Полипептид с активностью фосфолипазы и его применение - RU2363731C2
Код документа: RU2363731C2
Чертежи
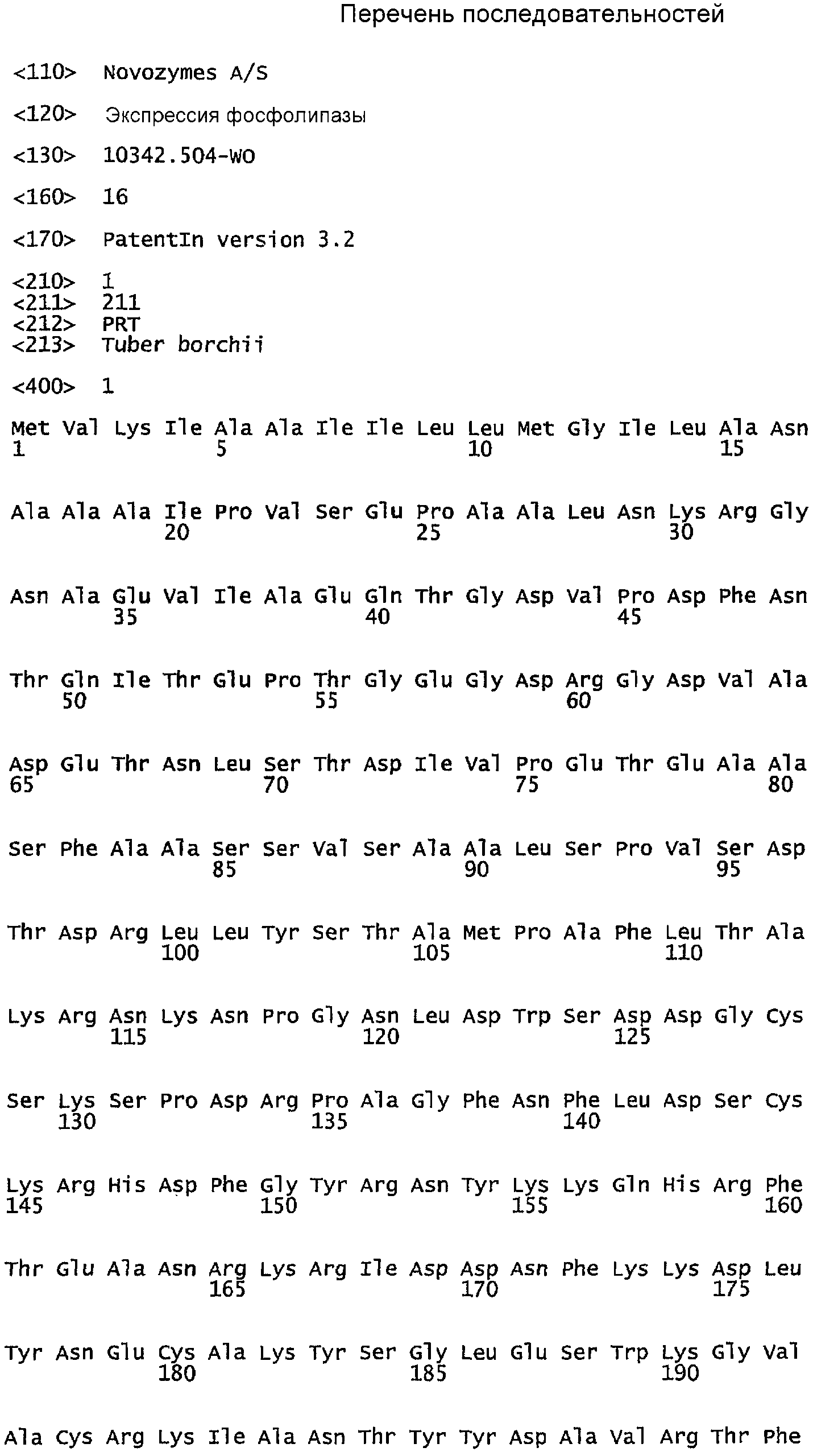
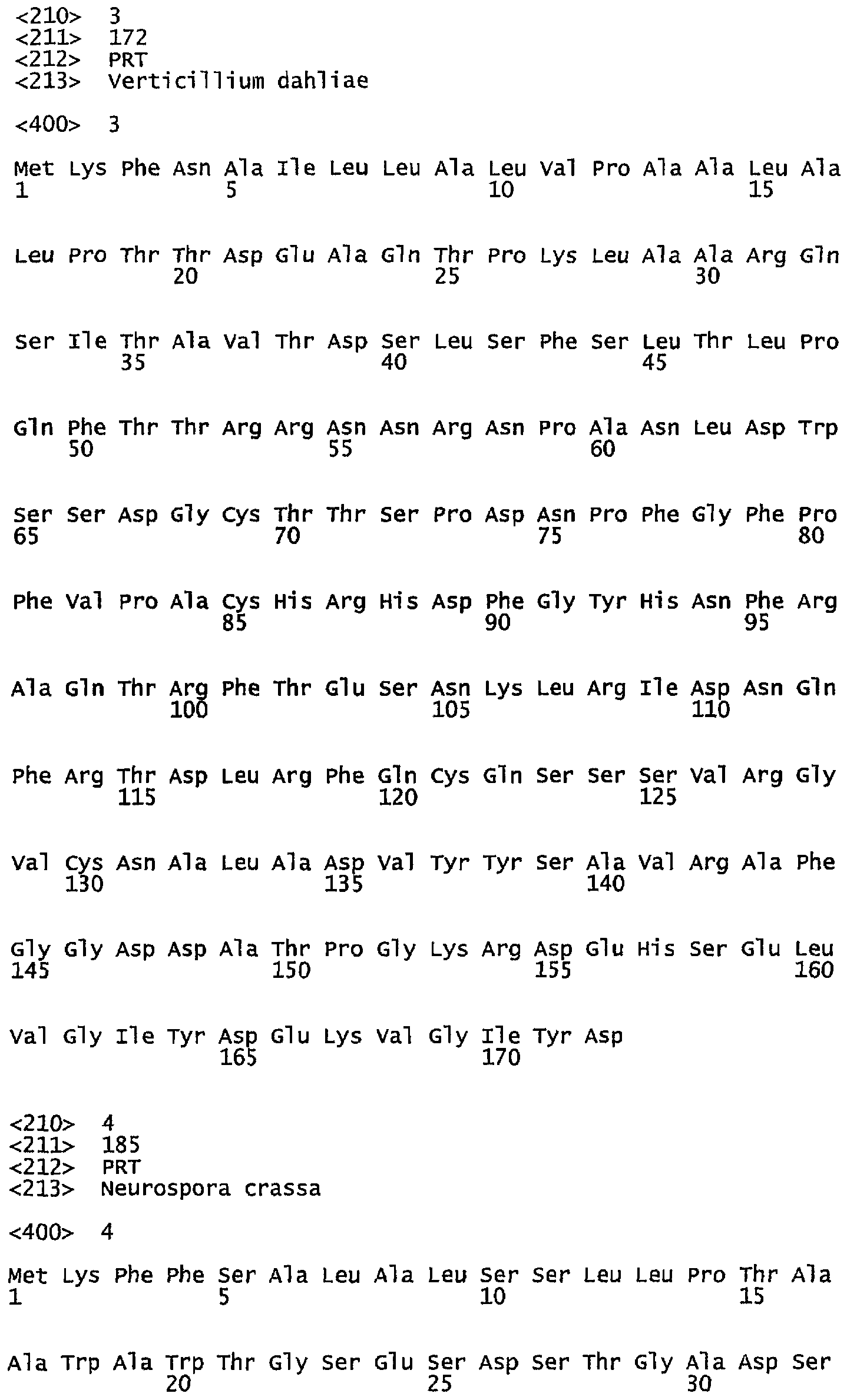
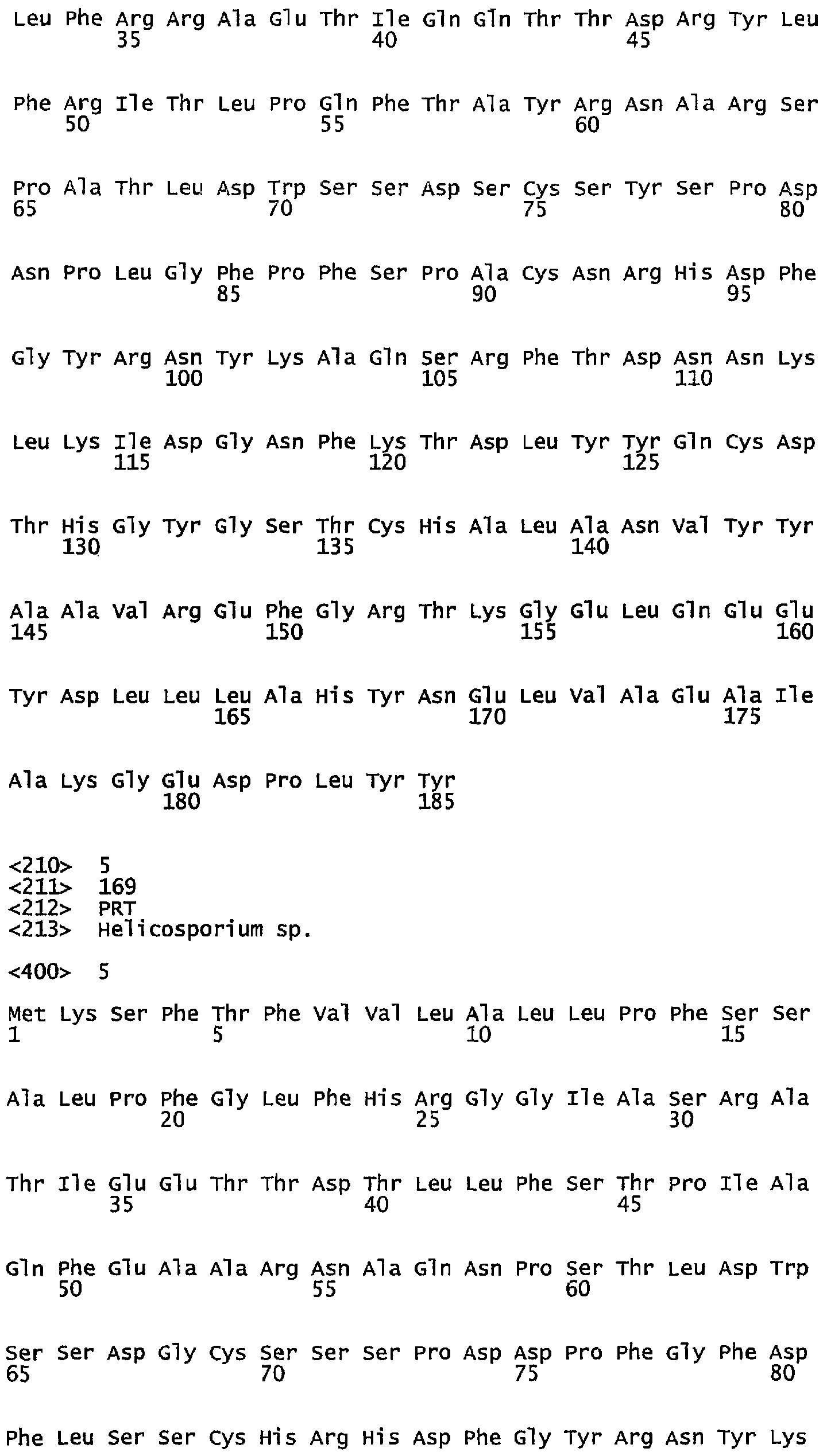









Описание
Изобретение относится к способу гидролиза фосфолипида, к способу получения фосфолипазы, к способу получения сыра и к фосфолипазе.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Сораньи с соавт. (Soragni, E., et al. (2001) EMBO J. 20:5079-5090) описывает фосфолипазу (TbSP1) из Tuber borchii и нуклеотидную последовательность к ДНК гена, кодирующего ее. Ниже приведена информация по разным источникам о пептидной последовательности, полученной из указанных ниже организмов:
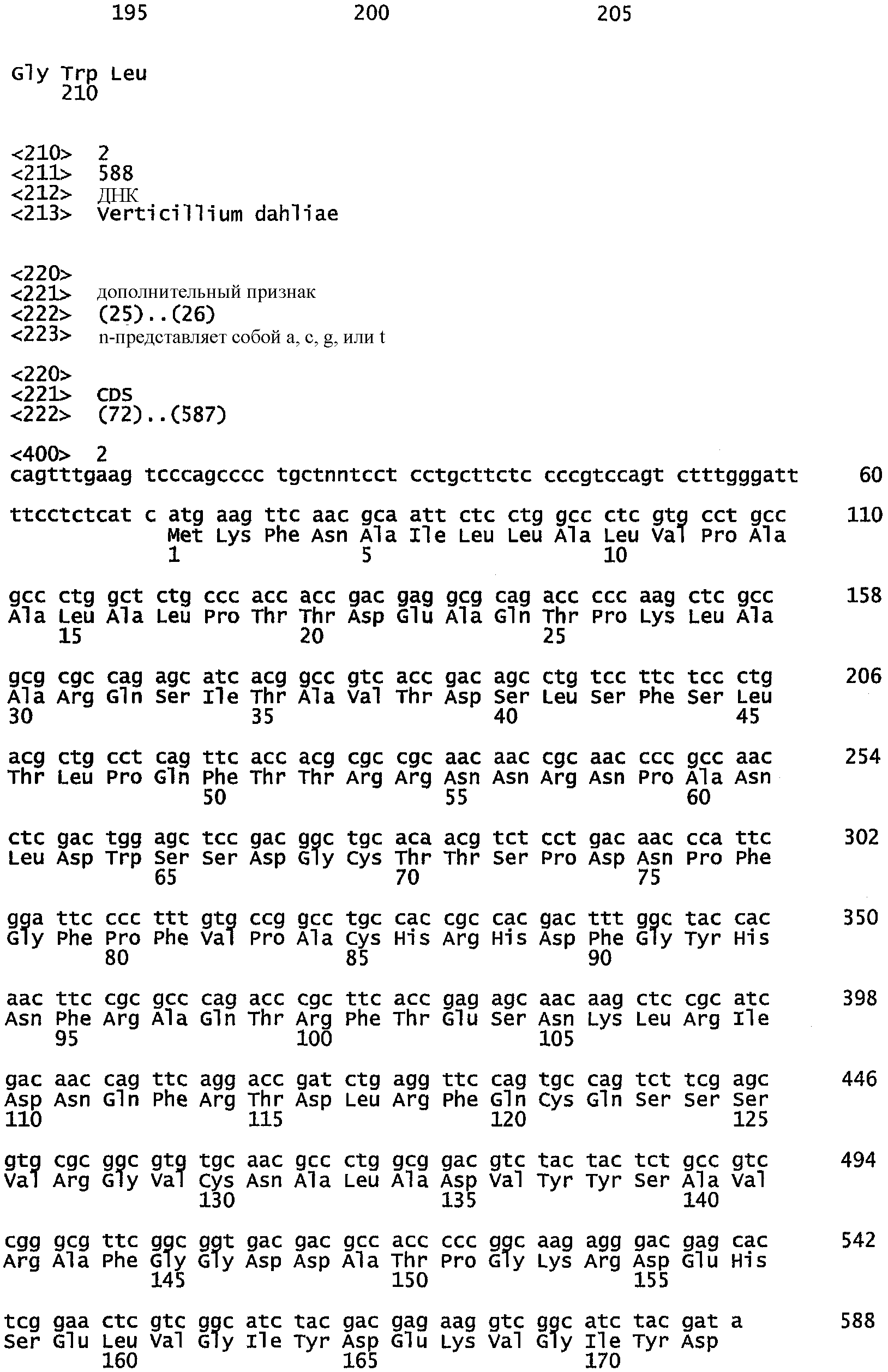
- COGEME Phytoраthogenic Fungi and Oomycete EST Database, Unisequence ID: VD0100C34, Vetricillium dahliae
- NCBI Protein database, gi: 18307435, Neurospora crassa
- NCBI Protein database, gi: 16519372, Helicosporum sp. NH1
- WO 0056762, SEQ ID NO: 5954, Aspergillus oryzae
- TREMBL Protein database, EAA28927, Neurospora crassa
В US 6399121 описывается использование фосфолипазы при получении сыра.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения проанализировали известные данные по последовательностям для грибных фосфолипаз А2 из группы XIII и идентифицировали дополнительные последовательности либо на основе данных, приведенных в опубликованных источниках по последовательностям, либо при скрининге соответствующих последовательностей из природных источников. Авторы показали, что при экспрессии генов, кодирующих грибные фосфолипазы А2 из группы XIII, в соответствующем организме-хозяине экспрессированные последовательности состоят из корового пептида, соединенного с пептидной последовательностью на N- или C-концевой стороне, или на обеих сторонах, при этом экспрессия гена в соответствующем организме-хозяине может вести к расщеплению экспрессируемого пептида с получением корового пептида без удлиняющего пептида, достигающего N- или C-конца. Они также показали, что коровый пептид, не включающий какого-либо пептидного удлинения пептида, обладает значительно более высокой активностью фосфолипаз, чем коровый пептид, соединенный с пептидным(ыми) удлинением(ями). И наконец, они показали, что найденный ими при использовании данного способа коровый пептид аналогичен по длине и последовательности известному зрелому пептиду из Helicosporum sp. (Wakatsuki, S. et al. (2001) Biochim. Biophys. Acta 1522: 74-81) с неизвестной функцией и к бактериальным фосфолипазам А2 из группы XIII, в которых отсутствуют пептидные удлинения, отличные от тех, которые относятся к сигналам секреции (Sugiyama, M. et.al. (2002) J. Biol. Chem. 277: 20051-20058).
Кроме того, авторы изобретения обнаружили, что фосфолипаза, обладающая сходством по последовательности активного сайта и по уровню консервативности по цистеиновому остатку с грибной фосфолипазой А2 из группы XIII, полезна для получения сыра.
Кроме того, авторы изобретения обнаружили и выделили ген, кодирующий новую фосфолипазу из Fusariumvenenatum A3/5, который был вначале депонирован как FusariumgraminearumATCC 20334, а затем классифицирован как Fusariumvenenatum (Yoder and Christianson, 1998, Fungal Genetics and Biology 23: 62-80 и O'Donnel et al., 1998, Fungal Genetics and Biology 23: 57-67). Данная фосфолипаза принадлежит к грибной/бактериальной группе XIII PLA2, в соответствии с определением Сораньи с соавт. (Soragni, E., et al., 20 (2001) EMBO J 20:5079-5090). Авторы изобретения также клонировали ген, кодирующий новую фосфолипазу, в штамме E. coli и затем использовали клонированный ген для создания конструкции, экспрессирующей ген фосфолипазы из Fusarium в Aspergillusoryzae. Авторы изобретения трансформировали Aspergillusoryzae данной конструкцией и выделили фосфолипазу из трансформированных клеток Aspergillus.
Соответственно, настоящее изобретение относится к способу получения фосфолипазы, который включает процессинг экспрессированного грибного пептида, позволяющий отщепить пептид от С-концевого конца и/или пептид от N-концевого конца с получением корового пептида, где коровый пептид включает:
а) аминокислотную последовательность, образованную аминокислотами 146-153 из SEQ ID NO: 1, аминокислотами 87-94 из SEQ ID NO: 3 или аминокислотами 79-86 из SEQ ID NO: 12, или последовательность, идентичную любой из указанных аминокислотных последовательностей, за исключением замещения одной аминокислоты другой аминокислотой; и
b) по меньшей мере два цистеиновых остатка, расположенных на N-концевом участке последовательности, приведенной в а); и
с) по меньшей мере два цистеиновых остатка, размещенных на С-концевом участке последовательности, приведенной в а).
Изобретение также относится к способу гидролиза фосфолипида фосфолипазой по настоящему изобретению. Кроме того, изобретение относится к способу получения сыра путем взаимодействия молока для сыроделия или фракции такого молока с фосфолипазой с последующим получением сыра из указанного молока для сыроделия.
И наконец, настоящее изобретение относится к фосфолипазе, которая является полипептидом с аминокислотной последовательностью, которая по меньшей мере на 80% идентична отдельным идентифицированным последовательностям.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Экспрессированный пептид
В практике осуществления настоящего изобретения используется экспрессированный грибной пептид, принадлежащий к группе, определяемой на основе сходства последовательностей активного сайта и уровня консервативности по цистеиновому остатку, как параметров, используемых при определении группы «фосфолипаза А2 из грибной/бактериальной группы XIII», в соответствии с предложением Сораньи с соавт. (Soragni, E., et al. (2001) EMBO J. 20:5079-5090). Пептид имеет грибную природу, то есть получен из Tuber, Verticillium, Neurospora, Helicosporum или Aspergillus, в особенности T. borchii, T. albidum, V. dahliae, V. tenerum, N. crassa, Helicosporum sp. HN1 или A. oryzae.
Пептид может обладать фосфолипазной активностью, например, активностью фосфолипазы А, такой как активность фосфолипазы А1 и/или активность фосфолипазы А2.
Некоторые конкретные примеры представляют собой известные пептиды с аминокислотными последовательностями, перечисленными ниже в перечне последовательностей. Ниже указаны организмы-источники и соответствующие литературные ссылки:
- SEQ ID NO: 1. Tuberborchii, Soragni, E., et al., (2001) EMBO J. 20:5079-5090;
- SEQ ID NO: 3. Vetricillium dahliae, COGEME Phytoраthogenic Fungi and Oomycete EST Database, Unisequence ID: VD0100C34;
- SEQ ID NO: 4. Neurospora crassa, NCBI Protein database, gi: 18307435;
- SEQ ID NO: 5. Helicosporum sp. NH1, NCBI Protein database, gi: 16519372;
- SEQ ID NO: 7. Aspergillus oryzae, WO 0056762, SEQ ID NO: 5954;
- SEQ ID NO: 8. Neurospora crassa, TREMBL Protein database, EAA28927.
Кроме того, авторами настоящего изобретения были выделены следующие грибные фосфолипазы с указанными ниже последовательностями, из природных источников, приобретенных из общественно доступных коллекций или собранных в указанной стране и в указанный год:
- SEQ ID NO: 10. Tuber albidum, Purshased from Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands, изолят CBS272.72;
- SEQ ID NO: 12. Vetricillium tenerum, Ireland, 1996.
Авторы осуществили встраивание гена из T.albidum (SEQ ID NO: 9) в Е. coli и депонировали клон в Коллекции на условиях Будапештского договора 12 февраля 2003 года. Депонирование было осуществлено в Коллекции микроорганизмов и клеточных культур в Германии (Deutsce Sammlung von Mikroorganismen und Zellkulturen (DSMZ), Mascheroder Weg 1b, D-38124 Braunschweig, Germany) и ему был присвоен депозитарный номер DSM 15441.
В одном варианте настоящее изобретение относится к фосфолипазе, которая является полипептидом с аминокислотной последовательностью, которая по меньшей мере на 80%, например, по меньшей мере на 85%, предпочтительно на 90%, более предпочтительно по меньшей мере на 95%, идентична аминокислотам на участке 91-210 в SEQ ID NO: 10 (T.albidum), аминокислотам на участке 92-211 в SEQ ID NO: 1 (T. borchii), аминокислотам на участке 30-137 в SEQ ID NO: 12 (V. tenerum), аминокислотам на участке 38-145 в SEQ ID NO: 3 (V.dahliae), аминокислотам на участке 44-151 в SEQ ID NO: 4 (N. crassa), аминокислотам на участке 37-157 в SEQ ID NO: 7 (A oryzae) или аминокислотам на участке 58-168 в SEQ ID NO: 8 (N. crassa).
Процессинг пептида
Анализируя последовательности фосфолипазы, приведенных в перечне последовательностей, авторы изобретения обнаружили, что каждая экспрессированная аминокислотная последовательность состоит из сигнального пептида, корового пептида и дополнительной пептидной последовательности с неизвестной функцией, присоединенной к С- или N-концу, или к обоим концам корового пептида.
Коровый пептид
Коровые пептиды характеризуются одинаковым сходством последовательности активного сайта и уровнем консервативности по цистеиновому остатку, найденным Сораньи с соавт. (Soragni, E., et al., (2001) EMBO J. 20:5079-5090), с фосфолипазой А2 из грибной/бактериальной группы XIII.
В предпочтительном варианте осуществления настоящего изобретения коровые пептиды включают: а) последовательность, образованную аминокислотами 146-153 в SEQ ID NO: 1, аминокислотами 87-94 в SEQ ID NO: 3 или аминокислотами 79-86 в SEQ ID NO: 12; или последовательность, идентичную любой из указанных аминокислотных последовательностей, за исключением замещения одной аминокислоты другой аминокислотой; и b) два цистеиновых остатка, расположенных на N-концевом участке последовательности, приведенной в а); и с) два цистеиновых остатка, расположенных на С-концевом участке последовательности, приведенной в а).
Один из цистеиновых остатков, расположенных на N-концевом участке последовательности, приведенной в а), может быть, например, отделен от последовательности, приведенной в а), 0-5 аминокислотами, например, 0-3 аминокислотами, предпочтительно 0-2 аминокислотами и еще более предпочтительно 1 аминокислотой. Другой из цистеиновых остатков, расположенных на N-концевом участке последовательности, приведенной в а), может быть, например, отделен от последовательности, приведенной в а), 14-20 аминокислотами, например, 15-19 аминокислотами, предпочтительно 16-18 аминокислотами и еще более предпочтительно 17 аминокислотами.
Один из цистеиновых остатков, расположенных на С-концевом участке последовательности, приведенной в а), может быть, например, отделен от последовательности, приведенной в а), 22-29 аминокислотами, например, 23-28 аминокислотами, предпочтительно 24-27 аминокислотами и еще более предпочтительно 25-26 аминокислотами. Другой из цистеиновых остатков, расположенных на С-концевом участке последовательности, приведенной в а), может быть, например, отделен от последовательности, приведенной в а), 27-49 аминокислотами, например, 29-46 аминокислотами, предпочтительно 30-43 аминокислотами, еще более предпочтительно 32-42 аминокислотами и наиболее предпочтительно 35-40 аминокислотами.
В предпочтительном варианте осуществления настоящего изобретения коровый пептид включает четыре цистеиновых остатка, сопоставляемых с цистеиновыми остатками в SEQ ID NO: 1, под номерами 128, 144, 180 и 194, соответственно, когда полная экспрессированная последовательность фосфолипазы сопоставляется (выравнивается) одновременно с последовательностями, приведенными в SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 10 и SEQ ID NO: 12.
Согласно настоящему изобретению, экспрессированный полипептид расщепляют, с тем чтобы отделить коровый пептид от присоединенного(ых) пептида(ов). Расщепление может быть осуществлено in vivo посредством экспрессии в клетке соответствующего нитевидного гриба-хозяина, или in vitro, например, при обработке соответствующей фосфолипазой, такой как Kex2.
Точки расщепления могут быть обнаружены в пределах 11 аминокислот в последовательности, которая представляет собой FG, или внутри 10 аминокислот в последовательности, которая представляет собой сайт Kex2. Сайты Kex2 представляют собой, например, RR, KR, KK или RK. В одном варианте осуществления изобретения коровый пептид имеет длину 100-150 аминокислот, например, 110-140 аминокислот, 115-133 аминокислоты, 118-129 аминокислот или 118-126 аминокислот.
В одном варианте осуществления настоящего изобретения экспрессированную фосфолипазу расщепляют на участке, включающем 0-18 аминокислот, таком как участок из 3-16 аминокислот, предпочтительно 5-14 аминокислот, на N-концевом участке последовательности, при сопоставлении с аминокислотами 97-101 в SEQ ID NO: 1, когда полная экспрессированная последовательность фосфолипазы сопоставляется (выравнивается) одновременно с последовательностями, приведенными в SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 10 и SEQ ID NO: 12.
В предпочтительном варианте осуществления настоящего изобретения экспрессированную фосфолипазу расщепляют на участке, включающем 0-11 аминокислот, таком как участок из 0-9 аминокислот, предпочтительно 0-7 аминокислот, на С-концевом участке последовательности, при сопоставлении с аминокислотами 204-209 в SEQ ID NO: 1, когда полная экспрессированная последовательность фосфолипазы сопоставляется одновременно с последовательностями, приведенными в SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 10 и SEQ ID NO: 12.
В предпочтительном варианте осуществления настоящего изобретения процессированная фосфолипаза обладает удельной фосфолипазной активностью, которая выше, чем активность экспрессированного пептида до процессинга, например, в одном варианте удельная активность фосфолипазы по меньшей мере в 2 раза, более предпочтительно по меньшей мере в 5 раз, более предпочтительно по меньшей мере в 10 раз превышает удельную активность фосфолипазы экспрессированного пептида до процессинга. В одном варианте осуществления настоящего изобретения экспрессированный пептид до процессинга не обладает измеряемой активностью фосфолипазы.
Активность фосфолипазы может быть, например, определена в тесте LEU при гидролизе соевого лецитина (L-альфа-фосфатидилхолин) при pH 8,0 и 40°С в течение 2 минут. Активность фосфолипазы выражают в виде относительного потребления титрующего агента (0,1 M NaOH), необходимого для поддержания постоянного pH, на фоне стандарта.
Экспрессия в клетках-хозяевах нитевидных грибов
Нитевидный гриб, взятый в качестве клетки-хозяина, может представлять собой, например Acremonium, Aspergillus, Fusarium, Humicola, Myceliophthora, Neurospora, Penicillium, Rhizomucor, Thermomyces, Thielavia, TolypocladiumилиTrichoderma, в особенностиA. awamori, A. foetidus, A. japonicus, A. nidulans, A. niger, A. oryzae, F. bactridioides, F. cerealis, F. crookwellense, F. culmorum, F. graminearum, F. graminum, F. heterosporum, F. negundi, F. oxysporum, F. reticulatum, F. roseum, F. sambucinum, F. sarcochroum, F. sporotrichioides, F. sulphureum, F. torulosum, F. trichothecioides, F. venenatum, H. insolens, M. thermophila, N. crassa, P. purpurogenum, R. miehei, Thermomyceslanuginosus, Thielaviaterrestris, Trichodermaharzianum, Trichodermakoningii, Trichodermalongibrachiatum, TrichodermareeseiилиTrichodermaviride.
В предпочтительном варианте организм хозяина представляет собой Aspergillus, Fusarium или Trichoderma, в особенности A. niger, A. oryzae, F. venenatum, F. sambucinum или F. cerealis.
Процедуры трансформации, культивирования, экспрессии и восстановления могут быть осуществлены традиционными способами, например, с использованием основных методик, описанных в EP 238023, EP 305216, WO 9600787, EP 244234 или в работе Кристенсена с соавт. (T. Christensen et al., BioTechnology, vol. 6, Dec. 1988, 1419-22).
Полипептид фосфолипазы и соответствующая ДНК
В одном варианте настоящее изобретение относится к полипептидам, обладающим фосфолипазной активностью, где указанные полипептиды включают аминокислотную последовательность (или предпочтительно состоят из нее), которая обладает идентичностью к аминокислотам 29-146 в SEQ ID NO: 16 (например, зрелому полипептиду) на уровне, равном по меньшей мере 80%, таком как по меньшей мере 85%, более предпочтительно по меньшей мере 90%, наиболее предпочтительно по меньшей мере 95%, например, по меньшей мере на уровне 96%, таком как по меньшей мере 97%, и еще более предпочтительно по меньшей мере 98%, таком как по меньшей мере 99%.
Предпочтительно полипептиды включают аминокислотную последовательность SEQ ID NO: 16; ее аллельный вариант или ее фрагмент, который обладает фосфолипазной активностью. В другом предпочтительном варианте полипептид по настоящему изобретению включает аминокислоты 29-149 из SEQ ID NO: 16. В еще одном предпочтительном варианте полипептид по настоящему изобретению состоит из аминокислот 29-146 из SEQ ID NO: 16.
Настоящее изобретение также относится к полинуклеотиду, который включает нуклеотидную последовательность (или предпочтительно состоит из нее), которая обладает по меньшей мере 80% идентичностью с нуклеотидами 133-495 в SEQ ID NO: 15. Предпочтительно нуклеотидная последовательность обладает идентичностью по меньшей мере на уровне 85%, такой как по меньшей мере 90% идентичность, предпочтительно по меньшей мере 95% идентичность, такой как по меньшей мере 96% идентичность, например, по меньшей мере 97% идентичность, еще более предпочтительно по меньшей мере 98% идентичность, например, по меньшей мере на уровне 99%, с нуклеотидами 133-495 в SEQ ID NO: 15. Предпочтительно нуклеотидная последовательность кодирует полипептид, обладающий фосфолипазной активностью.
Фосфолипаза может быть получена из штамма Fusarium, в особенности F. Venenatum, с использованием зондов, разработанных на основе последовательностей ДНК, приведенных в настоящем описании. В одном варианте фосфолипаза обладает активностью фосфолипазы А.
Фосфолипаза может быть получена путем трансформации соответствующей клетки-хозяина последовательностью ДНК, кодирующей фосфолипазу, с последующим культивированием трансформированного организма в условиях, обеспечивающих продуцирование фермента, и выделением фермента из культуры.
Организм хозяина представляет собой предпочтительно эукариотическую клетку, в частности грибную клетку, такую, как дрожжевая клетка, или клетку нитевидного гриба, такого как клетка штаммаAspergillus, Fusarium, TrichodermaилиSaccharomyces, в особенности A. niger, A. oryzae, F. venenatum, F. sambucinum, F. cerealis или S. cerevisiae, например, штамм A. niger, продуцирующий глюкоамилазу, такой как штаммы, описанные в US 3677902, или их мутанты. Продуцирование фосфолипазы в таком организме-хозяине может быть обеспечено при использовании основных методик, описанных в EP 238 023 (Novo Nordisk), WO 96/00787 (Novo Nordisk) или EP 244 234 (Alko).
Вектор экспрессии по настоящему изобретению обычно включает управляющие последовательности, функционирующие в качестве промотора, сигнала инициации трансляции и необязательно селектируемый маркер, терминатор транскрипции, ген-репрессор или различные гены-активаторы. Вектор может представлять собой автономно реплицирующийся вектор или он может быть интегрирован в геном хозяйской клетки.
Сопоставление последовательностей и определение идентичности
Сопоставление (выравнивание) нуклеотидных последовательностей может быть осуществлено с использованием приложения AlignX программы Vector NTI Program Suite 7,0 с использованием заданных по умолчанию установок, в которых используется модифицированный алгоритм ClustalW (Thompson, J.D., Higgins, D.G., and Gibson T.J. (1994) Nuc. Acid. Res. 22: 4673-4680), матрица swgapdnarnt score matrix, штраф за наличие гэпа 15 и штраф за продолжение гэпа 6,66.
Сопоставление аминокислотных последовательностей может быть осуществлено при использовании приложения AlignX программы Vector NTI Program Suite v8 с использованием заданных по умолчанию установок, в которых используется модифицированный алгоритм ClustalW (Thompson, J.D., Higgins, D.G., and Gibson T.J. (1994)), матрица blosum62mt2 score matrix, штраф за наличие гэпа 10 и штраф за продолжение гэпа 0,1.
В одном варианте осуществления настоящего изобретения сопоставление последовательностей и расчет балльных показателей гомологии производят с использованием метода Липмана-Пирсона (Lipman, D.J., and W.R. Pearson (1985) Rapid and sensitive protein similarity searches. Science 227: 1435-1441) с использованием таблицы весовых значений остатков PAM250 (Dayhoff, M.O., R.M. Schwartz and B.C. Orcutt (1978) A model of evolutionary change in proteins. In Dayhoff M.O. (ed.), Atlas of Protein Sequence and Structure. National Biomedical Research Foundation. Washington, D.C. Vol. 5. Suppl. 3: pp. 345-358) с использованием заданных по умолчанию установок в программе MegAlign v4.03, инсталированной в пакет прикладных программ Lasergene (DNASTAR Inc., 1228 South Park Street, Madison, Wisconsin 53715). Заданные по умолчанию установки включают параметр K-tuple, равный 2, штраф на наличие гэпа 4 и штраф на удлинение гэпа 12.
Гидролиз фософолипида
Практическое осуществление настоящего изобретения включает использование при гидролизе фосфолипида, такого как лецитин, цефалин или инозитид.
Изобретение может быть использовано по аналогии с имеющимися в технике процессами, путем замены фосфолипазы, например, в процессе производства хлебобулочных изделий (WO 0032758, WO 9953769), майонезов (GB 1525929, US 4034124) или при обработке растительного масла (US 5264367).
Использование фосфолипазы
Фосфолипаза по настоящему изобретению может использоваться в различных промышленных процессах, включающих фосфолипазы, например, как описано ниже.
Использование в хлебопечении
Фосфолипаза по настоящему изобретению может использоваться при изготовлении теста, хлеба и кексов, например, для повышения эластичности хлеба или кексов. Таким образом, фосфолипаза может использоваться в процессе хлебопечения, включающем добавление фосфолипазы к ингредиентам теста, перемешивание теста и выпечку теста, с получением хлеба. Указанный процесс может быть осуществлен по аналогии с процессом, описанным в US 4567056 или WO 99/53769.
Использование в детергентных материалах
Один из вариантов фосфолипазы может использоваться в качестве детергентной добавки, например, в концентрации (выражаемой в виде чистого ферментного белка) 0,001-10 (например, 0,01-1) мг на грамм детергента или 0,001-100 (например 0,01-10) мг на литр моющей жидкости.
Детергентная композиция по настоящему изобретению может быть, например, изготовлена в виде детергентной добавки, которая применяется при ручной или машинной стирке и которая включает композицию добавки для стирки, подходящую для предварительной обработки окрашенных материалов, и смягчающую композицию для полоскания, или может быть изготовлена в виде детергентной композиции, применяемой в домашнем хозяйстве для очистки сильно загрязненных поверхностей. В составе детергентной композиции для стирки данный вариант может быть эффективным для использования с целью удаления жирных пятен, для отбеливания и для чистки мебели. Детергентная композиция для стирки может быть изготовлена по методике, описанной в GB 2247025, WO 9901531 или WO 9903962.
Детергентная композиция по настоящему изобретению может быть, в частности, изготовлена в виде, подходящем для ручного или машинного мытья посуды, например, как описано в GB 2 247 025 (Unilever) или WO 99/01531 (Procter & Gamble). В композиции для мытья посуды данный вариант может быть эффективным для удаления жирных/масляных пятен, для предупреждения потемнения/обесцвечивания посуды и полимерных компонентов посуды сильно окрашенными компонентами и для избежания отложения мыла на посуде.
Другие варианты использования
Фосфолипаза по настоящему изобретению может использоваться для улучшения фильтруемости водного раствора или взвеси, получаемых из углеводов после обработки их фосфолипазой. Такая процедура особенно применима к раствору или взвеси, содержащих крахмальный гидролизат, в особенности гидролизат пшеничного крахмала, поскольку он труднее фильтруется и образует мутные фильтраты. Обработка может быть проведена по методике, аналогичной методике, описанной в EP 219 269 (СPC International).
Кроме того, фосфолипаза по настоящему изобретению может использоваться для частичного гидролиза фосфолипидов, предпочтительно лецитина, для получения улучшенных фосфолипидных эмульгаторов. Данный вариант применения описан в энциклопедии Ульмана (Ullmann's Encyclopedia of Industrial Chemistry (Publisher: VCH Weinheim (1996)), патент JP 2794574 и JP-B 6-087751).
Кроме того, фосфолипаза по настоящему изобретению может использоваться в способе получения корма для животных, который включает перемешивание фосфолипазы с кормовыми веществами и по меньшей мере с одним фосфолипидом. Указанная процедура может быть осуществлена по аналогии с методикой, описанной в EP 743 017.
В еще одном варианте фосфолипаза по настоящему изобретению может использоваться в способе снижения содержания фосфолипидов в пищевом масле, который включает обработку масла фосфолипазой, так что достигается гидролиз основной части фосфолипида, и отделение водной фазы, содержащей гидролизованный фосфолипид, от масла. Данный процесс применим к очистке любого пищевого масла, которое содержит фосфолипид, например, растительного масла, такого как соевое масло, рапсовое масло и подсолнечное масло. Фосфолипаза может, например, использоваться в способах, описанных в JP-A-2-153997 и US 5264367.
Способ получения сыра
Фосфолипаза по настоящему изобретению может использоваться для получения сыра по способу, аналогичному способу, приведенному в US 6399121.
В предпочтительном варианте осуществления настоящего изобретения сыр изготавливают при взаимодействии молока для сыроделия или фракции молока для сыроделия с фосфолипазой по настоящему изобретению с последующим получением сыра из молока для сыроделия.
В другом предпочтительном варианте осуществления настоящего изобретения сыр изготавливают при взаимодействии молока для сыроделия или фракции молока для сыроделия с фосфолипазой, где указанная фосфолипаза включает:
а) последовательность, приведенную на участке аминокислот 146-153 в SEQ ID NO: 1, аминокислот 87-94 в SEQ ID NO: 3, аминокислот 79-86 в SEQ ID NO: 12; или последовательность, идентичную любой из указанных аминокислотных последовательностей, за исключением замещения одной аминокислоты другой аминокислотой; и
b) два цистеиновых остатка, расположенных на N-концевой стороне последовательности, приведенной в а); и
c) два цистеиновых остатка, расположенных на С-концевой стороне последовательности, приведенной в а).
В контексте настоящего описания термин «молоко для сыроделия» следует понимать как относящийся к любой композиции на основе молока, используемой для производства сыра. Фракция молока для сыроделия может быть любой фракцией молока для сыроделия, такой как, например, сливки, сепарированное молоко, молоко, пахта, масло или молочный жир.
В предпочтительном варианте молоко для сыроделия или фракцию молока для сыроделия подвергают взаимодействию с фосфолипазой по настоящему изобретению в количестве, достаточном для снижения эффекта промасливания в сыре и/или для повышения выхода сыра. Эффект промасливания относится к тенденции сыра выделять масло при хранении и/или расплавлении.
В одном аспекте настоящее изобретения относится к способу изготовления сыра, включающему обработку молочной композиции фосфолипазой по настоящему изобретению, с последующим получением сыра из молочной композиции.
Другой аспект настоящего изобретения относится к способу изготовления сыра, включающему обработку молочной композиции фосфолипазой, с последующим получением сыра из молочной композиции, где фосфолипазу выбирают из грибных/бактериальных фосфолипаз PLA2 группы XIII. В предпочтительном варианте осуществления настоящего изобретения грибная/бактериальная PLA2 из группы XIII представляет собой фосфолипазу из гриба, более предпочтительно из гриба, относящегося к Ascomycetes. Фосфолипаза, принадлежащая к грибной/бактериальной PLA2 группы XIII, может быть любой фосфолипазой, принадлежащей к данной группе в соответствии с определением Сораньи с соавт. (Soragni, E., et al., (2001) EMBO Journal, 20:5079-5090), и может представлять собой фосфолипазу из любого вида Tube, например, T. borchii; Streptomyces, например, S. coelicor; Verticillium, например, V. dahliae; Aspergillus, например, A. oryzae; Neurospore, например, N.crassa или Helicosporum.
Молочная композиция по настоящему изобретению может представлять собой любую композицию, включающую компоненты молока. Компоненты молока могут быть такими компонентами молока, как молочный жир, молочный белок, казеин, белок сыворотки и лактоза. Молочная фракция может представлять любую фракцию молока, такую как, например, снятое молоко, пахта, сыворотка, сливки, молочный порошок, порошок цельного молока, порошок из снятого молока. В предпочтительном варианте осуществления настоящего изобретения молочная композиция включает молоко, снятое молоко, пахту, цельное молоко, сыворотку, сливки или любое их сочетание. В более предпочтительном варианте осуществления настоящего изобретения молочная композиция состоит из молока, такого как снятое молоко, цельное молоко, сливки, пахта или их любое сочетание.
Ферментативная обработка, осуществляемая в соответствии со способом настоящего изобретения, может проводиться путем диспергирования фосфолипазы в молочной композиции в условиях, позволяющих действие фермента, включающих соответствующее время выдерживания и подходящую температуру. Обработка фосфолипазой может проводиться в условиях, выбранных в соответствии с выбранным(ыми) ферментом(ами), на основе подходов, известных в данной области.
Ферментативная обработка может проводиться при любом подходящем pH, таком как, например, pH в диапазоне значений 2-10, таком как pH 4-9 или 5-7. В одном варианте обработку фосфолипазой проводят при температуре 3-60°С, такой как 25-45°С (например, в течение по меньшей мере 5 минут, в частности, в течение по меньшей мере 10 минут или по меньшей мере 30 минут, например, в течение 5-120 минут). Фосфолипазу добавляют в подходящем количестве, с тем чтобы достичь желательных свойств получаемого сыра. Предпочтительно фосфолипазу добавляют в количестве, эффективном для снижения эффекта промасливания сыра и/или для повышения выхода сыра. Подходящая дозировка фосфолипазы обычно укладывается в диапазон 0,001-0,5 мг ферментного белка на г молочного жира, предпочтительно в диапазон 0,01-0,3 мг ферментного белка на г молочного жира и более предпочтительно в диапазон 0,02-0,1 мг ферментного белка на г молочного жира.
Сыры, получаемые по способу настоящего изобретения, включают все виды сыра, такие как сыр кампезино, сыр честер, сыр данбо, сыр драбант, сыр херрегард, сыр манчего, сыр проволонский, сыр сан-полен, мягкий сыр, сыр шведский, сыр таледжо, белый сыр, включая творожно-сычужный сыр, сыр, получаемый при сычужном свертывании творога; зрелые сыры, такие как сыр чеддер, сыр колби, эдамский сыр, мюнстерский сыр, сыр грюйер, эмментальский сыр, сыр камамбер, сыр пармезан и сыр романо; голубой сыр, такой как датский голубой сыр; свежие сыры, такие как сыр фета; сыры, получаемые в процессе свертывания с добавлением кислоты, такие как сливочный сыр, сыр невшатель, сыр гуарг, домашний сыр и квезо бланко. В предпочтительном варианте настоящее изобретение относится к способу получения сыра паста филата, такого как, например сыр моцарелла и сыр пицца. Сыр паста филата, или тянущаяся сырная масса, обычно отличается уникальными пластифицирующими способностями и способностью поддаваться обработке путем проминания свежей сырной массы в горячей воде, которая придает готовому сыру его характерную волокнистую структуру и способностью к плавлению и растяжению (см., например, «Mozzarella and Pizza cheesе» by Paul S. Kindstedt, Chees: Chemistry, physics and microbiology, Volume 2: Mаjor Cheesе Groups, Second edition, page 337-341, Chapmаn & Hall).
Перечень последовательностей и депонированные микроорганизмы
Настоящее изобретение содержит информацию в виде перечня последовательностей, внесенного в описание, а также представленного на машиночитаемом носителе, прилагаемом к данной заявке. Кроме того, настоящее изобретение относится к депонированным микроорганизмам. Содержание машиночитаемого носителя данных и информации о депонированных микроорганизмах полностью включены в настоящее описание в качестве ссылки.
Депонирование биологического материала
Приведенный ниже биологический материал был депонирован в соответствии с условиями Будапештского договора в Коллекции микроорганизмов и клеточных культур в Германии (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmBH, Mascheroder Weg 1 B, D-38124 Braunschweig, Germany) и имеет следующие номера доступа:
МАТЕРИАЛЫ И МЕТОДЫ
Среды и субстраты
Среда YP+2%G
10 г дрожжевого экстракта
20 г пептона
вода до 1 литра.
Проводят автоклавирование при температуре 121°С в течение 20 минут, добавляют 100 мл 20% стерильного раствора глюкозы.
Среда RA для споруляции
50 г янтарной кислоты
12,1 г нитрата натрия
1 г глюкозы
20 мл 50х соли Фогеля (Davis, R.H and F.J. de Serres (1970), Meth. Enzymol. 17A:79-143)
Все компоненты перемешивают в одном литре дистиллированной воды и фильтруют в стерильных условиях.
Буфер Бриттона-Робинсона
0,023 М фосфорная кислота
0,023 М уксусная кислота
0,023 М борная кислота
Титруют с использованием NaOH или HCl до нужного pH.
Методы
Фосфолипазная активность (LEU)
Лецитин гидролизуют при постоянных значениях pH и температуры и определяют активность фосфолипазы как уровень потребления титрующего агента (0,1 N NаOH), добавляемого для нейтрализации высвобожденной жирной кислоты.
Субстрат представляет собой соевый лецитин (L-α-фосфатидилхолин) и условия обработки включают: pH 8,00, температура 40,0°С и длительность реакции 2 минуты. Величину активности в единицах определяют при сравнении со стандартом.
ПРИМЕРЫ
Пример 1: Экспрессия фосфолипазы А2 из Tuber albidum в Aspergillus oryzae
На основе последовательности ДНК, описанной Сораньи с соавт. (Soragni, E., et al., (supra)), разрабатывают праймеры для проведения реакции амплификации TbSP1 по методу ПЦР на основе геномной ДНК с введением на концы праймера соответствующих сайтов рестрикции для облегчения клонирования продукта ПЦР (SEQ ID NO: 13 и 14). Штамм Tuber albidum CBS 272.72 получают из Коллекции культур CBS в Нидерландах (Centraalbureau voor Schimmelkultures, Utecht, The Netherlands) и культивируют на X-агаре при 20°С, в соответствии с рекомендациями CBS, приведенными в перечне культур 1996 года. Отбирают мицелий с поверхности чашек Петри и выделяют суммарную ДНК с использованием набора FastDNA Spin Kit (BIO101, Inc., Vista, CA), в соответствии с инструкциями производителя. Амплификацию по методу ПЦР проводят с использованием основной смеси Extensor Hi-Fidelity PCR Master Mix (ABgene, Surrey, U.K.) в соответствии с инструкциями производителя и используют для отжига при температуре 52°С в течение первых 5 циклов и при 62°С в течение последних 25 циклов. Получают один продукт ПЦР и определяют последовательность, которая представлена в виде SEQ ID NO: 9, за исключением добавленных синтетических сайтов рестрикции. Сравнение данной геномной последовательности с последовательностью к ДНК, описанной Сораньи с соавт. (Е. Soragni, et al.), указывает на наличие единственного интрона. После удаления интрона нуклеотидная последовательность из T. albidum СBS272.72 становится на 92,5% идентична последовательности из T. borchii ATCC 96540, штамма, который был использован Сораньи с соавт. (Е. Soragni, et al.). Соответствующий пептид, предсказанный на основании генной последовательности из T. albidum СBS272.72, на 93,8% идентичен пептидной последовательности, сообщенной Сораньи с соавт. (Soragni et al.).
Фрагмент ПЦР подвергают рестрикции с использованием BamHI и XhoI и клонируют в векторе экспрессии Aspergillus pMStr57 с использованием стандартных методик. Вектор экспрессии pMStr57 содержит те же элементы, что и pCaHj483 (WO 98/00529) с небольшими модификациями, введенными в промотор Aspergillus Na2, и содержит последовательности, подходящие для селекции и размножения в E. coli, а также для селекции и экспрессии в клетках Aspergillus. Конкретно, селекцию в Aspergillus осуществляют по гену amdS из Aspergillusnidulans, который позволяет использовать ацетамид в качестве единственного источника азота. Экспрессия а клетках Aspergillus связана с вовлечением промотора модифицированной нейтральной амилазы II (NA2) из Aspergillusniger, который слит с 5'-лидерной последовательностью гена из Aspergillus nidulans, кодирующего триозофосфатизомеразу (tpi), и с терминатором из гена Aspergillusniger, кодирующего амилоглюкозидазу. Ген, кодирующий фосфолипазу А2, в полученной конструкции экспрессии Aspergillus, pMStr70 секвенируют и полученную последовательность сравнивают с последовательностью, определенной ранее для не клонированного фрагмента ПЦР, SEQ ID NO: 9. Единственная мутация Т на С была найдена на расстоянии 52 н.п. в направлении по ходу транскрипции от стоп-кодона.
Клетки Aspergillusoryzae трансформируют с использованием pMStr70 в рамках стандартных методик, описанных в работе Кристенсена с соавт. (T. Christensen et al., (1988), BioTechnology 6, 1419-22). Трансформанты культивируют в среде YP+2%G со встряхиванием при 275 об/мин при 30°С и отслеживают экспрессию фосфолипазы А2 из Tuber, TbPLA2, по результатам электрофореза в ДСН-ПААГ.
Характеристика белка
Анализ в ДСН-ПААГ выявляет два полосы, которые примерно соответствуют м.в. 25 и 16 кДа. Супернатант очищают ионообменной хроматографией на колонке с SР-сефарозой, уравновешенной 50 мМ ацетатным буфером, и проводят элюцию 1M NaCl, pH 5,0. Элюируют два белка в виде двух отдельных фракций. Концентрацию белка определяют с использованием набора Protein Assay ESL, поставляемого компанией Roche. Активность определяют в рамках теста LEU.
Белки подвергают секвенированию по N-концу. Было показано, что N-концевая последовательность пула 1 (полоса с м.в. 23 25 кДа) соответствует аминокислотам 32-50 в SEQ ID NO: 10. Блоттинг пула 2 (полоса с м.в. 16 кДа) выявляет две полосы с N-концевыми последовательностями, соответствующими аминокислотам 86-98 и 91-103, соответственно. Масс-спектральный анализ двух полос показывает массы, равные 13934 и 14348 Да соответственно, согласуясь с погрешностью 5 Да, с величинами, вычисленными на основании последовательностей аминокислот 86-210 и 91-210 в SEQ ID NO: 10, соответственно.
Пример 2: Процедура очистки двух форм T. albidum PLA2, экспрессированных в Aspergillus oryzae
Преимущественно при проведении ферментации в Aspergillus oryzae трансформанта из примера 1, который продуцирует T. albidumPLA2, выявляется две формы ферментов. Одна форма появляется в диапазоне 22-23 кДа, при анализе в ДСН-ПААГ, и соответствует пептиду, сообщенному Сораньи с соавт. (Soragni et al., (supra)). Дополнительно была обнаружена новая форма, которая выявляется в зоне 16-17 кДа при анализе в ДСН-ПААГ и которая обладает высокой удельной активностью и высокой изоэлектрической точкой.
Очистка пептида 22-23 кДа
Супернатант, получаемый при ферментации и содержащий фосфолипазу из T. albidum, экспрессируемую в клетках A. oryzae (полученную в примере 1), подвергают стерильному фильтрованию с использованием фильтра EKS, получаемого от компании Seitz Schenk Bad Kreuznach, Bettrongerstrasse 42, Germany D-73550, Waldstetten.
В стерильном профильтрованном супернатанте корректируют pH до значения 8 и ионную силу в районе 4 мСм.
Анионообменная хроматография
Первую стадию очистки проводят по методу анионообменной хроматографии на колонке с 50 мл сефарозы Fast Flow QTM, приобретаемой от компании Амершам Фармация (Amersham Pharmacia). Колонку подвергают предварительному уравновешиванию с использованием 50 мл Трис-ацетатного буфера, pH 8. Стерильный профильтрованный ферментированный бульон вносят на колонку, которую промывают тем же буфером до вымывания из нее несвязанного материала.
Связанные белки элюируют тем же самым буфером, pH 8, содержащим 1М хлорид натрия, со скоростью течения 5 мл/мин до получения конечного объема 500 мл. Собирают фракции по 5 мл каждая с использованием коллектора фракций и определяют активность фосфолипазы во всех фракциях по количественному методу с использованием лецитина в качестве субстрата и с использованием L-α-фосфатидилхолина, приобретаемого от компании Сигма (Sigma), продукт Р-5638, и активность анализируют с использованием набора NEFA C kit, приобретаемого от компании Вако (Wako Chemicals GmbH, Nissan Strasse 2, 41468 Neuss, Germany). Ниже описана подробная и точная процедура проведения анализа.
Растворы субстрата, содержащие 10 мг/мл лецитина в качестве субстрата, получают в различных буферах, таких как 50 мМ ацетатный буфер, pH 5, 50 мМ Hepes, pH 7, или 50 мМ Трис-ацетатный буфер, pH 9, содержащих 2 мМ CaCl2 и 0,1% Triton X-100, приобретаемых от компании Fluka chemicals. Затем субстрат эмульгируют при перемешивании и нагревании при температуре 50°С, охлаждают до 40°С и используют в качестве субстрата.
Количественное определение активности проводят с использованием 300 мкл эмульсии субстрата, инкубируемой с 25 мкл фракций фермента в течение 20 минут при 40°С, после чего 30 мкл реакционной смеси добавляют к 300 мкл красящего реагента А NEFA C, приготовленного в соответствии с инструкцией производителя, инкубируют в течение 10 минут при 37°С и затем добавляют к смеси 600 мкл красящего реагента В NEFA С и продолжают инкубировать данную смесь в течение 10 минут. Полученное синее окрашивание измеряют на спектрофотометре при длине волны 505 нм.
Характеристика белка
Объединяют фракции, содержащие ферментативную активность, и характеризуют по величине молекулярного веса при проведении электрофореза в ДСН-ПААГ с использованием предварительных изготовленных гелей Novix Pre casted, содержащих 4-20% Трис-глицина, получаемых от компании Invitrogen Life Technologies, Carlsbad CA 92008, USA.
Выявляют белок с М.в. 22-23 кДа, который подвергают блоттингу и анализу N-концевой последовательности с использованием секвенатора Applied Biosystem.
Определяют первые 19 аминокислотных остатков с N-конца, которые, как было показано, соответствуют последовательности аминокислот 32-50 в SEQ ID NO: 10.
Очистка пептида размером 16-17 кДа
Профильтрованный в стерильных условиях супернатант с экспрессированной в клетках A. oryzae фосфолипазой из T. albidum, корректируют до pH 4,7 и ионной силы до 4 мСм.
Катионообменная хроматография
Сефарозу SP-sepharoseTMfast flow приобретают от компании Амершам Фармация (Amersham Pharmacia). В колонку вносят 50 мл сефарозы и уравновешивают с использованием 50 мл ацетатного буфера, pH 4,7, затем на колонку вносят ферментационной супернатант и связанный материал вымывают тем же буфером.
Связанный белок с высоким значением pI элюируют в линейном градиенте соли с использованием 50 мМ ацетатного буфера, pH 4,7, содержащего 1 М хлорида натрия. В данной процедуре размер фракций и скорость течения аналогичны параметрам, использованным при характеристике формы фосфолипазы с низким pI. Активность фосфолипазы определяют количественно во фракциях с использованием набора NEFA, как указано выше. Фракции, содержащие активные фосфолипазы, объединят и проводят электрофорез в ДСН-ПААГ, как было описано выше.
Выявляют белок размером 16-17 кДа, который обладает высокой изоэлектрической точкой, выше 9.
Проводят анализ белка по N-концевой последовательности после блоттинга белка, секвенируют с использованием системы Applied biosystem, выявляя, что N-концевая последовательность полностью отличается от последовательности, сообщенной Сораньи с соват. (Soragni et al. suprа)). Таким образом, было показано, что PLA2 из T. albidum имеет две формы, полученные после разного N-концевого процессинга, с N-концевыми последовательностями, соответствующими аминокислотам на участках 86-105 и 91-110 в SEQ ID NO:10, соответственно.
Пример 3: Получение сыра с использованием фосфолипазы T. albidum
Используют пастеризованные не гомогенизированные сливки (North Carolina State University Dairy Plant) для стандартизации пятисот граммов пастеризованного, не гомогенизированного сепарированного молока (North Carolina State University Dairy Plant) до содержания жира 3,5%, нужного для получения сыра моццарелла с полным содержанием жира. Молоко для сыроделия, используемое в каждом эксперименте, обрабатывают либо фосфолипазой из T. albidum 16-17 кДа, приготовленной в соответствии с процедурой примера 2, либо коммерческим препаратом фосфолипазы Lecitase® 10L (Novozymes A/S, Bagsvаrd, Denmark), и помещают в водяную баню на 35°С до уравновешивания до заданной температуры. Используют исходный pH молока для сыроделия и добавляют 0,01 мас.% культуры закваски.
Отслеживают pH до достижения уровня 6,4. Разбавляют 250 мкл сычуга (Novozym 89L) деионизированной водой до общего объема раствора 9 мл, добавляют один мл данного раствора к молоку для сыроделия и молоко для сыроделия энергично перемешивают в течение 3 минут. Удаляют магнитную мешалку и сычужное молоко оставляют выстаиваться при 35°С.
После проведения указанных выше обработок сырная масса считается готовой для нарезания, если при введении в нее шпателя видны острые края среза. Сыр нарезают, проталкивая нож вниз, и, держа стакан, быстро поворачивают нож и выдергивают вверх. Сырную массу оставляют на 5 минут и затем осторожно перемешивают ложкой. Температуру поднимают до 41°С при перемежающемся осторожном перемешивании в течение примерно 40 минут или пока pH не упадет до 6,0-5,9. После проведения дренажа сырной массы с использованием специальной ткани ее затем заменяют в стакане и держат при температуре 41°С в водяной бане с удалением сыворотки, по мере необходимости.
Когда значение pH сырной массы достигает 5,3, чашку из нержавеющей стали, содержащую сырную массу, заливают в водяной бане при 69°C в течение 5 минут и затем растягивают руками. Сырную массу кондиционируют в холодной воде в течение 30 минут. Далее сырную массу сушат бумажным полотенцем, взвешивают и оставляют на холоду в течение ночи.
Проводят контрольные эксперименты по изготовлению сыра с той же самой партией молока и по тем же процедурам, за исключением добавления фосфолипазы.
Фактический выход сыра вычисляют на основании веса сыра после растягивания относительно общего веса молока для сыроделия.
Выход сыра после корректировки влажности выражают в виде фактического выхода, скорректированного по уровню стандартного постоянного содержания влаги. Выход, скорректированный по влажности, вычисляют путем умножения фактического выхода на коэффициент отношения фактического содержания влаги к стандартному уровню влажности в соответствии с формулой:
Yadj (Yact x 1-Mact)/(1-Mstd),
где Yadj=выход сыра, скорректированный по влажности, Yact=фактический выход сыра, Mact=фракция с фактической влажностью и Mstd=фракция со стандартной влажностью (0,48).
Выход сыра с корректированной влажностью во всех экспериментах и контролях показан в таблице 1.
Пример 4: Клонирование и экспрессия фосфолипазы (FvPLA2) из Fusariumvenenatum в Aspergillusoryzae
Клетки Fusariumvenenatum А3/5 (первоначально депонированные как Fusarium graminearum ATCC 20334 и недавно отнесенные к другому классу Fusariumvenenatum Иодером и Кристиансеном (Yoder and Christianson, 1998, Fungal Genetics and Biology 23: 62-80) и О'Донеллом с соавт. (O'Donnel et al., 1998, Fungal Genetics and Biology 23: 57-67) растят в течение двух дней в минимальной среде Фогеля (Davis, R.H and F.J. de Serres (1970), Meth. Enzymol. 17A:79-143) при 28°С в качалочной культуре, фильтруют через стерильный фильтр Miracloth (Calbiochem, San Diego, California, USA) и переносят в среду для споруляции RA, в которой их инкубируют в качалочной культуре еще в течение 24 часов при 28°С. Клетки и споры собирают центрифугированием и лизируют, после чего экстрагируют РНК и проводят транскрипцию в кДНК, которую клонируют в pZErO-2 в соответствии с методиками, описанными в WO 00/56762. Количество независимых клонов в данной библиотеке до амплификации составляло 2,5×105, в которой 92% содержали вставки, варьирующие по размеру от 550 до 2500 п.н. Частичные последовательности ДНК были определены примерно для 1000 случайным образом выбранных клонов и указанные последовательности были введены для хранения в компьютерной базе данных при использовании для обработки методик, описанных в WO 00/56762.
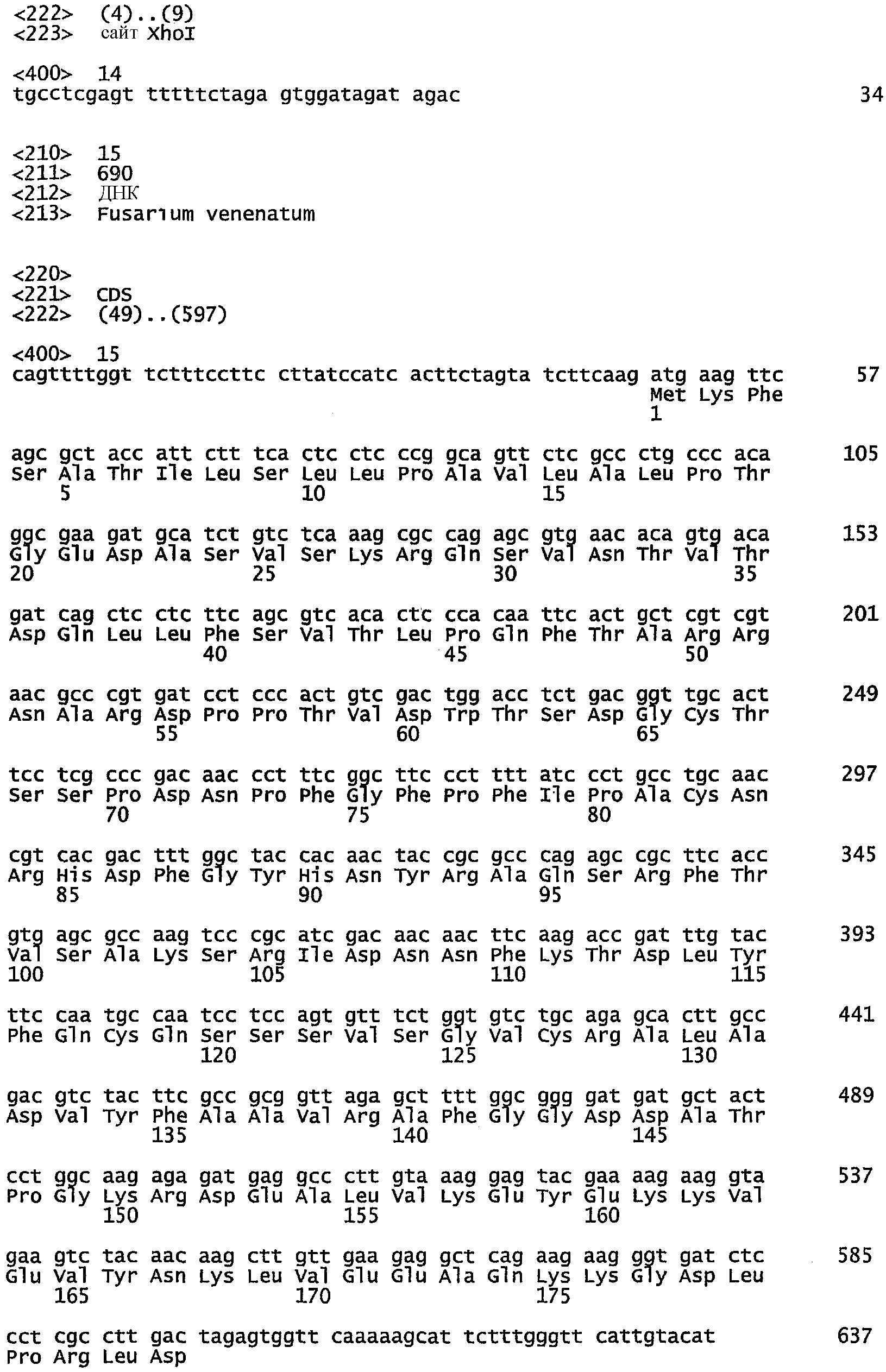
Нуклеотидная последовательность кДНК, кодирующая TbSP1, фосфолипазу A2 из Tuberborchii, и соответствующий пептид, полученный при ее трансляции, были сообщены Сораньи с соавт. (E. Soragni et al., 2001). Последовательность транслированного белка сравнивают с последовательностями белков, полученных при трансляции частичных последовательностей кДНК из Fusarium venenatum, с использованием программы TFASTXY, версия 3.3t08 (Pearson et al., 1997). Одна транслированная последовательность из F. venenatum была идентифицирована как обладающая 42% идентичностью к TbSP1 на перекрывающемся участке из 125 аминокислот. Была определена полная последовательность вставки кДНК из соответствующего клона FM0700, представленная в виде SEQ ID NO: 15, а последовательность пептида, транслированного на основе данной нуклеотидной последовательности, FvPLA2, показана в виде SEQ ID NO: 16. Указанная последовательность была использована для разработки праймеров FvPLA1 и FvPLA2.2 с целью применения в реакции амплификации по методу ПЦР гена из FM0700, кодирующего FvPLA2, с добавлением соответствующих сайтов рестрикции к концам праймера для облегчения субклонирования продукта ПЦР.
FvPLA1: CTGGGATCCTCAAGATGAAGTTCAGCG
FvPLA2.2: GACCTCGAGACCCGCCATTTAAGATТ
Амплификацию по методу ПЦР проводят с использованием основной смеси Extensor PCR Hi-Fidelity Master Mix (ABgene, Surrey, U.K.) в соответствии с инструкциями производителя и с использованием температуры отжига 52°С и температуры достраивания 60°С в течение 20 циклов.
Фрагмент, полученный в реакции ПЦР, подвергают рестрикции ферментами BamHI и Xhol и клонируют в векторе экспрессии pMStr57 для Aspergillus в соответствии со стандартными методиками. Вектор экспрессии pMStr57 содержит те же элементы, что и pCaHj483 (WO 98/00529) с небольшими модификациями, введенными в промотор Aspergillus Na2, как было описано для вектора pMT2188 в WO 01/12794, и включает последовательности, подходящие для селекции и размножения в E. сoli, а также для селекции и экспрессии в клетках Aspergillus. Конкретно, селекцию в Aspergillus осуществляют по гену amdS из Aspergillusnidulans, который позволяет использовать ацетамид в качестве единственного источника азота. Экспрессия в клетках Aspergillus вовлекает участие промотора модифицированной нейтральной амилазы II (NA2) из Aspergillusniger, который был слит с 5'-лидерной последовательностью гена из Aspergillusnidulans, кодирующего триозофосфатизомеразу (tpi), и с терминатором из гена Aspergillus niger, кодирующего амилоглюкозидазу. Ген, кодирующий фосфолипазу, в полученной конструкции экспрессии Aspergillus, pMStr70, секвенируют и получают последовательность, которая полностью соответствует последовательности, ранее определенной для вставки FM0700.
Штамм Aspergillus oryzae BECh2 (WO 00/39322) трансформируют с использованием pMStr77 в соответствии со стандартными методиками (T. Christensen et al., (1988). Трансформанты культивируют в среде YP+2%G на качалке со скоростью 275 об/мин при температуре 30°С и отслеживают экспрессию FvPLA2 проведением электрофореза в ДСН-ПААГ.
Штамм Escherichia coli, содержащий ген, кодирующий фосфолипазу из F. Venenatum, был депонирован авторами изобретения в соответствии с условиями Будапештского договора в Германской Коллекции микроорганизмов и клеточных культур (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmBH, Mascheroder Weg 1 b, D-38124 Braunschweig, Germany). Депонирование было осуществлено 12 февраля 2003 года под номером доступа DSM 15442.
Пример 5: Очистка и сравнение полученной последовательности FvPLA2
FvPLA2, полученную при ферментации в рамках процедуры примера 4, очищают ионообменной хроматографией на колонке с SP-сефарозой, уравновешенной 50 мМ ацетатным буфером, pH 4,7, и проводят элюцию 1M NaCl, pH 4,7. Фракции анализируют электрофорезом в ДСН-ПААГ и объединяют фракции, содержащие белок размером 14 кДа. Идентичность чистого белка подтверждают определением его N-концевой последовательности, которая идентична последовательности аминокислот 29-40 в SEQ ID NO:16. Дополнительно определяют массу белка при проведении масс-спектрального анализа, поскольку кажущийся размер, установленный при проведении электрофореза в ДСН-ПААГ, 14 кДа, меньше, чем соответствующая величина для пептида, предсказанная для случая процессинга теоретического пептида, в SEQ ID NO: 16. Было показано, что масса очищенного активного FvPLA2 составляет 13336 Да. Данная молекулярная масса указывает на дополнительный процессинг на С-конце, что согласуется с наличием расщепления между аминокислотами 149 и 150 в SEQ ID NO: 16, поскольку пептидная последовательность на участке аминокислот от 29 до 149 имеет теоретическую массу, равную 13335,66 Да.
Сравнение зрелого процессированного пептида (аминокислоты 29-149 в SEQ ID NO: 16) с известными последовательностями показывает, что ближайшая известная последовательность соответствует фосфолипазе из Verticillium dahliae, транслированной на основе последовательности Unisequence ID: VD0100C34 из базы данных по фитопатогенным грибам и оомицетам (COGEME Phytoраthogenic Fungi and Oomycete EST Database, Version 1.2 (http://cogeme.ex.ac.uk/) (Soanes et al., (2002) Genomics of phytopathogenic fungi and the development of bioinformatic resources. Mol Plant Microbе Interact. 15(5):421-7). Оценивают процессинг частичного пептида, предсказанного по данным о последовательности из V. dahliae, при сравнении с результатом процессинга FvPLA2. Расчеты показывают, что идентичность между аминокислотами 29-149 в SEQ ID NO: 16 и определенной последовательностью зрелого пептида фосфолипазы из V. dahliae составляет 77%.
Пример 6: Физические свойства FvPLA2
Каталитическая активность
Фосфолипазную активность как функцию концентрации фермента определяют в тесте LEU для FvPLA2 из примера 4. Результаты показаны в таблице 1.
Температурный профиль
Ферментативную активность как функцию температуры определяют для раствора фермента с концентрацией 5,3 мкг/мл. Другие условия соответствуют условиям, использованным в тесте LEU. Результаты приведены в таблице 2.
Стабильность в зависимости от pH
Фермент разбавляют в буфере Бриттона-Робинсона (Britton Robinsоn) при заданном pH в течение 30 минут при 30°С. После дальнейшего разбавления водой измеряют каталитическую активность в тесте LEU. Результаты показаны в таблице 3.
Термостабильность
Фермент разбавляют в буфере Бриттона-Робинсона (Britton Robinsоn) при заданных pH 3 и 10, соответственно, и при pH 7 с использованием 30% сорбита. После инкубации при заданной температуре в течение 30 минут раствор охлаждают до температуры реакционной смеси и анализируют в тесте LEU. Результаты показаны в таблице 4; активности приведены относительно наивысшей измеренной активности.
Пример 7: Изготовление сыра с использованием FvPLA2
Используют пастеризованные не гомогенизированные сливки (North Carolina State University Dairy Plant) для стандартизации пятисот граммов пастеризованного не гомогенизированного сепарированного молока (North Carolina State University Dairy Plant) до содержания жира 3,5%, получая при этом сыр моццарелла с полным содержанием жира.
Молоко для сыроделия, используемое в каждом эксперименте, обрабатывают либо фосфолипазой из F. venenatum (FvPLA2), полученной по процедуре примера 5, либо коммерческим препаратом фосфолипазы Lecitase® 10L (Novozymes A/S, Bagsvard, Denmark) и помещают в водяную баню при 35°С до уравновешивания до этой температуры. Используют исходный pH молока для сыроделия и добавляют 0,01 мас.% культуры закваски.
Отслеживают pH до достижения уровня 6,4. Разбавляют 250 мкл сычуга (Novozym 89L) деионизированной водой до общего объема раствора 9 мл, добавляют один мл данного раствора к молоку для сыроделия и молоко для сыроделия энергично перемешивают в течение 3 минут. Удаляют магнитную мешалку и сычужное молоко оставляют для выстаивания при 35°С.
После проведения указанных выше обработок сырная масса считается готовой для нарезания, если при введении в нее шпателя видны острые края среза. Сыр нарезают, проталкивая нож вниз, и, держа стакан, быстро поворачивают нож и выдергивают вверх. Сырную массу оставляют на 5 минут и затем осторожно перемешивают ложкой. Температура поднимается до 41°С при перемежающемся осторожном перемешивании в течение примерно 40 минут и пока pH не упадет до 6,0-5,9. После проведения дренажа сырной массы с использованием специальной ткани ее затем заменяют в стакане и держат при температуре 41°С в водяной бане с отливкой сыворотки, по мере необходимости.
Когда значение pH сырной массы достигает 5,3, чашку из нержавеющей стали, содержащуюся сырную массу, заливают в водяной бане при 69°C в течение 5 минут и затем растягивают руками. Сырную массу кондиционируют в холодной воде в течение 30 минут. Далее сырную массу сушат бумажным полотенцем, взвешивают и оставляют на холоду в течение ночи.
Проводят контрольные эксперименты по изготовлению сыра с той же самой партией молока и по тем же процедурам, за исключением добавления фосфолипазы.
Фактический выход сыра вычисляют на основании веса сыра после растягивания относительно общего веса молока для сыроделия.
Выход сыра после корректирования влажности выражают в виде фактического выхода, скорректированного по уровню стандартного постоянного содержания влаги. Выход, скорректированный по влажности, вычисляют путем умножения фактического выхода на коэффициент отношения фактического содержания влаги к стандартному уровню влажности в соответствии с формулой:
Yadj=(Yactx1-Mact)/(1-Mstd),
где Yadj=выход сыра, скорректированный по влажности, Yact=фактический выход сыра, Mact=фракция с фактической влажностью и Mstd = фракция со стандартной влажностью (0,48).
Выход сыра с корректированной влажностью во всех экспериментах и контролях показан в таблице 5.
Пример 8: Изготовление сыра с использованием FvPLA2
Молоко пастеризуют при 72°С в течение 15 секунд и затем охлаждают до температуры ниже 10°С. Молоко стандартизуют до содержания жира 2,4% с использованием сливок. После стандартизации молоко подвергают предварительному нагреванию в теплообменнике при температуре предварительного созревания жира 34,5°С. 150 кг молока вливают в каждый чан с изготавливаемым сыром и добавляют 15 г культуры (F-DVS ST-M6). Добавляют фосфолипазу, полученную по процедуре примера 5, в дозе 5 LEU/г жира и молоко инкубируют в течение 1 часа при температуре 34,5°С. Добавляют сычуг (Chy-Max Plus, 200 IMCU) и продолжают перемешивание в течение не более 4 минут.
После примерно 60 минут, когда видно, что произошло образование сгустка, его нарезают с использованием 10 мм ножей. Подводят смеситель к чану и через 10 минут начинают обваривание при повышении температуры до 40°С в течение 30 минут. После достижения температуры 41°С продолжают дополнительное перемешивание еще в течение примерно 20 минут до уровня титруемой кислотности 0,15-0,16%. Сырной массе позволяют осесть в чане и проводят ее дренирование. Сырную массу нарезают на равные блоки, которые переворачивают и складывают по два в стопку. Далее сырную массу измельчают в размалывающей машине при pH примерно 5,15-5,20. В кусочки сырной массы добавляют два процента соли (вес/вес).
После измельчения всю сырную массу вводят в устройство для растягивания, которое содержит 70 л уже нагретой до 74°С воды. Переносят в верхнюю камеру примерно 20 л горячей воды и добавляют туда сыр. Когда температура сырной массы достигает 62°С, останавливают растягивание и всю массу перемещают в экструдер. Сырную массу подвергают экструзии с получением 8-9 кусков сыра, каждый весом по 2,3 кг, которые охлаждают при температуре 5-7°С в течение 20 минут. Через 20 минут охлажденный сыр переносят в насыщенный соляной раствор и засаливают в течение 1,5 часа при температуре 5-6°С. Соляной раствор получают при перемешивании 120 кг воды, в которую добавлены соль до 22 Be, 750 г CaCl2 (34% раствор), и доводят pH до 5,1. После просаливания сыр сушат примерно в течение 30 минут и взвешивают перед проведением вакуумной упаковки. Отбирают образцы для определения pH и анализа состава (уровень влажности, содержание соли, жира и белка) после хранения в течение примерно 1 недели в холодной комнате.
Фактический выход (AY) корректируют до 48% влажности в сыре:
Adj Yield = AY x (100-% влажности) / 100-48
Пример 9: Сверхэкспрессия Aspergillus oryzae PLA2 (AoPLA2) в клетках Aspergillusoryzae
Среда
DAP2C-1
11 г MgSO4 х7H2O
1 г KH2PO4
2 г лимонной кислоты, моногидрат
30 г мальтодекстрина
6 г K3PO4 х3H2O
0,5 г дрожжевого экстракта
0,5 мл раствора микроэлементов металла
1 мл Плуроник РЕ 6100 (Pluronic PE 6100) (BASF, Ludwigshafen, Germany)
Компоненты смешивают в одном литре дистиллированной воды и порциями отбирают в колбы, добавляя 250 мг CаCO3 к каждой порции по 150 мл.
Среду стерилизуют в автоклаве. После охлаждения к 1 л среды добавляют следующие компоненты:
23 мл 50% (вес/объем) (NH4)2HPO4, подвергнутого стерильному фильтрованию
33 мл 20% молочной кислоты, подвергнутой стерильному фильтрованию
Раствор микроэлементов металла
6,8 г ZnCl2
2,5 г CuSO2 х5H2O
0,24 г NiCl2 х6H2O
13,9 г FeSO4 х7H2O
8,45 г MnSO4 хH2O
3 г лимонной кислоты, моногидрат
Компоненты смешивают в одном литре дистиллированной воды.
Клонирование и частичное секвенирование кДНК, кодирующей фосфолипазу А2 из Aspergillusoryzae, описаны в WO 00/56762. Полная последовательность клонов AS3812 показана в SEQ ID NO: 6. Данную последовательность используют для разработки праймера AoPLA1, используемого с векторным праймером pYESrev в реакции амплификации по методу ПЦР гена из AS3812, кодирующего PLA2, при добавлении сайта рестрикции для облегчения субклонирования ПЦР продукта:
AoPLA: TGAGGATCCATCATGAAGAACATCTTCG
pYESrev: gggcgtgaatgtaagcgtgac
Амплификацию по методу ПЦР проводят с использованием основной смеси Extensor PCR Hi-Fidelity Master Mix (ABgene, Surrey, U.K.) по инструкциям производителя и с использованием для отжига температуры 52°С в течение первых 5 циклов и температуры 60°С в течение 20 циклов и времени достраивания 1,5 минуты.
Фрагмент ПЦР подвергают рестрикции ферментами BamHI и Xhol и клонируют в векторе экспрессии Aspergillus pMStr57 (как описано в примере 1) в соответствии со стандартными методиками. В полученной конструкции экспрессии Aspergillus, pMStr71, ген, кодирующий фосфолипазу, секвенируют, выявляя при этом, что полученная последовательность полностью соответствует последовательности, ранее определенной для вставки AS3812.
Штамм Aspergillus oryzae BECh2 (WO 00/39322) трансформируют с использованием pMStr71, в соответствии со стандартными методиками (T. Christensen et al., (1988). Трансформанты культивируют в среде DAP2C-1 на качалке со скоростью 270 об/мин при температуре 37°С в течение 4 дней, оценивая экспрессию фосфолипазы по электрофорезу в ДСН-ПААГ.
Пример 10: Очистка и выявление процессинга пептида
Фосфолипазу Aspergillusoryzae после ферментации по процедуре примера 9 фильтруют через стерильный фильтр с размером 0,22 мкм SeitZ-EKS, получаемый от компании Pall Corporation (Pall SeitzSchenk Filter Systems GmbH Pianiger Str. 137 D-55543 Bad Kreuznach, Germany). Полученный после стерилизующего фильтрования раствор корректируют до pH 4,7 с использованием разбавленной уксусной кислоты. Далее корректируют ионную силу ферментационного супернатанта, так чтобы концентрация соли была низкой, а ионная сила ниже 4 мСм. Очистку желательного белка PLA2 проводят катионообменной хроматографией с использованием SP-сефарозы fast Flow matrix, от компании Амершам Фармация (Amersham-Pharmacia (Sweden)). Катионообменную матрицу вносят в колонку, промывают и подвергают предварительному уравновешиванию с использованием 50 мМ натрий-ацетатного буфера, pH 4,7 (буфер А) на колонке XK26 от компании Амершам Фармация (Amersham-Pharmacia). Супернатант, полученный после ферментации, который содержит искомую PLA2, доводят до соответствующих значений pH и ионной силы и затем наносят на колонку. Несвязанный материал смывают буфером А, промывая до тех пор, пока не выйдет весь УФ-поглощающий материал, по показаниям УФ-детектора, присоединенного к коллектору сбора фракций. Далее связанные белки элюируют линейным градиентом соли с использованием буфера В, который содержит 1М хлорид натрия, в качестве соли, в 50 мМ натрий-ацетатном буфере, pH 4,7. Общий объем линейного градиента, достигающий в итоге 1 М концентрации соли, составляет примерно 500 мл (10 объемов колонки). Собирают в ходе элюции фракции по 10 мл каждая. Все фракции анализируют на фосфолипазную активность с использованием в качестве субстрата лецитина от компании Сигма (Sigma). Жирные кислоты, высвобождаемые из лецитина при инкубации с фосфолипазой, определяют с использованием набора NEFA, от компании Waco. Фракции, содержащие фосфолипазную активность, проверяют на чистоту белка с использованием стандартной методики проведения электрофореза в ДСН-ПААГ. Объединяют фракции, которые содержат одну полосу желательного PLA2, демонстрирующую молекулярную массу около 16 кДа, которую определяют в сравнении с молекулярной массой стандарта, от компании Амершам Фармация (Amersham-Pharmacia).
Идентичность чистого белка подтверждают определением его N-концевой последовательности, которая была идентична последовательности аминокислот (АК) 37-45 в SEQ ID NO: 7. Дополнительно определяют массу пептида с помощью масс-спектрального анализа. Очищенный активный PLA2 из Aspergillus характеризуется двумя массами, 14114 и 14242 Да. Данные различающиеся молекулярные массы указывают на дополнительный процессинг на С-конце, соответствующий расщеплению между аминокислотами 121 и 122 в SEQ ID NO: 7, так как пептидная последовательность аминокислот 37-121 имеет теоретическую массу 14114,11 Да и расщепление между аминокислотами 122 и 123, что позволяет прогнозировать пептидную последовательность от 37 до 123 аминокислоты как имеющую теоретическую массу 14242,29 Да.
Пример 11: Экспрессия не полностью процессированной фосфолипазы из Aspergillus oryzae и Fusarium venenatum
Процессинг PLA2 из Aspergillusoryzae (AoPLA2) и PLA из Fusariumvenenatum (FvPLA2) на обоих, N- и C-концах, осуществляется по одному или многим основным остаткам (lys или arg), что типично для сайтов расщепления Kexin-подобных матураз, которые часто отвечают за процессинг пропептидов (Jalving, R., et al., (2000) Appl. Environ. Microbiol. 66: 363-368). Для определения влияния процессинга на активность AoPLA2 и FvPLA2 ферменты экспрессируют в Kеxin-дефицитном штамме Aspergillus oryzae. Далее оценивают процессинг по результатам проведения электрофореза в ДСН-ПААГ и определяют фосфолипазную активность в культурах штаммов, экспрессирующих AoPLA2 и FvPLA2, в варианте дикого типа и в Kexin-дефицитном штамме.
Кехin-дефицитный штамм Aspergillusoryzae (kexB-) конструируют путем разрушения гена kexB из A. oryzae (EMBL:AB056727) с использованием методик, известных в данной области техники, таких как методики, описанные в WO 98/12300 и US6013452. Разрушение kexB подтверждают при анализе по методу саузерн-блоттинга и отслеживают экспрессию пептидов, относительно которых известно, что kexB отвечает за их созревание. Штамм kexB- трансформируют экспрессирующей конструкцией AoPLA2, описанной в примере 9, и экспрессирующей конструкцией FvPLA2, описанной в примере 4. Данные штаммы ферментируют в среде YP+2%G при температуре 30°С, вместе со штаммами с kexB+ экспрессией и AoPLA2, и FvPLA2, как описано в примерах 9 и 4, используя нетрансформированные штаммы в качестве контроля. Штаммы, экспрессирующие AoPLA2, культивируют в качалочной культуре при 200 об/мин в течение 4 дней, а штаммы, экспрессирующие FvPLA2, культивируют в качалочной культуре при 275 об/мин в течение 3 дней. Экспрессию фосфолипазы и процессинг оценивают по результатам электрофореза в ДСН-ПААГ.
При анализе ДСН-ПААГ показано, что AoPLA2 разрешается в виде четкой единичной полосы в обоих штаммах: kexB+ и kexB-. При экспрессии в штамме kexB+ AoPLA2 демонстрирует свойства, соответствующие размеру 16 кДа, что согласуется с данными по его миграции, полученными ранее для полностью процессированного AoPLA2 (пример 10), тогда как в случае kexB- штамма AoPLA2 движется соответственно размеру примерно 27-28 кДА, что согласуется с фактом отсутствия процессинга или неполного процессинга. При экспрессии в штамме kexB+ FvPLA2 разрешается в виде двух полос с кажущимися молекулярными массами 17 кДа и 15 кДа. Полоса 14 кДа соответствует полностью процессированному пептиду (пример 5), тогда как пептид 17 кДа представляет собой частично процессированную форму. При экспрессии в штамме kexB- FvPLA2 движется в виде отдельной полосы размером примерно 18-19 кДа, и этот размер соответствует неполному процессингу. Не отмечается похожих полос в каком-либо из контрольных образцов из нетрансформированных штаммов. Относительные интенсивности полосы позволяют полагать, что экспрессия AoPLA2 в штамме kexB- составляет от 1/5 до 1/10 от ее уровня в штамме kexB+, тогда как экспрессия FvPLA2 в штамме kexB- была либо той же самой, либо достигала Ѕ от уровня в штамме kexB+.
Активность фосфолипаз, продуцируемых в каждом из штаммов, определяют в тесте LEU и их значения приведены в таблице 7.
Реферат
Изобретение относится к области биотехнологии и может быть использовано в пищевой промышленности, а также в производстве детергентных материалов. Клонирован ген новой фосфолипазы (FvPLA2) из Fusarium venenatum, принадлежащей к группе грибных и бактериальных фосфолипаз XIII PLA2. Определена нуклеотидная последовательность, кодирующая FvPLA2 и ее активную форму, путем экспрессии которой в гетерологичной системе получен рекомбинантный продукт, обладающий фосфолипазной активностью. Проведено исследование физико-химических свойств и показателей каталитической активности новой фосфолипазы, в соответствии с результатами которого предложено использование FvPLA2 в хлебопечении, производстве растительного масла, изготовлении сыра, а также в составе детергентных композиций. 8 н.п. ф-лы, 7 табл.
Формула
a) полипептид, кодируемый частью последовательности ДНК, клонированной в плазмиде, которая присутствует в Escherichia coli, депонированной под номером DSM 15442, и соответствующей нуклеотидам 133-495 последовательности SEQ ID NO: 15, или
b) полипептид с аминокислотной последовательностью, соответствующей аминокислотам 29-149 последовательности SEQ ID NO: 16, или аминокислотной последовательностью, которая может быть получена из нее путем консервативной замены одной или нескольких аминокислот, или
c) аналог полипептида, определенного в (а) или (b), с аминокислотной последовательностью, по меньшей мере на 90% гомологичной аминокислотной последовательности указанного полипептида.
а) часть последовательности ДНК, клонированной в плазмиде, которая присутствует в Escherichia coli, депонированной под номером DSM 15442, и соответствующей нуклеотидам 133-495 последовательности SEQ ID NO: 15, или
б) аналог последовательности согласно (а), который по меньшей мере на 90% идентичен указанной последовательности или гибридизуется в жестких условиях с комплементарной цепью указанной последовательности ДНК.
Комментарии