Мутантная молочная бактерия вида streptococcus thermophilus, содержащая нефосфорилируемую лактозопермеазу - RU2422527C2
Код документа: RU2422527C2
Чертежи





Описание
Настоящее изобретение относится к области молочной ферментации. Более конкретно это изобретение относится к новым мутантам Streptococcus thermophilus, экспрессирующим мутантную лактозопермеазу, транспортная активность которой модифицирована. Эти штаммы и ферменты, их содержащие, можно использовать для получения ферментированных молочных продуктов, обладающих улучшенными свойствами хранения.
Йогурты обычно получают путем ферментации молока при помощи ассоциации различных молочных бактерий, выбираемых из штаммов Streptococcus thermophilus и Lactobacillus bulgaricus. В процессе ферментации, которую проводят при температуре примерно 40-45°С, эти бактерии используют лактозу главным образом в качестве энергетического субстрата и продуцируют молочную кислоту, вызывающую коагуляцию молока; когда рН достигает значения примерно 4,8-4,5, эту стадию ферментации (называемую также «окислением») останавливают путем охлаждения продукта. Последний затем выдерживают на холоде во время дальнейшего процесса производства и хранения до его употребления.
Однако охлаждение не полностью останавливает молочную ферментацию; даже если температура продукта поддерживается 4°С, со временем наблюдается постепенное повышение его кислотности.
Это явление, известное под названием постокисления, вызывает ухудшение органолептических показателей продукта при его хранении.
Постокисление является по существу результатом присутствия через бактерии остаточной лактозы в продукте по окончании стадии контролируемого окисления. Для того чтобы его избежать, предлагалось использовать штаммы молочных бактерий, не ферментирующие или ферментирующие очень незначительно лактозу.
Ферменты, являющиеся главными для ферментации лактозы в Streptococcus thermophilus и Lactobacillus bulgaricus, кодируются опероном лактозы, который содержит ген lacS, кодирующий лактозопермеазу, и ген lacZ, кодирующий β-галактозидазу. Эти белки соответственно отвечают за транспорт и гидролиз лактозы. Было таким образом предложено для получения непостокисляющих штаммов молочных бактерий получать искусственные варианты или селекционировать натуральные мутанты, в которых активность по меньшей мере одного из этих ферментов подавлена.
Патент ЕР 1078074 (Compagnie Gervais Danone) относится к мутантам L.Bulgaricus с подавленной активностью β-галактозидазы, содержащим антисмысловую мутацию по меньшей мере в одном из генов оперона лактозы. В этом патенте более конкретно описан мутант, анализ последовательности которого показывает две точечные мутации: одна вводит терминирующий кодон в ген β-галактозидазы, который индуцирует неспособность этого мутанта использовать лактозу; другая мутация индуцирует изменение аминокислоты на уровне гена пермеазы (Lys->Ans в положении 122); в ЕР 1078074 не указано влияние этой мутации на фенотип мутанта.
В WO 0188150 описаны мутанты штамма Lactobacillus. Эти мутанты неспособны использовать лактозу, но сохраняют способность экспрессировать β-галактозидазу. В WO 0188150 не уточняется природа или положение мутации, о которой идет речь, а указано только, что она может находиться в одном из структурных генов оперона лактозы, например, в пермеазе, или на участке регуляции операна лактозы или в гене, участвующем в контроле экспрессии оперона лактозы.
Общим для мутантов, описанных в указанных выше документах, является полная неспособность использовать лактозу. Они могут расти на молоке, только если в него добавлен другой сахар, не являющийся лактозой, обычно глюкоза. Свойства окисления и постокисления этих мутантов контролируются посредством количества добавленной глюкозы.
Для обнаружения альтернативы этим мутантам заявители рассмотрели возможность получения штаммов молочных бактерий, обладающих, с одной стороны, способностью использовать лактозу в процессе их роста, сопоставимой со способностью диких штаммов, а с другой стороны, ограниченной способностью использовать лактозу в стационарной фазе для уменьшения или устранения явления постокисления. С этой целью они заинтересовались возможностью воздействовать на регуляцию транспорта лактозы в клетки молочных бактерий и, в частности, в клетки S.Thermophilus.
Транспорт внеклеточной лактозы в клетки S.Thermophilus осуществляется посредством лактозопермеазы LacS. Этот транспорт лактозы является протонным с импортом или антипортом с помощью внеклеточной галактозы в результате разложения лактозы.
Транспорт лактозы зависит от двух явлений: с одной стороны, состояния фосфорилирования лактозопермеазы и, с другой стороны, экспрессии гена lacS, кодирующего эту лактозопермеазу. Оба аспекта подробно описаны ниже.
Фосфорилирование лактозопермеазы
Белок LacS состоит из домена транслокации и домена регуляции (IIA). Эти домены содержат различные гистидиновые остатки, фосфорилирование которых участвует в регуляции транспорта лактозы. В частности, домен IIA фосфорилируется по гистидину 552. Это фосфорилирование осуществляется белком HPr (histidine-containing phosphocarrier protein), который предварительно фосфорилирован.
HPr может фосфорилироваться:
по серину АТФ-зависимой протеинкиназой; реакция обратного гидролиза HPr(Ser-P) катализуется фосфатазной активностью (HPr(Ser-P) фосфатазой);
по гистидину HPr(His-P) фосфорильной группой, происходящей из фосфоенолпирувата и через посредство фермента I(EI).
Только форма HPr, фосфорилированная по гистидину, позволяет фосфорилировать лактозопермеазу по гистидину 552.
На модели in vitro протеолипосом, реконструирующих мембранное окружение белка LacS и его фосфорилирование посредством HPr(His-P), было обнаружено, что это фосфорилирование не воздействует на симпорт лактозы протоном (Gunnewijk and Poolman, 2000a), а увеличивает примерно на фактор 2 поток обмена лактоза/галактоза.
Транскрипция гена lacS
Транскрипция оперона лактозы индуцируется ростом в лактозной среде. Промотер генов lacS и Z содержит сайт cre (catabolite responsive element), который обеспечивает регуляцию при помощи СсрА: СсрА подавляет экспрессию lacS и lacZ. СсрА, напротив, активирует транскрипцию гена, кодирующего лактат дегидрогеназу (van den Bogaard et al., 2000).
Форма HPr(Ser-P) способна взаимодействовать с СсрА. Вместе эти белки обеспечивают образование комплекса с сайтом cre, вследствие чего происходит репрессия транскрипции гена lacS (Jones et al., 1997).
Было отмечено (Gunnewijk and Poolman, 2000b), что форма HPr(Ser-P) является доминирующей в начале экспотенциальной фазы роста культур S.thеrmophilus и в процессе последней уменьшается, тогда как форма HPr(His-P) появляется во время экспотенциальной фазы и достигает максимума при входе в стационарную фазу. Переход от HPr(Ser-P) к HPr(His-P) происходит параллельно с уменьшением содержания лактозы и повышением содержания галактозы в культурной среде и очень существенным увеличением экспрессии лактозопермеазы.
Таким образом состояние фосфорилирования протеина HPr очевидно играет роль в регулировании транспорта лактозы, компенсируя уменьшение содержания лактозы в среде посредством, с одной стороны, уровня экспрессии белка lacS и, с другой стороны, регуляции его активности.
На основании наблюдений, указанных выше, Gunnewijk и Poolman предложили следующую модель: если лактоза содержится в среде в избыточном количестве, экспрессия гена lacS подавляется комплексом HPr(Ser-P)/CcpA. Во время ферментации аккумуляция галактозы в среде и уменьшение свободной лактозы приводит к снижению способности бактерии вызывать проникновение лактозы (и, следовательно, понижение окисления среды). Это понижение вызывает снижение гликолитической активности и падение концентрации АТФ, а также повышение концентрации неорганического фосфата, вызывая снижение активности HPr(Ser-P)киназы в пользу активности HPr(Ser-P)фосфатазы, в результате чего понижается концентрация HPr(Ser-P). Это понижение концентрации HPr(Ser-P) позволяет повысить катаболическую репрессию гена lacS и, следовательно, повысить продуцирование лактозопермеазы. Параллельно увеличение HPr(His-P) позволит усилить фосфорилирование лактозопермеазы по гистидину 552 и, следовательно, способность к антипорту лактозы галактозой.
Эта модель, показывающая, что фосфорилирование лактозопермеазы по гистидину 552 при помощи HPr(His-P) увеличивает приток лактозы в клетки, когда количество субстрата в среде уменьшается, позволяет предполагать, что окисление в конце экспотенциальной фазы могло бы замедлиться, если этому фосфорилированию препятствовать. Однако она частично основана на экспериментах in vitro и не позволяет судить о реальном вкладе in vivo усиления фосфорилирования лактозопермеазы по сравнению с усилением ее экспрессии в притоке извне лактозы in vivo. Кроме того наблюдения, касающиеся влияния концентрации HPr(Ser-P) и HPr(His-P) на усиление экспрессии и фосфорилирования LacS, проводились на бактериях в экспотенциальной фазе или в начале стационарной фазы; не представлено никакой информации относительно концентраций этих двух форм HPr на более поздних стадиях стационарной фазы.
Единственная информация о влиянии отсутствия фосфорилирования LacS по гистидину 552 представлена только в публикации Poolman et al. (Poolman et al., 1992), в которой описаны различные плазмиды, содержащие последовательность, кодирующую фермент LacS Streptococcus thermophilus, мутированную на различных остатках гистидина. Эти плазмиды использовали для трансформирования штамма E.coli, в котором эндогенный ген lacS был предварительно делетирован. Транспорт лактозы в штаммах, трансформированных различными мутантами, оценивали по сравнению с доставкой, наблюдаемой в том же штамме E.coli, содержащем плазмиду, кодирующую фермент дикого типа LacS Streptococcus thermophilus. Никакой существенной разницы не наблюдалось в том, что касается мутанта H552R, в котором нативный гистидин заменен аргинином. Poolman et al. связывают этот результат или с неэффективностью фосфорилирования дикого фермента LacS Streptococcus thermophilus посредством HPr(His-P) E.coli или с тем фактом, что это фосфорилирование не выполняет функции транспорта лактозы.
Авторы тем не менее искали ответ на вопрос, может ли мутация, предшествующая фосфорилированию LacS по остатку гистидина, влиять на свойства окисления и постокисления мутантной бактерии.
Они выбрали для этого исследования промышленный штамм Streptococcus thermophilus. Это штамм, депонированный в CNCM 12.12.2002 под номером I-2967, позволяет получать молочные продукты с интересной текстурой; тем не менее этот штамм приводит к значительному постокислению.
Авторы получили и охарактеризовали мутант этого штамма, экспрессирующий вместо дикой лактозопермеазы мутантную лактозу пермеазу, нефосфорилируемую по гистидину 552.
Они констатировали, что этот мутантный штамм имеет кривую окисления, отличающуюся от кривой материнского штамма. Окисление начинается медленнее в случае мутанта, чем в случае материнского штамма, и максимальная скорость окисления мутанта менее высокая. Тем не менее эквивалентная величина рН достигается через 6 часов с начала ферментации для обоих штаммов. Наиболее четкое различие между этими двумя штаммами прослеживается на уровне постокисления. В одинаковых условиях хранения (28 дней хранения при 10°С), ΔрН (разница между рН в нулевой день и рН в 28-й день) составляет порядка 0,6 в случае материнского штамма и порядка 0,4 в случае мутантного штамма. Это различие в постокислении происходит не вследствие разной выживаемости бактерий. Последняя является действительно одинаковой для обоих штаммов. Кроме того, ферментированные продукты, полученные с использованием мутанта, имеют те же качества текстуры, что и полученные с использованием материнского штамма.
Таким образом, прежде всего настоящее изобретение относится к способу получения мутантного штамма молочной бактерии, обладающего более слабым постокислением, чем материнский штамм, от которого он произошел, отличающемуся тем, что в ДНК генома, в частности в хромосомный ДНК указанного материнского штамма, вводят мутацию кодона, кодирующего гистидин, фосфорилируемый посредством HPr(His-P) домена IIA лактозопермеазы, причем указанная мутация индуцирует замену указанного гистидина нефосфорилируемой аминокислотой.
В соответствии с предпочтительным вариантом осуществления настоящего изобретения указанный штамм является штаммом Streptococcus thermophilus, а указанная мутация индуцирует замену гистидина в положении 552 лактозопермеазы нефосфорилируемой аминокислотой.
Указанная нефосфорилируемая аминокислота может быть любой аминокислотой за исключением серина, тирозина, гистидина и треонина. Предпочтительно выбирают аланин.
Преимущественно кодон, кодирующий гистидин в положении 552 лактозы пермеазы, заменяют кодоном, кодирующим аланин. Эта мутация генерирует сайт рестрикции BstUI, который облегчает сортировку полученных мутантов.
Способ по изобретению можно осуществлять путем использования традиционных технологий управляемого мутагенеза, хорошо известных специалисту, в частности, мутагенеза посредством PCR.
Мутантную ДНК, полученную таким способом, затем вводят в вектор, позволяющий интегрировать ген в хромосому бактерии. Предпочтительно это интегрирование осуществляют путем рекомбинации вставки, которую несет вектор с гомологичной областью бактериальной хромосомы.
Обычно мутантную ДНК вводят в интегративный вектор, несущий маркер селекции (например, ген, устойчивый к антибиотику) и вводят этот вектор в бактерии, в которых хотят провести мутацию. Последний затем культивируют на селективной среде (например, если маркер селекции является геном, устойчивым к антибиотику, в присутствии соответствующего антибиотика) и рекуперируют бактерии, способные расти в этих условиях, являющиеся бактериями, в которые интегрировали вектор путем гомологичной рекомбинации между вставкой и гомологичной областью хромосомы бактерии. Структура, интегрированная в хромосому, состоит из последовательностей вектора, фланкированного с одной стороны мутантной последовательностью, происходящей из вставки, а с другой стороны гомологичной последовательностью бактерии хозяина.
Отобранные таким образом бактерии культивируют на неселективной среде с тем, чтобы позволить удаление последовательностей, происходящих из вектора, которое осуществляется путем гомологичной рекомбинации между областями, фланкирующими эти последовательности. Половина бактерий, в которых произошла такая рекомбинация, содержит «дикую» последовательность, происходящую из бактерии хозяина, а другая половина содержит мутантную последовательность, происходящую из вставки. Бактерии, содержащие мутацию, затем отбирают любым пригодным способом. Например, если мутация создает сайт рестрикции, селекцию можно осуществлять на основании присутствия этого сайта рестрикции в продукте амплификации посредством PCR мутантной области.
Интегративные векторы имеются во многих молочных бактериях. Обычно для данного вида бактерий интегративный вектор является вектором, который можно вводить в бактерии этого вида, но который неспособен к репликации в них.
В качестве примеров векторов, пригодных в качестве интегративных векторов в Streptococcus thermophilus, можно назвать Pgem5, Puc19 (Mollet et al., 1993), Pnd324 (Duan et al., 1999).
Преимущественно для повышения эффективности трансформации можно использовать в качестве интегративного вектора вектор с обусловленной репликацией в выбранной бактерии. В этом случае бактерии, в которые был введен вектор, на первой стадии культивируют в условиях, благоприятных для репликации, что позволяет ему обосноваться в этих трансформированных бактериях; на второй стадии бактерии культивируют в условиях, неблагоприятных для репликации вектора, и, как в случае традиционного интегративного вектора, можно проводить селекцию бактерий, в хромосому которых вектор был интегрирован.
В качестве примеров векторов с обусловленной репликацией, пригодных в качестве интегративных векторов в большей части молочных бактерий, можно назвать термочувствительные векторы, описанные BISWAS et al., и MAGUIN et al. (Biswas et al., 1993, Maguin et al., 1996), а также в заявке PCT WO 93/18164, или векторы pwv01 (Law et al., 1995) и Puc122 (Frere et al., 1998).
Объектом изобретения является также штамм молочных бактерий, который можно получить способом по изобретению.
Этот штамм отличается тем, что в его хромосомной ДНК содержится мутация кодона, кодирующего фосфорилируемый гистидин посредством HPr(His-P) домена IIA лактозопермеазы, причем указанная мутация индуцирует замену указанного гистидина нефосфорилируемой аминокислотой.
В соответствии с предпочтительным вариантом настоящего изобретения указанный штамм является штаммом Streptococcus thermophilus, в котором ген лактозопермеазы содержит мутацию, вызывающую замену гистидина в положении 552 белка нефосфорилируемой аминокислотой.
Штамм молочных бактерий по изобретению был депонирован в соответствии с Будапештским договором 10 мая 2004 г. в CNCM (Национальная коллекция культур микроорганизмов), Париж, ул. Доктора Ру, д.25, под номером I-3213. Речь идет о мутантном штамме S.thermophilus, который является производным штамма CNCM I-2967 (депонирован в CNCM 12 декабря 2002 г.) путем ввода посредством управляемого мутагенеза мутации, заменяющей кодон гистидина 552 аланиновым кодоном.
Штаммы молочных бактерий по изобретению во время фазы своего роста имеют активность лактозопермеазы, подобную активности материнского штамма, от которого они произошли. Таким образом, они обладают свойствами ассимиляции лактозы и окисления, сравнимыми со свойствами материнского штамма, от которого они произошли. Но их активность лактозопермеазы во время стационарной фазы ниже, чем активность материнского штамма, за счет чего постокисление уменьшается.
Преимущественно штаммы молочных бактерий по изобретению происходят от молочных бактерий, обладающих активностью β-галактозидазы, и сохраняют эту активность. Они могут нормально расти на молоке без добавления другого сахара кроме лактозы.
Предпочтительно штаммы молочных бактерий по изобретению являются мутантами, пригодными для применения в пищевой промышленности (или мутантами “food-grade”). Предпочтительно они являются производными охарактеризованных штаммов бактерий, обладающих преимущественными свойствами ферментации молочных продуктов.
Настоящее изобретение относится также к молочной закваске, содержащей по меньшей мере один штамм бактерий, такой как описан выше. В соответствии с частным способом осуществления, молочная закваска по изобретению содержит по меньшей мере один мутантный штамм S.Thermophilus, экспрессирующий лактозопермеазу, в которой гистидин 552 был заменен нефосфорилируемым остатком, в комбинации по меньшей мере с другим штаммом молочной бактерии, например, штаммом l.bulgaricus, который также может обладать пониженным постокислением (например, в результате мутации согласно изобретению лактозопермеазы или в результате мутации, инактивирующей β-галактозидазу).
Способ получения ферментированного молочного продукта, содержащий стадию, во время которой ферментируют молоко при помощи молочной закваски, такой как описана выше, также является неотъемлемой частью настоящего изобретения, также как любой ферментированный молочный продукт, который можно получить таким способом, такой как йогурт, ферментированное молоко, ферментированный напиток, кефир, сыр или ферментированное молоко для детского питания.
Экспериментальные примеры, приведенные ниже, иллюстрируемые фигурами, более подробно описывают некоторые аспекты настоящего изобретения, не ограничивая его.
Описание фигур
Фигура 1: сравнение последовательности гена lacS варианта I-3213 с последовательностью гена lacS материнского штамма I-2967.
Фигура 2: сравнение кривых окисления, полученных с мутантом I-3213 и с материнским штаммом I-2967.
Фигура 3: сравнение скорости окисления мутанта I-3213 и материнского штамма I-2967.
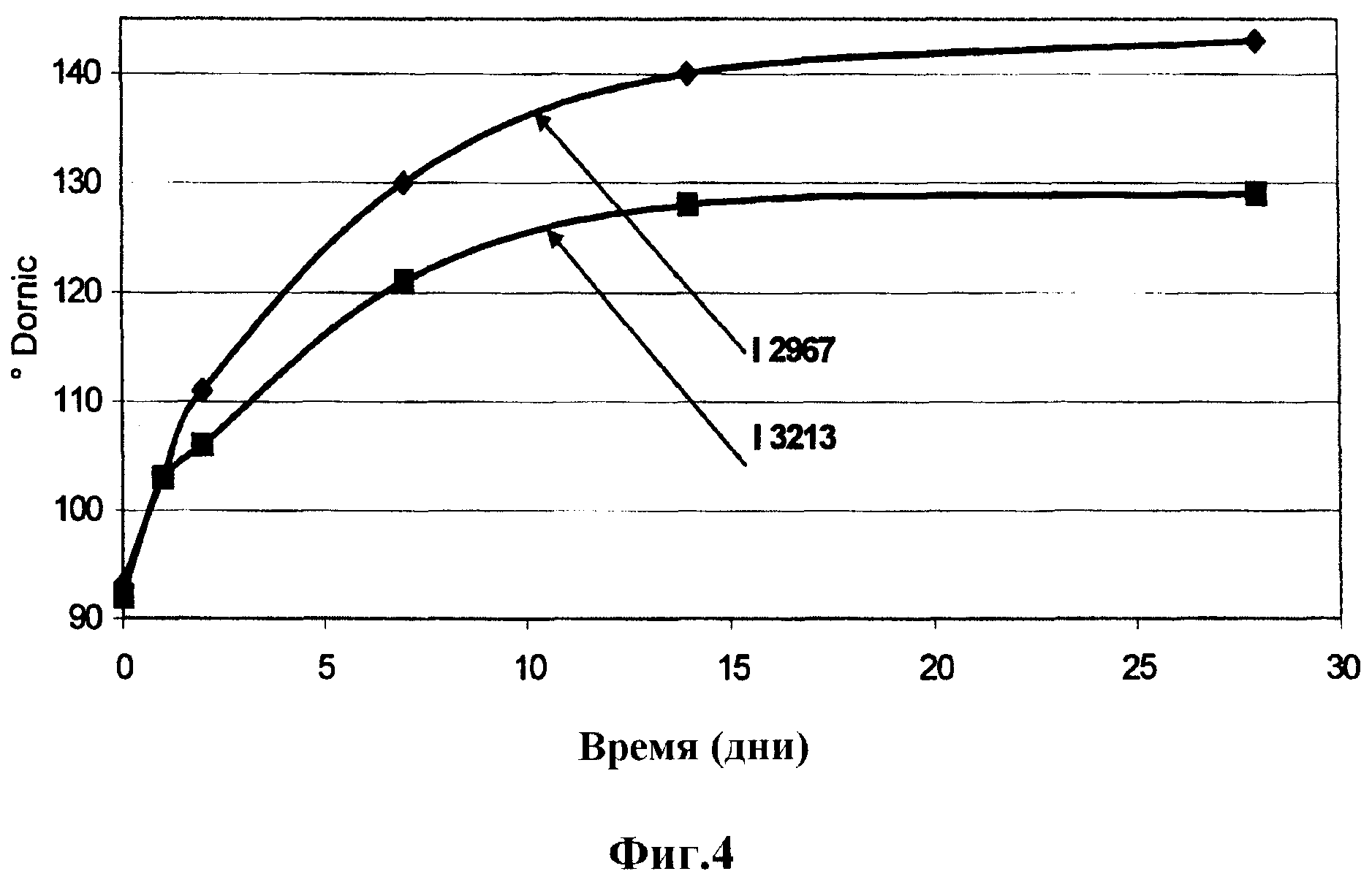
Фигура 4: Наблюдение за кислотностью Dornic во время хранения продуктов. Сравнение мутанта I-3213 и материнского штамма I-2967.
Фигура 5: Принцип измерения вязкости η продукта.
Примеры
Пример 1: Получение мутанта
Исходный штамм является штаммом S.Thermophilus I-2967, депонированным в CNCM 12 декабря 2002 г.
В последовательности гена lacS кодон гистидина 552 (фосфорилируемый гистидин) заменили кодоном аланина путем двойной рекомбинации. Кроме того, проведенная мутация (замена второго нуклеотида кодона 552 цитозином вместо аденина) создала в гене сайт рестрикции BstU1. Это позволило селекционировать клоны, интегрировавшие целевую мутацию в гене lacS.
Стабильность мутации проверяли по одному из полученных клонов (депонирован в CNCM 10 мая 2004 под номером I-3213) путем 6 последовательных пассажей с последующим секвенированием гена LacS. На фиг.1 показано сравнение последовательности гена LacS варианта I-3213 (SEQ ID NO:2) с последовательностью гена LacS материнского штамма I-2967 (SEQ ID NO:10. Наличие мутации очевидно: теперь кодон гистидина 552 кодирует аланин. Других нежелательных мутаций в гене LacS нет.
Мутант I-3213 пригоден для применения в агропищевой промышленности, т.к. не содержит никакой остаточной последовательности плазмида, используемого для интегрирования мутации в ген LacS.
Пример 2: Физиологические тесты, выполненные с мутантом I-3213
С тем чтобы установить, что мутант I-3213 обладает меньшим постокислением, чем материнский штамм I-2967, получали продукты с чистым штаммом и наблюдения проводили до J+28: окисление, постокисление, выживаемость, текстура.
2.А Протокол
Оживление штаммов путем 2 пассажей
Получение заквасок на стерильном молоке с добавлением экстрактов дрожжей и инкубацией при 44°С до достижения кислотности 85°D (соответствует 108, 109UFC/мл).
Посев смеси 120 г порошка обезжиренного молока + 930 мл воды + 1г пептона желатина N3 (Organotechnie), пастеризованной в течение 10 минут при 95°С с 1% (v/v) фермента.
Инкубация в банках в сушильном шкафу при 44°С до достижения рН 4,65.
Остановка ферментации путем помещения банок в ледяную воду на 30 минут.
Хранение продуктов в течение 28 дней в холодильной камере при 10°С.
2.В Наблюдение-Сравнение окисления при помощи мутантов и штаммов I-2967.
1.В.1 Кривые окисления
При помощи системы CINAC (Кинетика окисления, Alliance Instruments) рН измеряют непрерывно в течение всего времени. Таким образом можно получить:
кривую окисления по каждому штамму,
первичную производную относительно времени, которая ускоряет окисление.
2.В.2 Измерение рН
Наблюдение за эволюцией рН во время хранения продуктов осуществляют при помощи рН-метра МР 220 фирмы Mettler Toledo.
1.В.3 Измерение кислотности Dornic
Измерение кислотности Dornic (D°) позволяет титровать молярную концентрацию в ионах Н3О+. Количество градусов Dornic соответствует числу в десятых миллиметра раствора гидроксида натрия при концентрации 0,1 N, необходимых для нейтрализации 10,32 г молока. Нейтральность визуализируют с использованием цветового индикатора, фенолфталеина. В зоне действия (рН 8,2) фенолфталеин из бесцветного становится розовым. Градус Dornic соответствует 100 мг молочной кислоты на литр молока.
2.В.4. Получение смеси для тестирования продуктов
Молочную среду получают из 120 г порошка обезжиренного молока (Milex 240, Arla Food Ingredients)+930 мл пермутированной воды + 1 г пептида N3 (Vital Armor 950, Armor Protéines). Смесь перемешивают до полной гомогенизации. Затем среду повторно гидрируют в течение 30 минут при комнатной температуре, затем пастеризуют при 95°С в течение 10 минут.
2.В.5 Выживаемость штаммов
Измерения выживаемости штаммов проводят в среде, содержащей агар-агар М17 и сахарозу. Изолируют на поверхности при помощи спирального устройства (ensemenceur spirale)(WASP фирмы AES). Инкубируют при 37°С в присутствии Н2СО2. Считывают через 72 часа с начала инкубирования.
Кривые окисления в чистых штаммах строят в двух экземплярах.
2.В.6 Интерпретация
Результаты, изображенные на фиг.2 и 3, показывают, что кривая окисления мутанта I-3213 более плавная, чем кривая материнского штамма, с более выраженным эффектом мочевины и что максимальная скорость окисления ниже. Тем не менее рН 4,50 обоих штаммов достигается за 6 часов.
2.С Наблюдение-Сравнение постокисления при помощи мутанта I-3213 и штамма I-2967
На фиг.4 показаны результаты наблюдения за кислотностью Dornic во время хранения продуктов путем сравнения продуктов, полученных ферментацией при помощи мутанта I-3213 и материнского штамма I-2967.
Результаты измерения рН непосредственно после остановки ферментации (J0) и через 28 дней хранения при 10°С представлены ниже в таблице 1.
Совокупность этих результатов подтверждает, что мутант обладает меньшим постокислением, чем материнский штамм.
2.D - Наблюдение-Сравнение выживания мутантов и штамма I-2967
Ниже в таблице 2 показано число колоний бактерий, присутствующих в 1 мл продукта через 28 дней с начала хранения при 10°С.
Эти результаты показывают, что через 28 дней хранения после остановки ферментации мутанты обладают такой же высокой выживаемостью, что и материнский штамм.
2.Е - Наблюдение-Сравнение текстуры продуктов, полученных при помощи мутанта, и продуктов, полученных при помощи материнского штамма I-2967
Проводят измерения текстуры продуктов одной серии производства. Все измерения проводят в трех экземплярах (3 банки для снятия одного измерения).
Для измерения текстуры продуктов в J+7 применяли три метода измерения:
Измерение пенетрометрией с помощью TAXT2 (10°С).
Измерение потока после перемешивания вручную при помощи Rhéomat 260 (4°С).
Измерение вязкости по сыворотке после центрифугирования на MCR300 (20°С).
Эти разные методы измерения текстуры подробно описаны ниже.
2.Е.1 - Измерение вязкости по сыворотке твердых продуктов
Интерес этого метода заключается в анализе сыворотки твердых продуктов в целях подтверждения реологических характеристик молока, ферментированного при помощи штаммов.
Этот метод анализа позволяет на первой стадии рекуперировать сыворотку твердых продуктов. Для этого количество продукта, примерно 50 г, центрифугируют при 631 g в течение 10 минут при комнатной температуре, что позволяет собрать сыворотку, присутствующую в желеобразных молочных продуктах, ферментированных штаммами. Затем сыворотку извлекают и подвергают такому же центрифугированию с тем, чтобы устранить большую часть остатков продукта. Остаток этих частиц осаждается и образует хрупкий осадок.
Затем вязкость сыворотки измеряют при 20°С и фиксированном градиенте сдвига 100s-1 в течение минуты. Три измерения проводят также по трем банкам с молоком, ферментированным одним и тем же штаммом в одних и тех же условиях. Устройство, применяемое для этого анализа, представляет собой реометр Anton Paar Physica® MCR 300, снабженный коаксиальной геометрией с двойными зазорами типа DG 26.7/TEZ 150 р-с, а также системой регулировки температуры с эффектом Peltier. Эта поворотная система позволяет оценивать вязкость сыворотки с постоянным градиентом сдвига на один пункт в секунду.
Обычно две первые величины этого измерения являются непоследовательными и искаженными инициализацией поворотной системы. Поэтому вязкость каждой сыворотки [Vs] определяют по средней величине, вычисляемой по величинам, измеренным устройством, за исключением двух первых.
2.Е.2 - Измерение пенетрометрией (Fgel-Dgel-F15vv)
Устройством, используемым для этого измерения, является Thermo Rhéo TAXT2 (Anton Peer Physica, Австрия).
Цилиндр диаметром примерно 1 см проникает в гель (при температуре 10°С) с постоянной скоростью на 15 мм глубины. Во время погружения подвижного элемента в продукт гель сопротивляется и деформируется прежде, чем происходит разрыв. Измеряют силу, полученную в результате.
Получают следующие параметры:
Fgel - сила геля (g), соответствующая величине силы, прилагаемой подвижным элементом в момент разрыва геля (первый пик кривой).
Dgel - расстояние, соответствующее силе геля (мм), т.е. глубина, на которую погружается подвижный элемент в момент разрыва геля.
F15 - сила на глубине 15 мм (g), соответствующая силе, измеренной при остановке движения подвижного элемента.
2.Е.3 Измерение потока-Вязкость потока
Этот метод заключается в определении вязкости твердых продуктов после перемешивания вручную и инкубации в течение 30 минут при 4°С. При 4°С проводят три измерения по трем банкам молока, ферментированного одним и тем же штаммом в одних и тех же условиях. Для этого анализа используют вязкозиметр Mettler® RM 260, охлажденный и снабженный коаксиальной системой типа DIN 145. Эта поворотная система позволяет наблюдать деструкцию продукта в зависимости от градиента линейного сдвига, т.е. напряжение при заданном градиенте.
Результаты получают в виде кривой непрерывного потока, с восходящим и нисходящим участком от 0 до 20s-1.Продукт подвергается градиенту сдвига, увеличивающемуся от 0 до 20s-1 за 1 минуту. Эта фаза соответствует восходящему участку. Затем он подвергается градиенту сдвига, уменьшающемуся от 20 до 0 s-1 за 1 минуту, что соответствует нисходящему участку.
Каждая нисходящая кривая моделируется затем по модели Кассона:

τ - напряжение (Па);
τ0 - порог потока продукта (Па) [порог 4];
η - вязкость продукта (Па·с) [V4];
D - градиент сдвига (s-1).
Это моделирование по Кассону с последующей прямой линейной регрессии на нисходящей части кривой позволяет установить важный параметр, которым является вязкость продукта η, соответствующий наклону прямой регрессии.
На фиг.5 показан способ вычисления вязкости по этой модели.
2.Е.4 Результаты
Результаты, полученные каждым методом измерения, представлены ниже на Таблице 3.
Дисперсионный анализ (Р<0,05) проводят по результатам измерений текстуры (по каждому параметру величины сравнивают тестом Student):
Параметры Cила геля, Расстояние при силе геля и Сила при 15 мм показывают, что оба штамма не имеют существенных различий.
Параметр вязкости, полученный при измерении потока, показывает, что оба штамма не имеют существенных различий.
Вязкость сыворотки, будучи воспроизводимой, показывает существенное различие между двумя штаммами, но при этом мутант обладает большей вязкостью, чем штамм I-2967, это доказывает, что изменений текстуры не произошло.
2.Е.5 - Интерпретация
Измерения текстуры ферментированных продуктов, полученных при помощи мутанта и материнского штамма, позволяют показать, что изменений текстуры, связанных с мутацией, не произошло.
2.F - Выводы
Мутант штамма I-2967 с нефосфорилируемой лактозопермеазой был получен двумя рекомбинациями.
Этот мутант, названный I-3213, имеет:
кривую окисления, отличную от кривой материнского штамма (замедленная скорость),
меньшее постокисление на 28 день,
текстуру, подобную текстуре материнского штамма, и
Хорошую выживаемость на 28 день.

Реферат
Изобретение относится к области биотехнологии, а именно к способу получения мутантной молочной бактерии вида Streptococcus thermophilus, молочной закваске, способу получения ферментированного молочного продукта и к ферментированному молочному продукту. Предложенное изобретение может быть использовано для получения ферментированных молочных продуктов, обладающих улучшенными свойствами хранения. Способ получения мутантной молочной бактерии вида Streptococcus thermophilus, обладающей более слабым постокислением, чем материнский штамм, осуществляют введением в ДНК генома указанного материнского штамма мутации кодона 552, кодирующего гистидин, домена IIА лактозопермеазы. При этом указанная мутация индуцирует замену указанного гистидина аминокислотой, отличной от серина, тирозина, гистидина и треонина. Предложенное изобретение позволяет получить мутантную молочную бактерию вида Streptococcus thermophilus, обладающую пониженным постокислением и пригодную, в частности, для получения ферментированных молочных продуктов. 5 н. и 5 з.п. ф-лы, 5 ил., 3 табл.
Комментарии