 (0,8 кг) добавляли к полученной надосадочной жидкости (1,5 л) до 80% насыщения. После оставления на ночь при 4 градусах C осадок собирался центрифугированием (10000 об./мин, 20 мин, 4 градуса C) и подвергался диализу против дистиллированной воды и лиофилизации с получением фракции Naematoloma sublateritium, полученной фракционированием 80% сульфатом аммония.
[0172]
(Гидрофобная хроматография)
Naematoloma sublateritium, полученный фракционированием 80% сульфатом аммония, заливали в Butyl-TOYOPEARL 650M (TOSOH CORPORATION), уравновешенный с помощью 2M сульфата аммония-PBS, для очистки путем гидрофобной хроматографии. Этот вид хроматографии предусматривает сбор, ультрафильтрацию и лиофилизацию фракций, элюированных дистиллированной водой, с получением грубой фракции лектина из Naematoloma sublateritium (показана ←→ на рис.30).
[0173]
(Обращенно-фазовая хроматография)
Грубую фракцию лектина из Naematoloma sublateritium загружали в колонку с C8 (Wako Pure Chemical Industries, Ltd.), равновесие в которой достигалось с помощью 0,05% трифторуксусной кислоты (TFA)/ацетонитрила (100/0). При использовании этой разновидности хроматографии собирали фракцию, элюированную 0,05% TFA/ацетонитрилом (70/30) (показана ←→ на рис.31). Далее удаляли растворитель путем испарения при комнатной температуре, а образовавшиеся сухие порошки собирали с получением NSL.
[0174]
SDS-PAGE (PhastGel, Gradient 10-15) проводили с помощью системы Phastsystem (GE Healthcare Bio-Sciences). Исследуемый раствор и маркер молекулярного веса брали в количестве 1 мкл. Электрофорез проводили на основе протокола продукта с помощью традиционного метода. На рис.32 показаны результаты SDS-PAGE для NSL, где полосы 1 и 2 означают следующее. Полоса M: маркер молекулярного веса, полоса 1: NSL, 2-меркаптоэтанол (+), полоса 2: NSL, 2-меркаптоэтанол (-), гель: градиент 10-15 (GE Healthcare Bioscience), Образец: 1 мкл/полоса, краситель: серебряный.
[0175]
При проведении SDS-PAGE с 10-15% гелем подтверждено, что основным компонентом является NSL.
[0176]
(2) Получение LSL (пример 4)
На основе процесса очистки, показанного на рис.33, лектин из Lepista sordida (LSL) был очищен из гриба Lepista sordid.
[0177]
(Экстракция)
Высушенные сублимацией порошки Lepista sordida (40 г), полученные сублимационной сушкой примерно 400 г Lepista sordida, подвергали экстракции с помощью 0,8 л PBS при 4 градусах C в течение 2 часов. Полученную жидкость центрифугировали (10000 об./мин, 20 мин, 4 градуса C). Далее надосадочную жидкость фильтровали через марлю с получением первого экстракта. Остаток подвергался повторной экстракции 0,4 л PBS при 4 градусах C в течение ночи. Затем жидкость центрифугировали (10000 об./мин, 20 мин, 4 градуса C) и надосадочную жидкость фильтровали через марлю с получением второго экстракта. Затем эти экстракты объединяли с получением жидкого экстракта Lepista sordida.
[0178]
(Осаждение сульфатом аммония)
Твердый (NH4)2SO4 (0,8 кг) добавляли к полученной надосадочной жидкости (0,5 л) до 80% насыщения. После оставления на ночь при 4 градусах C осадок собирали центрифугированием (10000 об./мин, 20 мин, 4 градуса C) и подвергали диализу против дистиллированной воды и лиофилизации с получением фракции Lepista sordid, полученной фракционированием 80% сульфатом аммония.
К 0,5 л жидкости добавляли примерно 0,8 кг сульфата аммония для достижения концентрации 80% насыщения сульфата аммония и полученную жидкость перемешивали. Затем было подтверждено полное растворение. После этого жидкость оставили на ночь при 7 градусах C. Этот раствор центрифугировали (10000 об./мин, 20 мин, 4 градуса C) и к осадку добавляли небольшое количество дистиллированной воды. Далее полученный раствор суспендировали с целью сбора фракции Lepista sordid, полученной фракционированием 80% сульфатом аммония. Собранная фракция Lepista sordid, полученная фракционированием 80% сульфатом аммония, подвергалась диализу против чистой воды с использованием диализной пленки (фракция 6000-8000).
[0179]
(Гидрофобная хроматография)
Фракцию Lepista sordid, полученную осаждением 80% сульфатом аммония, загружали в Butyl-TOYOPEARL 650M (TOSOH CORPORATION), уравновешенный 2M сульфатом аммония-PBS для очистки путем гидрофобной хроматографии. Этот вид хроматографии предусматривает сбор, ультрафильтрацию и лиофилизацию фракций, элюированных дистиллированной водой, с получением грубой фракции лектина из Lepista sordida (показана ←→ на рис.34).
[0180]
(Обращенно-фазовая хроматография)
Грубую фракцию лектина из Lepista sordida загружали в колонку с C8 (Wako Pure Chemical Industries, Ltd.), равновесие в которой достигалось с помощью 0,05% трифторуксусной кислоты (TFA)/ацетонитрила (100/0). При использовании этой разновидности хроматографии собирали фракцию, элюированную 0,05% TFA/ацетонитрилом (70/30) (показана ←→ на рис.35). Далее удаляли растворитель путем испарения при комнатной температуре, а образовавшиеся сухие порошки собирали с получением LSL.
[0181]
SDS-PAGE (PhastGel, Градиент 10-15) проводили с помощью системы Phastsystem (GE Healthcare Bio-Sciences). Исследуемый раствор и маркер молекулярного веса брали в количестве 1 мкл. Электрофорез проводили на основе протокола продукта с помощью традиционного метода. На рис.36 показаны результаты SDS-PAGE для LSL. На рис.36 полоса M и полоса 1 означают следующее. Полоса M: маркер молекулярного веса, полоса 1: LSL (гидрофобная хроматография, обращенно-фазовая): 2-меркаптоэтанол (+), гель: Градиент 10-15 (GE Healthcare Bioscience), образец: 1 мкл/полоса, краситель: серебряный.
[0182]
При проведении SDS-PAGE с 10-15% гелем подтверждено, что основным компонентом является LSL.
[0183]
(3) Свойства NSL и LSL
(Масс-спектрометрический анализ MALDI-TOF)
NSL из примера 3 и LSL из примера 4 в количестве 10 мкг соответственно отдельно растворяли в TA (смесь с объемным соотношением 0,1% TFA и ацетонитрила 2:1) с помощью процедуры, аналогичной процедуре из примера 1. Далее смесь, полученную путем растворения насыщенного образца, в TA- и лектиновом TA-растворе при объемном соотношении 4:1, добавляли по каплям в количестве 1,0 мкл на подложку с получением образца. Для измерения молекулярного веса NSL и LSL в режиме LP использовали прибор Autoflex (Bruker Daltonics K.K.).
Как показывает результат, молекулярный вес составляет около 4500 m/z (рис.37 и рис.38).
[0184]
(Анализ аминокислотной последовательности)
Для NSL из примера 3 анализировали аминокислотную последовательность с помощью прибора Protein Peptide Sequencer PPSQ-21 (SHIMADZU CORPORATION). NSL из примера 3 представлял собой смесь двух белков, которые имеют аминокислотную последовательность SEQ ID NO: 5-6. Эти последовательности оказались новыми.
[0185]
Аналогично для LSL из примера 4 анализировали аминокислотную последовательность с помощью прибора Protein Peptide Sequencer PPSQ-21 (SHIMADZU CORPORATION). Результат представлял собой аминокислотную последовательность SEQ ID NO: 4. Эта последовательность также оказалась новой.
[0186]
(4) Анализ сахарсвязывающей специфичности NSL и LSL
Для NSL из примера 3 и LSL из примера 4 оценивалась сахарсвязывающая специфичность по отношению к L-фукоза α1→6 сахарной цепи тем же методом, что и в примере 1. В частности, рассчитывались константы ассоциации (Ka) для NSL и LSL. Результаты представлены в таблицах 11-13.
[0187]
NT: Испытания не проводились
[0188]
[0190]
В случае NSL из примера 3 и LSL из примера 4 обнаруживали только L-фукоза α1→6 сахарную цепь, а не содержащая L-фукозы α1→6 сахарная цепь и сахарная цепь, не содержащая L-фукозы, не обнаруживались. Кроме того, NSL из примера 3 и LSL из примера 4 также прочно связывались с три- или тетраантенной L-фукоза α1→6 сахарной цепи. Более того, даже при добавлении сиаловой кислоты константа ассоциации с L-фукоза α1→6 сахарной цепью не уменьшалась.
[0191]
И наконец, в таблице 14 показаны результаты расчета сходства между белками или пептидами для номеров последовательностей 2-6. Как видно из результатов, L-фукоза α1→6 специфичный лектин может быть получен при достижении как минимум 37% сходства с аминокислотной последовательностью, выбранной из SEQ ID NO: 2-6.
[0192]
Реферат
Группа изобретений относится к области биохимии. Предложен L-фукоза α1→6-специфичный лектин, экстрагируемый из базидиального гриба или сумчатого гриба, характеризующийся пиковым значением молекулярной массы около 4500 m/z, определяемым при масс-спектрометрическом анализе MALDI-TOF. Новый L-фукоза α1→6-специфичный лектин обладает высоким сродством к L-фукоза α1→6 сахарной цепи, определяемым константой ассоциации 1,0×10Мили более (при 25ºС), и имеет константу ассоциации 1,0×10Мили менее (при 25ºС) с высокоманнозными сахарными цепями и/или гликолипидами, не содержащими L-фукоза α1→6 сахарную цепь. В одном варианте предложенный L-фукоза α1→6 специфичный лектин представляет собой белок или пептид, состоящий из аминокислотной последовательности, выбранной из SEQ ID NO:2-6. L-фукоза α1→6-специфичный лектин используют для специфического обнаружения L-фукоза α1→6 сахарной цепи и эффективной очистки L-фукоза α1→6 сахарной цепи или сахарной цепи, не содержащей L-фукозу α1→6. 6 н. и 10 з.п. ф-лы, 38 ил., 8 табл., 4 пр.
Комментарии