Применение клеточного экстракта одной или более микроскопических водорослей рода amphidinium для его фунгицидной и/или бактерицидной активности в отношении грибов, оомицетов и/или патогенных бактерий растений и семян культур - RU2762235C2
Код документа: RU2762235C2
Чертежи



Описание
Изобретение относится к области противогрибковых и антибактериальных средств для семян.
Для удовлетворения потребностей в связи с ростом населения во всем мире период после Второй мировой войны ознаменовался наступлением «Зеленой революции» - ряда современных программ, относящихся к селекции сортов, мелиорации, удобрениям и синтетическим пестицидам, цель которых состояла в контроле плодородия почвы и патогенных организмов. Эти различные элементы, таким образом, дали возможность на протяжении сорока с небольшим лет почти утроить мировое производство пищевых продуктов. Сегодня задача сельского хозяйства состоит в том, чтобы к 2050 году обеспечить питанием 9 млрд. человек и продолжать наращивать производство на единицу площади в ситуации постоянно возрастающего ограничения ресурсов и повышения требований. Например, потенциальные потери производства зерновых в связи с биотическими стрессами и отсутствием способов защиты сельскохозяйственных культур могут доходить более чем до половины. Традиционные препараты для защиты растений, предназначенные для ограничения потерь, таким образом, составляют неотъемлемую часть защиты сельскохозяйственных культур во всем мире. Однако эти химические вещества оказывают сильное отрицательное воздействие на здоровье человека и окружающую среду, что приводит к использованию других средств контроля инфекционных заболеваний, таких как биоконтроль (все способы защиты растений, использующие природные механизмы). Цель данного патента состоит в разработке одноклеточных водорослей, выделенных из фитопланктона, в качестве источника новых природных молекул, способных действовать в качестве «органического пестицида», непосредственно влияющего на выживаемость фитопатогенных организмов, инфицирующих культуры, имеющие важное значение в агрономии, такие как пшеница и виноградная лоза.
Фузариозные болезни
В Европе потери урожайности и снижение качества зерна, влияющее на здоровье, обусловлены несколькими болезнями, поражающими пшеницу (Triticum aestivum). Одной из важнейших является септориозная пятнистость листьев (Septoria spp.). Фузариозный вилт связан с комплексом биологических видов, состоящим из двух родов фитопатогенных грибов - Fusarium и Microdochium (1). Эти два рода включают около 19 видов, способных вызывать красную фузариозную гниль пшеницы. Наиболее распространенными в Европе видами являются F. graminearum, F. culmorum, F. avenaceum, F. poae, M. nivale и M. majus. Род Fusarium относится к отделу аскомицетов (лат. Ascomycota) и к семейству Nectriaceae. Род Microdochium относится к семейству Tuberculariaceae и включает два вида, М. nivale и М. majus, вызывающие такие же симптомы на колосе и листьях, как и Fusarium. В заданном регионе, на заданном участке земли или на одном колосе можно одновременно обнаружить несколько видов рода Fusarium, из которых наиболее распространенным представителем является Fusarium graminearum; таким образом, образуется комплекс Fusarium. Выраженность поражения, заболеваемость и распространенность каждого вида изменяются в зависимости от географического положения, изменений климата и практики выращивания сельскохозяйственных культур. Вероятно, присутствие нескольких из этих видов на одном и том же колосе модифицирует их равновесие и динамику образования токсинов.
Красная фузариозная гниль пшеницы может полностью уничтожить культуру за несколько недель до сбора урожая. Она может сопровождаться высокими потерями урожайности (недозрелость и низкая масса зерна), снижением всхожести семян или снижением качества за счет наличия токсинов в зернах. Действительно, грибы рода Fusarium, но не рода Microdochium, способны вырабатывать токсические вторичные метаболиты микотоксины, наличие которых приводит к повышению заболеваемости сельскохозяйственной продукции и создает значительные проблемы для экономики и здравоохранения. Основные средства борьбы с фузариозным вилтом включают практику выращивания сельскохозяйственных культур, разработку устойчивых сортов и химические методы борьбы. В настоящее время несколько сортов пшеницы устойчивы к фузариозному вилту. Однако имеются толерантные сорта с уровнями частичной устойчивости, которые ограничивают потери урожайности и накопление токсинов в урожае. Химические методы борьбы можно применять сразу после посева культуры, но они недостаточно эффективны. Эти методы борьбы осложняются вследствие разнообразия патогенных организмов и их различной чувствительности к активным веществам. Например, грибы рода Fusarium чувствительны к триазолам, тогда как грибы рода Microdochium чувствительны к стобилуринам.
Септориозные болезни
Септориозная пятнистость представляет собой болезнь пшеницы, которая ответственна за значительные потери урожая и вызывает наибольшие экономические потери во всем мире, особенно в регионах с влажным умеренным климатом. Можно различить две основные формы септориозной пятнистости пшеницы: септориоз колосковой чешуи (Phaeosphaeria nodorum) и пятнистость листьев (Mycosphaerella graminicola). Во Франции септориоз колосковой чешуи, в основном, присутствует в континентальных областях, тогда как пятнистость листьев, в основном, присутствует в северо-западных и прибрежных регионах, где климатические условия, по-видимому, благоприятны для развития гриба. Симптомы, вызываемые М. graminicola, появляются последовательно - в виде светло-зеленых пятен хлороза, которые затем изменяются до буроватых пятен, называемых некротическими очагами. Эти некротические очаги в конечном счете сливаются (коалесценция). Затем на этих некротических очагах появляются пикнидии - черные плодовые тела, едва заметные невооруженным глазом. Несколькими группами исследователей была изучена вредоносность септориозной пятнистости в отношении утраты способности к фотосинтезу, ослабления роста или потерь урожайности. Таким образом, можно было установить качественный показатель вредоносности, выражающий влияние болезни на содержание белка в собранных зернах.
М. graminicola является гемибиотрофным грибом, у которого при инфицировании живой ткани устанавливается первая биотрофная фаза, за которой следует некротрофная фаза, во время которой гриб экспрессирует токсины, вызывающие гибель колонизированных тканей. В зависимости от условий окружающей среды М. graminicola размножается либо половым (образование аскоспор), либо бесполым (образование пикнидоспор) путем. Аскоспоры, переносимые ветром на большие расстояния, вносят вклад в выживание гриба в отсутствие растения-хозяина и рассматриваются как основной источник первичного заражения для начала болезни. Пикнидоспоры, в свою очередь, в основном образуются в ходе эпидемической фазы болезни в течение нескольких последовательных инфекционных циклов. Эти споры распространяются вспышками на короткие расстояния под действием капель дождя. Будущий урожай снижается тем больше, чем сильнее поражаются болезнью последние листья под колосом, вовлеченные в налив зерна. Потери урожайности из-за септориозной пятнистости по оценкам составили в среднем 1-2 т.га-1, при этом случаи снижения урожайности вплоть до 3-3,5 т.га-1 составляют 40%.
Способы борьбы с М. graminicola основаны на применении фунгицидов и создании устойчивых сортов. Однако в последние годы наблюдается значительное снижение эффективности фунгицидов в связи с сильным действием отбора патогенных организмов, устойчивых, например, к семейству стробилуринов, а также недавнее снижение эффективности триазолов в полевых условиях.
Болезни виноградной лозы
В настоящее время виноградную лозу выращивают во всем мире, и она играет центральную роль в экономике многих стран. Виноград употребляют в виде столового винограда и сока, но его основной областью применения остается виноделие. Европейский Союз является крупнейшим производителем вина и крупнейшим экспортером винной продукции в мире. Вклад этого сектора в экономику Европейского Союза, таким образом, составляет около 15 млрд. евро в год (www. ceev.be). В 2010 г. французские виноградарские хозяйства охватывали почти 865000 гектаров или почти 3% пахотной земли, поэтому Франция является ведущим производителем вина во всем мире, производящим 51,1 млн. гектолитров. Виноградные лозы должны справляться с атаками множества патогенных организмов, в том числе с криптогамными болезнями. В случае поражения ими деревянистых частей растения их называют «болезнями древесины», конкретными примерами которых являются эска, некроз побегов винограда и эутипиоз. Грибы, поражающие ягоды и травянистые части виноградной лозы (листья, стебли и т.д.), вызывают «криптогамные болезни листвы», включающие серо-бурую гниль, черную гниль, ложную мучнистую росу и настоящую мучнистую росу.
Эска
Хотя тремя основными криптогамными болезнями, поражающими виноградные лозы во всем мире, являются ложная мучнистая роса, настоящая мучнистая роса и серо-бурая гниль, ограничивающими факторами в производстве винограда становятся болезни древесины, вызванные грибковыми агентами. В настоящее время виноделы сталкиваются с двумя основными проблемами, касающимися этих болезней древесины: нехваткой способов борьбы и значительной нехваткой сведений о различных биотических и абиотических факторах.
Наиболее распространенными видами грибов во всем мире, вызывающими болезнь эска, являются аскомицеты Diplodia seriata, Diplodia mutila, Neofusicoccum parvum и Neofusicoccum luteum. Во Франции наиболее изолированными видами являются Diplodia seriata и Botryosphaeria dothidea. Многие другие грибы, включая некоторые патогенны, часто выделяют из некроза древесины у растений, пораженных болезнью эска. Таким агентом, вызывающим эутипоз, например, является гриб Eutypa lata. Эта болезнь имеет две формы: медленную форму и апоплектическую форму. Для медленной формы характерны листовые симптомы, хотя они могут присутствовать и при апоплектической форме. Медленная форма характеризуется специфичной окраской листьев: желтоватыми междужилковыми пятнами на белых сортах винограда и красно-бордовыми междужилковыми пятнами на черных сортах винограда, при этом жилки остаются зелеными. Эти пятна постепенно изменяют свой цвет на буроватый и высыхают. Листовые симптомы медленной формы могут быть видны на посадочном материале виноградной лозы в течение года и исчезают на следующий год. Для апоплектической формы характерно быстрое полное или частичное высыхание надземных органов, ветвей, листьев и гроздьев виноградной лозы. Этот симптом обычно возникает в условиях жаркого лета, приводя к гибели виноградных лоз всего лишь за несколько дней без предупреждающих симптомов. Разнообразие источников заражения и очень медленный и невидимый рост грибов на древесине виноградной лозы в значительной степени затрудняет осуществление способов борьбы. Кроме того, в результате изменений нормативного регулирования препаратов для защиты растений на европейском уровне химические вещества на основе арсенита натрия были запрещены в связи с канцерогенными действиями на человека и высокой токсичностью этих препаратов для окружающей среды. Во всем мире проводятся обширные исследования по испытанию новых молекул, которые можно применять в питомниках или виноградарских хозяйствах.
Серо-бурая гниль
Серо-бурая гниль является криптогамным заболеванием, вызываемым грибом-аскомицетом под названием Botrytis cinerea. Он принадлежит к классу леоциомицетов, порядку Helotiales и семейству Sclerotiniaceae. В. cinerea является некротрофным грибом, способным колонизировать как здоровые, уже инфицированные ткани растений, так и мертвые ткани (сапрофитизм). На листьях симптомы появляются в виде бурых пятен с сероватым войлочным налетом на нижней стороне (образование плодовых тел гриба), которые склонны к разрастанию и инвазии всей листовой пластинки. Грозди могут поражаться перед цветением и отсыхать. Они особенно чувствительны на стадии начала созревания винограда, где наблюдается развитие бурой окраски ягод белых сортов винограда и появление толстого серого войлочного налета. Конидии распространяются ветром и проникают в травянистые органы непосредственно или через повреждения. Именно поэтому лопание ягод под действием ложной мучнистой росы способствуют инфекциям В. cinerea. Эта болезнь вызывает не только потери урожая вплоть до 40% (Viniflhor, данные 2006 г.), но также ухудшает органолептические качества вин. Тем не менее, Botrytis cinerea также вызывает «благородную гниль» винограда, необходимую для получения некоторых сладких вин.
Ложная мучнистая роса
В настоящее время двумя заболеваниями, тяжелее всего поражающими виноградники, являются ложная мучнистая роса и настоящая мучнистая роса. Агент, вызывающий ложную мучнистую росу, представляющий собой оомицет Plasmospora viticola, относящийся к порядку Peronosporales, является облигатным паразитом; чтобы выживать и сохранять многочисленность, он обязательно должен размножаться на живых виноградных листьях. P. viticola поражает все травянистые ткани виноградной лозы, а также гроздья. Он вызывает опадение листьев, побурение и высыхание ягод и стеблей. В отсутствие обработки и при благоприятных климатических условиях ложная мучнистая роса может уничтожить вплоть до 75% сезонного урожая.
Жизненный цикл P. viticola включает фазу полового и бесполого размножения. Фаза бесполого размножения ведет к образованию спор, необходимых для вторичных инфекций и для распространения патогенного организма на короткое расстояние, тогда как в фазе полового размножения образуются неподвижные и холодостойкие ооспоры, обеспечивающие перезимовку и первичные инфекции. Первым признаком поражения виноградника ложной мучнистой росой на макроскопическом уровне является появление бледно-желтых, неправильной формы пятен (маслянистых пятен), распространяющихся на верхней, или адаксиальной, поверхности листьев. По мере прогрессирования внутренней колонизации мицелия, развитие белой мшистой поросли на нижней стороне, соответствующей маслянистым пятнам, становится более выраженным. При развившихся инфекциях этим симптомам сопутствует бурая погибшая ткань. С ложной мучнистой росой, в основном, борются профилактическими мерами, используя опрыскивание фунгицидами. Хотя поражение возможно остановить, уже причиненный вред соцветиям и гроздьям необратим.
Настоящая мучнистая роса
Настоящую мучнистую росу винограда (Erysiphe necator) вызывает облигатный биотрофный аскомицет, относящийся к порядку Erysiphales. Этот гриб колонизирует поверхность всех зеленых органов виноградной лозы, особенно верхнюю поверхность листьев, и распространяется на ягоды. Фаза полового размножения, характеризующаяся образованием клейстотециев, содержащих аскоспоры, может чередоваться с фазой бесполого размножения, ведущей к образованию конидиофор, несущих конидии. Во время фазы зимнего покоя виноградной лозы гриб выживает в виде гиф в спящих почках или в виде клейстотециев на поверхности растения. Споры, содержащиеся в клейстотеции, высвобождаются весной для прорастания на поверхности почек и молодых листьев. Первичная гифа развивается на поверхности листа, затем она все больше усложняется и разветвляется, формируя мицелиальную сеть, покрывающую поверхность листа. Впоследствии в начале стадии споруляции из мицелия дифференцируются конидиоспоры и колонизируют другие зеленые ткани растения, приводя к вторичным инфекциям.
Присутствие на поверхности инфицированных тканей растения-хозяина мицелия и конидиофор, несущих конидии, создает внешний вид серовато-белой пудры. На цветочных почках развивается белый войлочный налет, и они высыхают. Заражению настоящей мучнистой росой подвержены только молодые ягоды с содержанием сахара менее 8%. Инфекции могут быть подвержены все поверхности листьев независимо от их возраста. Молодые инфицированные листья сначала становятся темно-зелеными, а затем деформируются и останавливаются в развитии. На верхней поверхности листьев могут находиться более светлые хлоротические пятна, подобные маслянистым пятнам ложной мучнистой росы. В настоящее время основные средства борьбы с болезнями, сильнее всего поражающими виноградники, составляет применение больших количеств пестицидов и фунгицидов. В виноградарстве нагрузка санитарно-гигиенических мер особенно сильна.
Для предотвращения вреда, вызванного возникновением болезни, обработки фунгицидами, предназначенные для борьбы, в основном, с ложной мучнистой росой и настоящей мучнистой росой, проводят по точному графику. В Европейском Союзе (ЕС) для борьбы с болезнями виноградной лозы применяют около 68000 тонн фунгицидов в год, что составляет 65% фунгицидов, применяемых в сельском хозяйстве, при этом виноградниками занято лишь 3,3% полезной сельскохозяйственной площади ЕС (Eurostat, 2007). В целях ограничения сильной химической нагрузки на окружающую среду и здоровье необходимо выделять молекулы природного происхождения, которые сыграют роль в защите сельскохозяйственных культур от инфекционных заболеваний и в конечном счете заменят химические препараты для защиты растений, используемые в настоящее время.
Парша яблок
Наряду с бурой гнилью и настоящей мучнистой росой парша является одной из основных грибковых болезней яблонь (род Malus). Ее вызывает гриб-аскомицет, называемый Venturia inaequelis, у которого существует несколько тысяч штаммов, вызывающих черные или бурые очаги поражения на поверхности листьев, почек или плодов, а иногда даже на древесине. Этой болезни особенно подвержены плоды и нижние части листьев.
Гриб зимует на листьях, опадающих с инфицированных деревьев, в форме перитециев. Весной, когда раскрываются почки, перитеции наполнены аскоспорами. Во влажные дни аскоспоры вылетают в воздух сада и попадают на деревья посредством перемещения по воздуху. Этот выброс аскоспор начинается при раскрытии почек и продолжается в течение 6-10 недель, чаще всего до конца июня. Аскоспоры, попадая на листву, когда листья в течение некоторого периода времени влажные, прорастают и проникают в листья, что приводит в результате к первичной инфекции. В зависимости от условий температуры и влажности грибковая инфекция становится заметной на различных частях дерева в течение от одной до трех недель. На листьях появляются темно-оливковые или бурые пятна размером около 5 мм, которые иногда могут целиком покрывать лист. Инфицированные цветки могут опадать. Инфекцию плодов идентифицируют прежде всего по серым пятнам на стебле.
После первичной инфекции и в течение остального летнего периода гриб развивается и образует конидии, которые являются другой формой репродуктивной структуры. При отделении конидий происходит вторичная инфекция. Конидии могут инфицировать любую часть дерева, и те из них, которые образовались в поздний летний период, могут расти даже на плодах, помещенных на хранение. Конидии распространяются при сильном дожде.
Эта болезнь в редких случаях уничтожает растение-хозяина, но в отсутствие обработки фунгицидами может значительно снижать (вплоть до 100%) качество и продукцию плодов. После превентивных мер, заключающихся в сборе опавших листьев в осенний период, стратегия борьбы требует эффективных действий весной по предотвращению инфекции или развития на деревьях высвобождаемых спор. Традиционный способ защиты состоял в том, чтобы начать нанесение фунгицидов при раскрытии почек и повторно наносить их каждые семь дней или до конца июня для защиты молодых побегов. Яблоневые сады больше всего обрабатывают фунгицидами и инсектицидами, и из всего количества обработок фунгицидами в год, составляющего в среднем 28,8, 19 приходятся на яблони (по данным Французского национального института сельскохозяйственных исследований).
Микроскопические водоросли
Молекулы природного происхождения, обладающие новым механизмом действия и способные преодолеть устойчивость, развившуюся у патогенных организмов, обладают огромной перспективой для разработки новых экологически благоприятных препаратов для защиты растений. В океанах представлено значительное разнообразие организмов (бактерий, микроскопических водорослей, водорослей, позвоночных и беспозвоночных животных), которые являются источником новых биологически активных молекул и до сих пор недостаточно изучены (2). Например, морские микроорганизмы накапливают биологически активные вторичные метаболиты, уникальная структура которых не обнаруживается у наземных организмов. Поэтому такие метаболиты потенциально являются новыми молекулами, представляющими интерес. Описаны некоторые вещества, выделенные из морских организмов, обладающие противогрибковой активностью или естественной защитной активностью веществ, но поиск этих молекул еще недостаточно разработан (3).
Микроскопические водоросли представляют собой одноклеточные организмы, играющие ключевую роль в водных экосистемах. Продуцируя органический материал, они играют важную экологическую роль, поскольку составляют нижнюю часть пищевой цепочки морской среды. Однако их невероятная способность к колонизации всех мировых океанов позволяет предположить, что у них, вероятно, развиты эффективные стратегии борьбы с патогенными организмами, а именно посредством продуцирования природных пестицидов. Например, обильная пролиферация микроскопических водорослей, продуцирующих биотоксины, в прибрежных областях приводит к вредоносному цветению водорослей (HABs, от англ. harmful algal blooms), оказывающему значительное влияние на пищевой каскад.
Среди микроскопических водорослей в умеренных и тропических морских водах присутствуют динофлагелляты, относящиеся к порядку Gymnodiniales и семейству Gymnodiniacae, живущие в свободной форме или в симбиозе с беспозвоночными (например, кораллами). Динофлагелляты синтезируют большое число вторичных метаболитов, называемых поликетидами (соединения с биологической или фармакологической активностью, которые могут быть токсичными для придания преимуществ выживания), несколько из которых были охарактеризованы, в том числе ответственные за HABs (4). Например, модельный вид динофлагеллят Amphidinium carterae продуцирует изобилие различных биологически активных соединений, многие из которых обладают потенциалом для разработки в качестве терапевтических агентов (5). Поликетиды, продуцируемые видом Amphidinium, крайне разнообразны по структуре и составляют три категории: макролиды, линейные поликетиды и длинноцепочечные поликетиды. Например, амфидинолы представляют собой полигидроксиполиены (длинноцепочечные поликетиды), обладающие сильной противогрибковой и гемолитической активностью. Таким образом, они повышают проницаемость мембраны за счет связывания с мембранными липидами (6). Среди различных штаммов Amphidinium выделены амфидинол-подобные соединения с длинной полигидрокси-цепью, такие как лингшуйолы (lingshuiols), каратунгиолы (karatungiols), картераол Е, лютеофанолы, колопсинолы и амфезонол А (5).
В целях ограничения сильной химической нагрузки на окружающую среду и здоровье необходимо выделять молекулы природного происхождения, которые сыграют роль в защите сельскохозяйственных культур от инфекционных заболеваний и в конечном счете заменят химические препараты для защиты растений, используемые в настоящее время. Эти «органические пестициды» могут, таким образом, непосредственно влиять на выживание фитопатогенов сельскохозяйственных культур, имеющих важное агрономическое значение, таких как пшеница и виноградная лоза.
Авторы изобретения неожиданно обнаружили фунгицидное действие клеточного экстракта Amphidinium carterae на многие грибы, патогенные для растений.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Первый объект изобретения относится к применению клеточного экстракта одной или более микроскопических водорослей рода Amphidinium для его фунгицидной и/или бактерицидной активности в отношении патогенных грибов, оомицетов и/или бактерий сельскохозяйственных культур и семян.
Еще один объект изобретения относится к способу получения клеточного экстракта одной или более микроскопической(-их) водоросли(-ей) рода Amphidinium, характеризующемуся следующими стадиями:
- сбор свежих клеток из одной или более микроскопической(-их) водоросли (-ей) рода Amphidinium;
- необязательно замораживание и/или лиофилизация указанных клеток;
- суспендирование указанных свежих или замороженных клеток или лиофилизата в неорганическом или органическом растворителе в массовом соотношении от 1:200 до 1:2;
- необязательно лиофилизация полученного экстракта.
Еще один объект изобретения относится к клеточному экстракту или лиофилизату клеток Amphidinium, которые могут быть получены описанным выше способом.
Еще один объект изобретения относится к способу борьбы с патогенными грибами, оомицетами и/или бактериями сельскохозяйственных культур и семян, включающему нанесение на сельскохозяйственные культуры и/или покрытие семян клеточным экстрактом одной или более микроскопических водорослей рода Amphidinium или экстрактом в соответствии с изобретением.
Еще один объект изобретения относится к способу борьбы с патогенными грибами, оомицетами и/или бактериями сельскохозяйственных культур и семян, включающему следующие стадии:
- перемешивание клеточного экстракта по изобретению в воде в соотношении от 1:4 до 1:800 непосредственно перед применением;
- нанесение этой смеси на сельскохозяйственные культуры и/или покрытие семян указанной смесью.
Еще один объект изобретения относится к способу борьбы с патогенными грибами, оомицетами и/или бактериями сельскохозяйственных культур и семян, включающему следующие стадии:
- ресуспендирование лиофилизата культуры клеток одной или более микроскопических водорослей рода Amphidinium в массовой концентрации, составляющей от 5 до 500 г/л, предпочтительно в массовой концентрации, составляющей от 50 до 400 г/л, предпочтительно в массовой концентрации, составляющей от 100 до 200 г/л, в воде или в органическом или неорганическом растворителе в массовом соотношении от 1:200 до 1:2;
- смешивание в воде в соотношении от 1:4 до 1:800 непосредственно перед применением;
- нанесение полученного экстракта на сельскохозяйственные культуры и/или покрытие семян указанной смесью.
ПОДПИСИ К ФИГУРАМ
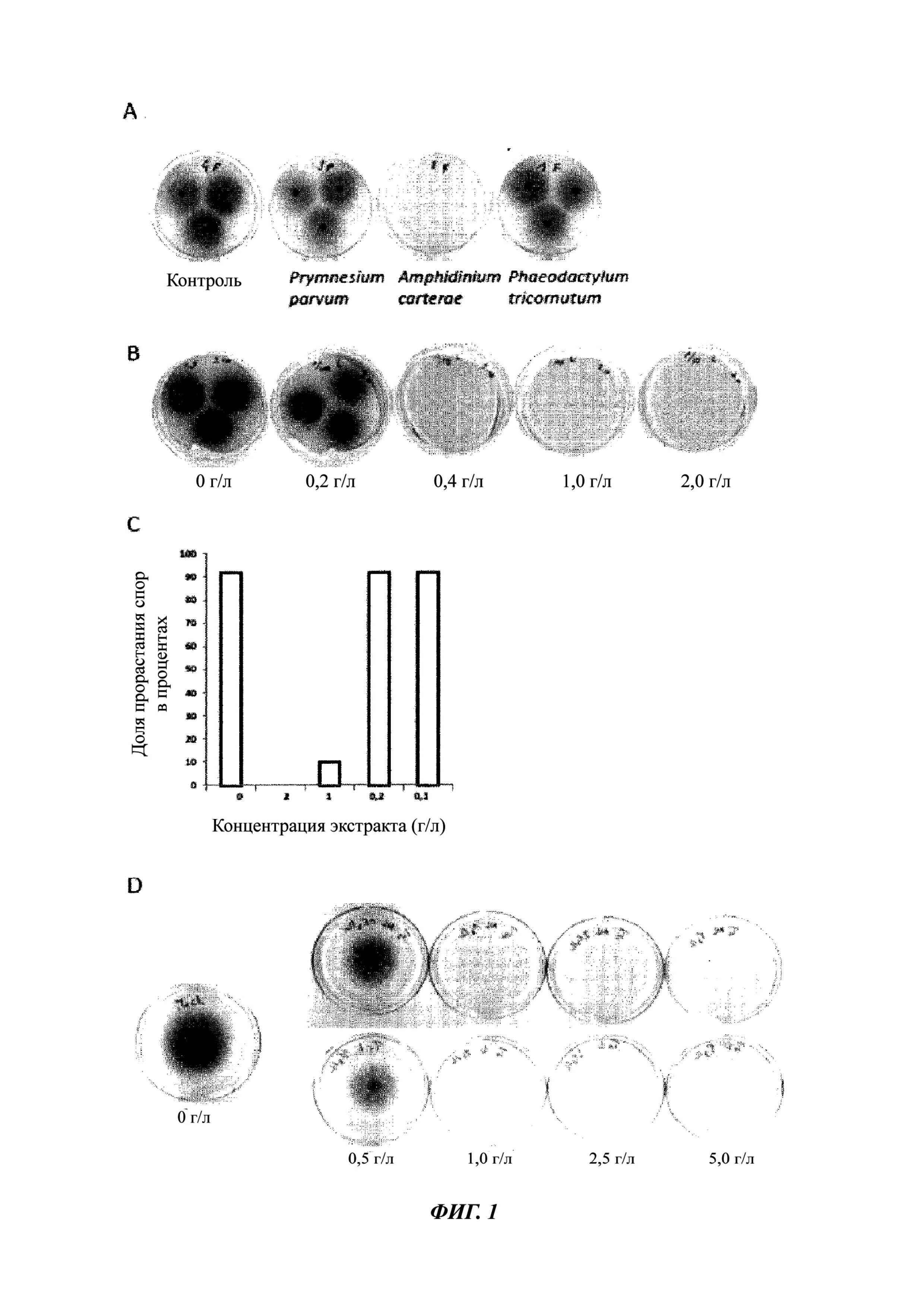
Фиг. 1. Экстракт Amphidinium carterae обладает противогрибковой активностью в отношении Fusarium graminearum.
A. Споры Fusarium graminearum инкубировали в присутствии экстрактов из различных культур микроскопических водорослей, а затем помещали на культуральную среду in vitro. Фотоснимки были получены через 72 часа.
B. Споры Fusarium graminearum инкубировали в присутствии экстрактов из культуры Amphidinium carterae в диапазоне концентраций (от 0 г/л до 2,0 г/л), а затем помещали на культуральную среду in vitro. Фотоснимки были получены через 72 часа.
C. Аналогично В, но количество проросших спор подсчитывали через 6 часов после инкубации.
D. Споры Fusarium graminearum инкубировали в присутствии экстрактов Amphidinium carterae, которые были предварительно заморожены (Фиг. 1D, верхний ряд) или лиофилизированы (Фиг. 1D, нижний ряд).
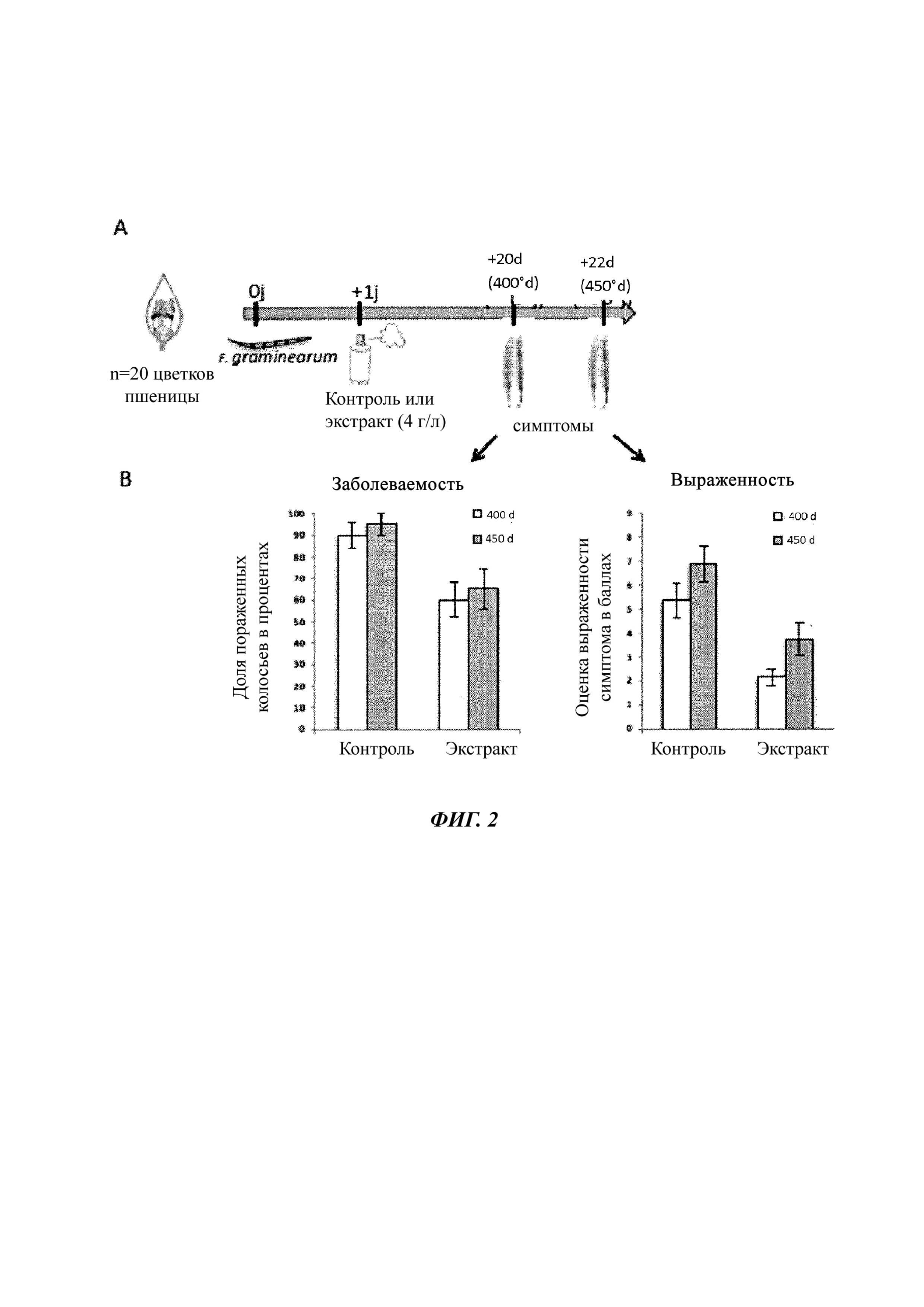
Фиг. 2. Экстракт Amphidinium carterae ингибирует рост патогенных грибов пшеницы in vitro и in planta.
A. Процедура инокуляции цветков пшеницы спорами Fusarium graminearum и их обработка спустя 24 часа раствором в присутствии или в отсутствие (холостая проба) экстракта Amphidinium carterae.
B. Отслеживают появление и развитие симптомов и регистрируют уровни охвата и выраженности (балл от 0 до 9) при 400°D и 450°D.
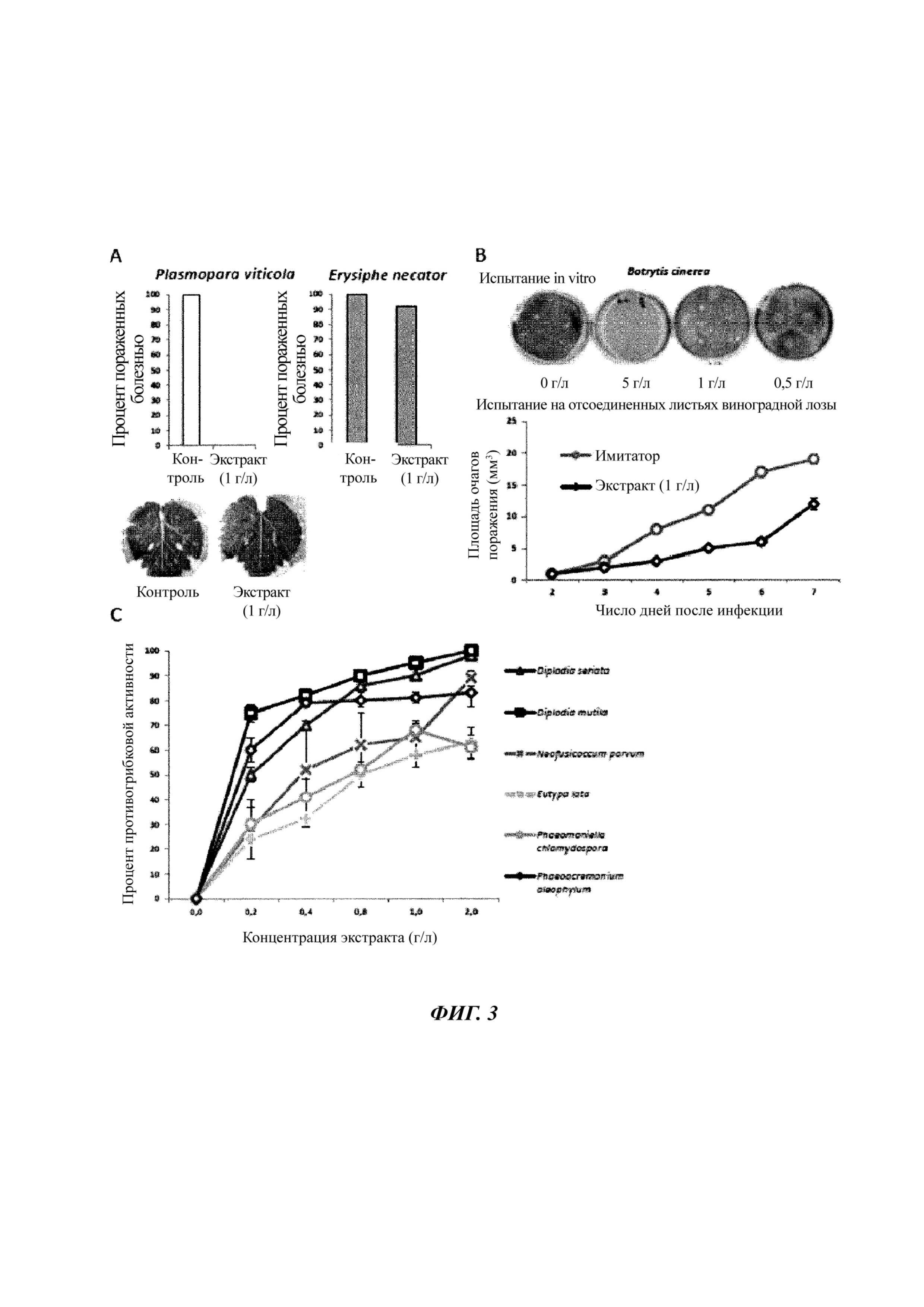
Фиг. 3. Экстракт Amphidinium carterae ингибирует рост патогенных грибов виноградной лозы in vitro и in planta.
A. Отсоединенные листья виноградной лозы, поддерживаемые в условиях in vitro, опрыскивали экстрактом Amphidinium carterae (1 г/л) или стерильной водой (имитатор), а затем высевали на эти листья спорангии Plasmopara viticola или конидии Erysiphe necator. Симптомы регистрировали через 7 дней и через 12 дней соответственно.
B. Аналогично А, но на обработанные листья высевали мицелиальные импланты Botrytis cinerea. Симптомы регистрировали через 7 дней путем измерения размера некротических очагов поражения.
C. Мицелиальные имплантаты различных грибов, вовлеченных в болезнь эска виноградной лозы, высевали на культуральную среду и спустя 24 часа обрабатывали водой (имитатор) или экстрактом A. carterae в различных концентрациях. Симптомы регистрируют путем измерения площади поверхности мицелия через 4 дня.
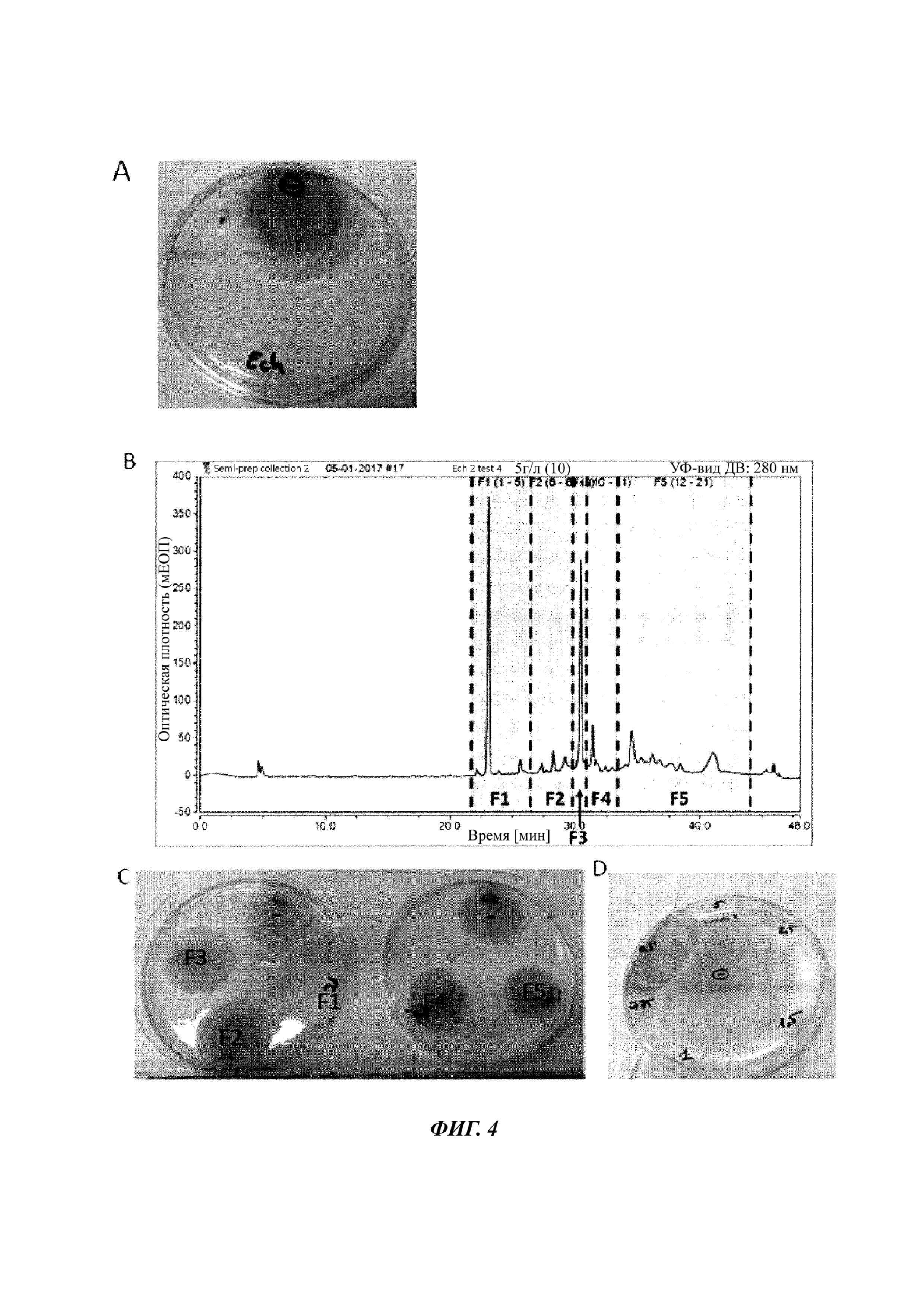
Фиг. 4. Фракционирование экстракта D на колонке С18 с обращенной фазой и испытание на активность различных фракций экстракта D, полученных методом ВЭЖХ.
A. Испытание на ингибирование роста F. graminearum in vitro экстрактом D в концентрации 5 г/л, полученным после экстракции метанолом.
B. Жидкостная хроматограмма, полученная после введения экстракта D. Образцы были собраны в соответствии с пунктирными линиями с образованием 5 фракций, обозначенных F1-F5.
C. Испытание на ингибирование роста F. graminearum in vitro фракциями F1-F5 в концентрации 5 г/л.
D. Испытание на ингибирование роста F. graminearum in vitro фракцией F1 в концентрации 0,5 г/л, 0,75 г/л, 1 г/л, 1,5 г/л, 2,5 г/л и 5 г/л.
-: отрицательный контроль: споры инкубировали в присутствии только буферного раствора.
Фиг. 5. Фракционирование фракции F1 на колонке С18 с обращенной фазой и испытание на активность различных фракций, полученных методом ВЭЖХ
A. Жидкостная хроматограмма, полученная после введения фракции F1. Образцы были собраны в соответствии с пунктирными линиями с образованием 5 фракций, обозначенных F1-1-F1-5.
B. Испытание на ингибирование роста F. graminearum in vitro фракциями F1-F5 в концентрации 5 г/л.
C. Испытание на ингибирование роста F. graminearum in vitro фракцией F1-2 в концентрациях в диапазоне от 0,0005 г/л до 1 г/л.
-: отрицательный контроль: споры инкубировали в присутствии только буферного раствора.
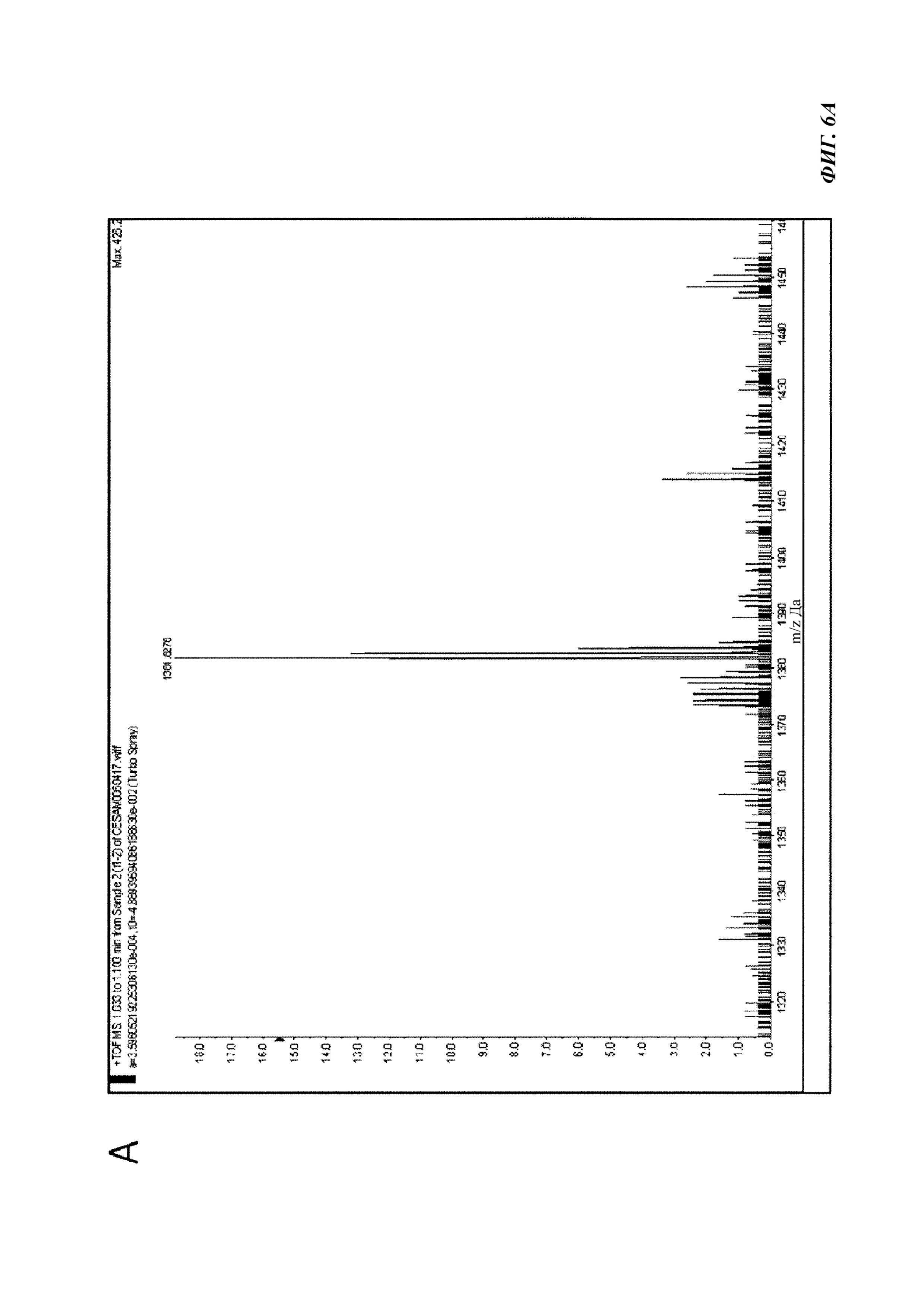
Фиг. 6. Анализ фракции F1-2 методом масс-спектрометрии.
A. Масс-спектр, полученный при ионизации электрораспылением в режиме регистрации положительного иона для интересующей молекулы из собранной фракции F1-2.
B. Тандемный масс-спектр иона 1381,8276 Да, детектированного в А. Круговые пики соответствуют обнаруживаемым у соединения AM 18, охарактеризованного Nuzzo et al., 2014 (7).
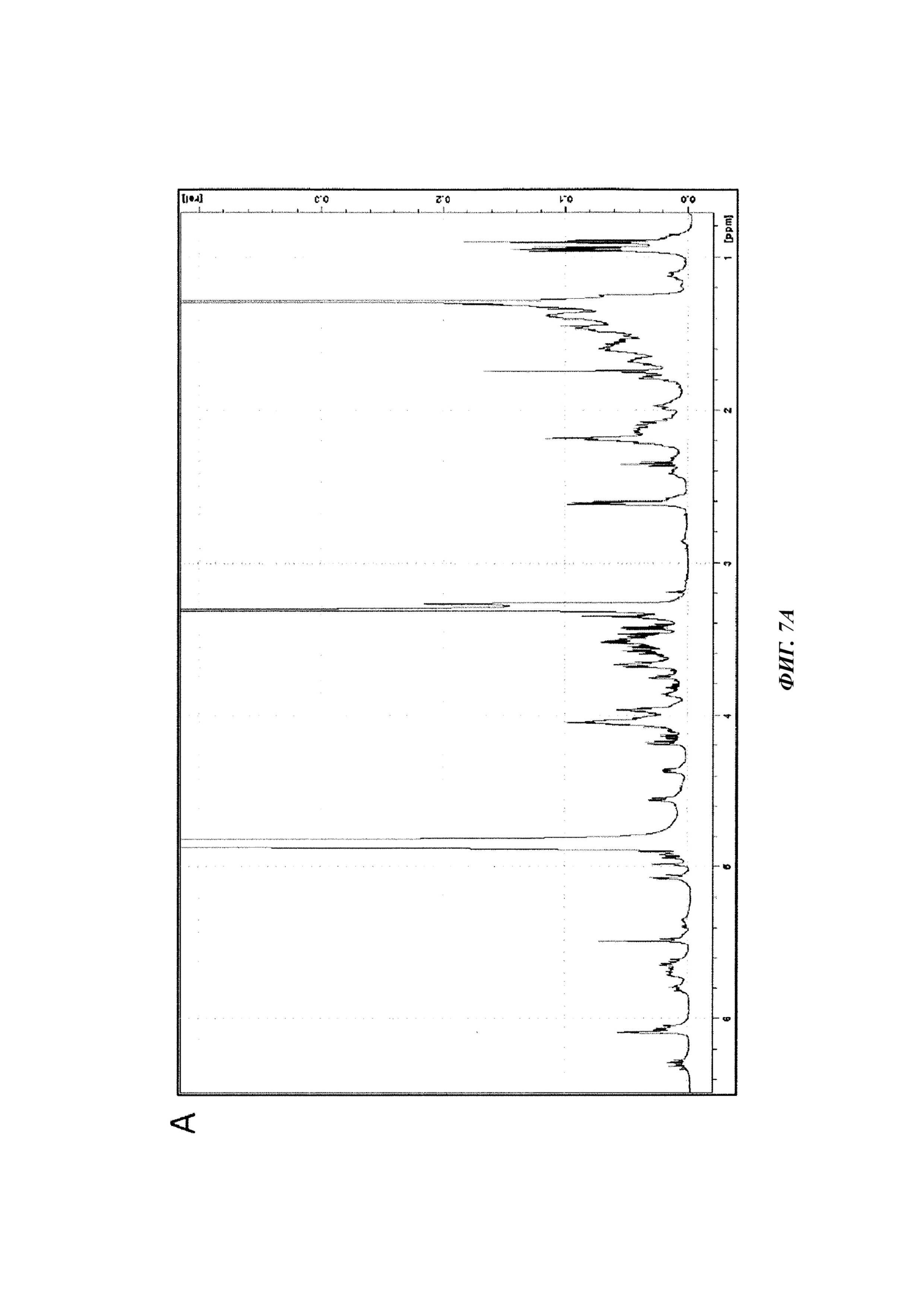
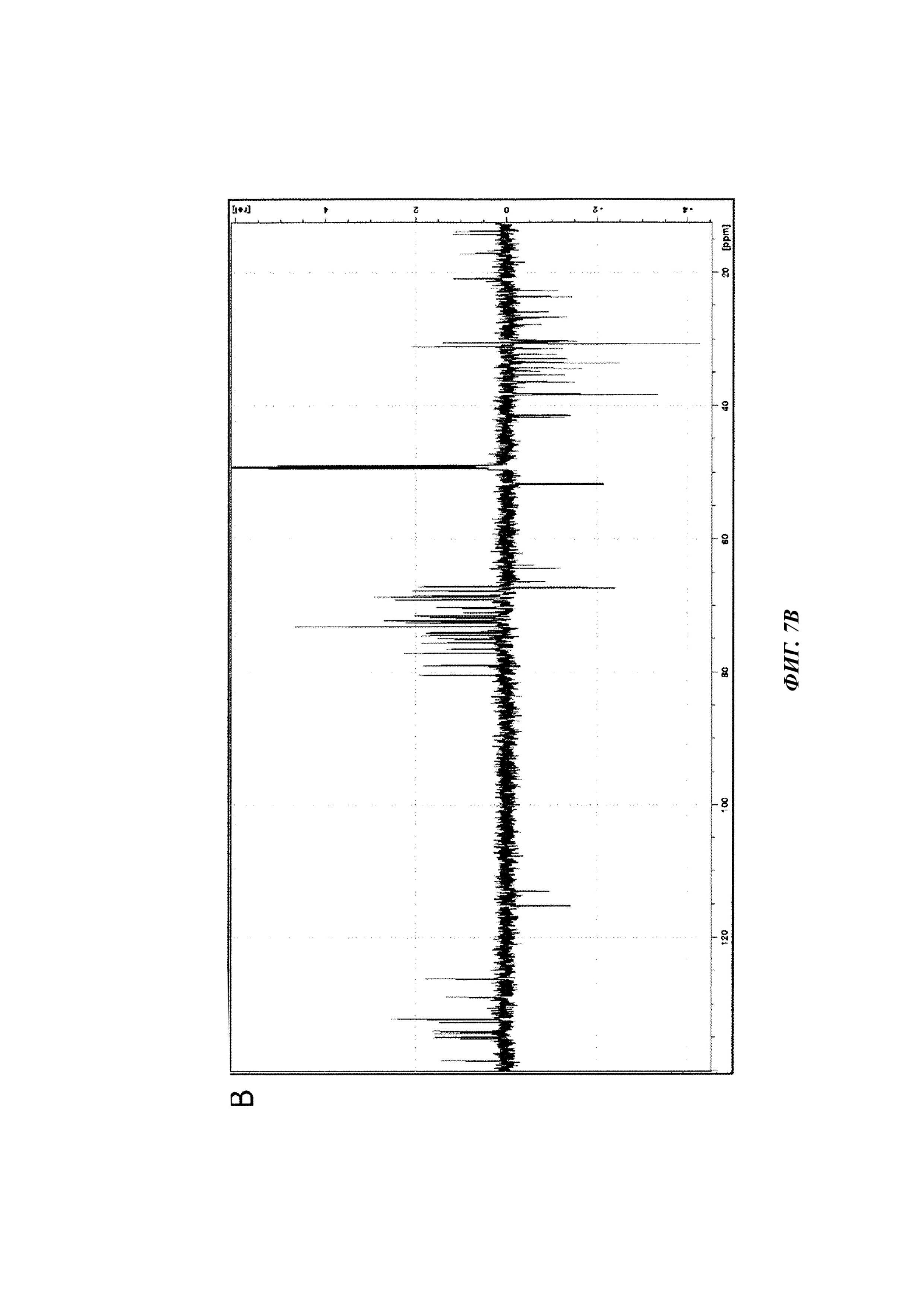
Фиг. 7. Анализ фракции F1-2 методом ЯМР.
A. Спектр1Н ЯМР (растворитель MeOD4), полученный для молекулы фракции F1-2, выделенной из экстракта D.
B. Спектр13С DEPT135 ЯМР (растворитель MeOD4), полученный для молекулы фракции F1-2, выделенной из экстракта D.
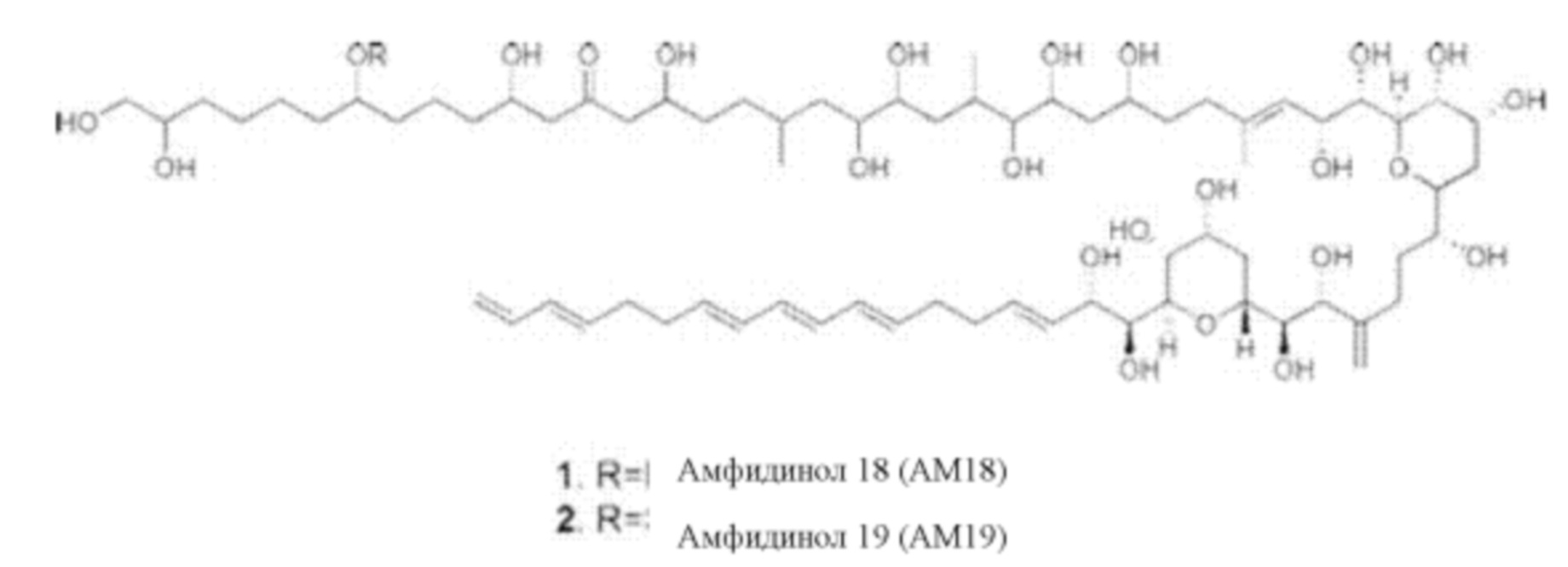
Фиг. 8. Структурная формула молекулы амфидинола 18, выделенной из фракции F1-2 экстракта D
Все корреляции COSY (англ. аббревиатура от "correlation spectroscopy" - корреляционная спектроскопия) и TOCSY (англ. аббревиатура от "Total Correlation Spectroscopy" - полная корреляционная спектроскопия) между каждым из протонов показаны полужирным шрифтом.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Первый объект изобретения относится к применению клеточного экстракта одной или более микроскопических водорослей рода Amphidinium для его фунгицидной и/или бактерицидной активности в отношении патогенных грибов, оомицетов и/или бактерий сельскохозяйственных культур и семян.
Вид Amphidinium
Подходящий вид Amphidinium выбирают из группы, состоящей из Amphidinium achromaticum, Amphidinium aculeatum, Amphidinium acutissimum, Amphidinium acutum, Amphidinium alinii, Amphidinium aloxalocium, Amphidinium amphidinioides, Amphidinium asymmetricum, Amphidinium aureum, Amphidinium belauense, Amphidinium bidentatum, Amphidinium bipes, Amphidinium boekhoutensis, Amphidinium boggayum, Amphidinium caerulescens, Amphidinium carbunculus, Amphidinium carterae, Amphidinium celestinum, Amphidinium chattonii, Amphidinium coeruleum, Amphidinium conradii, Amphidinium conus, Amphidinium coprosum, Amphidinium corallinum, Amphidinium corpulentum, Amphidinium crassum, Amphidinium cristatum, Amphidinium cucurbita, Amphidinium cucurbitella, Amphidinium cupulatisquama, Amphidinium curvatum, Amphidinium cyaneoturbo, Amphidinium dentatum, Amphidinium discoidale, Amphidinium dubium, Amphidinium eilatiensis, Amphidinium emarginatum, Amphidinium fastigium, Amphidinium filum

Предпочтительно микроскопической водорослью или одной из них рода Amphidinium, используемой в соответствии с изобретением, является Amphidinium carterae. В коллекции имеется несколько штаммов Amphidinium carterae, такие как штаммы ССМР 124, 1314, 3177 (ССМР (от англ. Culture Collection of Marine Phytoplankton) = коллекция культур морского фитопланктона), АС 208, 792 (АС = Algobank Cean), BEA 01198 (BEA = Banco Espanol de Algas).
Преимущественно используемым в соответствии с изобретением штаммом Amphidinium carterae является ССМР 1314, АС208 или АС792.
Экстракция
Указанный экстракт может быть получен любым способом экстракции из клеток, известным специалисту в данной области техники - экстракцией в системе твердое вещество-жидкость или жидкость-жидкость, например экстракцией в неорганический или органический растворитель, который может быть выбран из группы, состоящей из воды, водных растворов, кетонов, сложных эфиров, кислот, простых эфиров, спиртов и смесей этих растворителей в любом смешиваемом соотношении.
Предпочтительным растворителем преимущественно будет вода или окисленные растворители, предпочтительно спирты, особенно предпочтительно С1-С4 спирты, такие как метанол или этанол.
Особенно предпочтительны С1-С4 спирты.
Предпочтительно указанный экстракт представляет собой водорастворимую фракцию.
Предпочтительно экстракт содержит амфидинол 18 или амфидинол 19, особенно предпочтительно амфидинол 18, преимущественно в количестве, составляющем более 1% масс./масс. в расчете на общую массу экстракта, предпочтительно составляющем от 2 до 10% масс./масс. в расчете на общую массу экстракта, особенно предпочтительно составляющем от 3 до 5% масс./масс. в расчете на общую массу экстракта.
Механизм действия
Указанная фунгицидная активность в отношении патогенных грибов и/или оомицетов сельскохозяйственных культур и семян может, в частности, проявляться посредством ингибирования прорастания спор или посредством ингибирования роста грибов и/или оомицетов.
Данная активность осуществляется посредством литической активности в отношении клеточной стенки и мембраны, приводящей к лизису клетки.
Сельскохозяйственные культуры
Сельскохозяйственные культуры, в частности, выбраны из группы, состоящей из зерновых, таких как пшеница, кукуруза, ячмень, рис, соя; фруктов и овощей, таких как картофель, морковь, яблони, персиковые деревья, абрикосовые деревья, томаты, редис, фасоль; виноградной лозы и декоративных растений.
Указанные сельскохозяйственные культуры, в частности, выбраны из группы, состоящей из родов Abelmoschus, Acacia, Achras, Agave, Agrostis, Aleurites, Allium, Anacardium, Ananas, Annona, Apium, Arachis, Areca, Armoracia, Arracacia, Artocarpus, Asparagus, Aspidosperma, Avena, Bertholletia, Beta, Boehmeria, Borassus, Brassica, Cajanus, Camellia, Cannabis, Capsicum, Carica, Carthamus, Carum, Carya, Castanea, Ceiba, Ceratonia, Chenopodium, Chrysanthemum, Cicer, Cichorium, Cinchona, Cinnamomum, Citrullus, Citrus, Cocos, Coffea, Cola, Colocasia, Corchorus, Corylus, Crotalaria, Cucumis, Cucurbita, Cydonia, Cymbopogon, Cynara, Dactylis, Daucus, Dioscorea, Diospyros, Echinochloa, Elaeis, Elettaria, Eleusine, Eragrostis, Eriobotrya, Eugenia, Fagopyrum, Ficus, Foeniculum, Fragaria, Furcraea, Glycine, Glycyrrhiza, Gossypium, Guizotia, Helianthus, Hevea, Hibiscus, Hordeum, Humulus, Ilex, Indigofera, Ipomoea, Jasminum, Juglans, Lactuca, Lagenaria, Lavandula, Lawsonia, Lens, Lepidium, Lespedeza, Linum, Litchi, Lolium, Lopmoea, Lotus, Lupinus, Lycopersicon, Lygeum, Macadamia, Malus, Mangifera, Manihot, Maranta, Medicago, Mentha, Mespilus, Metroxylon, Moringa, Musa, Myristica, Nicotiana, Olea, Onobrychis, Oryza, Panicum, Papaver, Pastinaca, Pelargonium, Pennisetum, Persea, Phaseolus, Phleum, Phoenix, Phormium, Pimpinella, Piper, Pistacia, Pisum, Prunus, Psidium, Punica, Pyrus, Raphanus Rheum, Ribes, Ricinus, Rose, Rubus, Saccharum, Scorzonera, Secale Sechium, Sesamum, Setaria, Solanum, Sorghum, Spinacia, Theobroma, Tragopogon, Trifolium, Trigonella, Triticum, Urena, Vaccinium, Valerianella, Vanilla, Vicia, Vigna, Vitellaria, Vitis, Xanthosoma, Zea, Zingiber.
Патогенные организмы
Патогенные грибы указанных сельскохозяйственных культур и семян являются аскомицетами или базидиомицетами, предпочтительно аскомицетами.
Патогенными грибами указанных сельскохозяйственных культур и семян являются грибы, патогенные для сельскохозяйственных культур и семян родов:
Acrocalymma, Acrocalymma medicaginis,
Fusarium, Fusarium affine, Fusarium arthrosporioides, Fusarium crookwellense, Fusarium culmorum, Fusarium graminearum, Fusarium moniliforme, Fusarium incarnatum, Fusarium solani, Fusarium langsethiae, Fusarium mangiferae, Fusarium oxysporum f. sp. albedinis, Fusarium oxysporum f. sp. asparagi, Fusarium oxysporum f. sp. batatas, Fusarium oxysporum f. sp. betae, Fusarium oxysporum f. sp. cannabis, Fusarium oxysporum f. sp. carthami, Fusarium oxysporum f. sp. cattleyae, Fusarium oxysporum f. sp. ciceris, Fusarium oxysporum f. sp. coffea, Fusarium oxysporum f. sp. cubense, Fusarium oxysporum f. sp. cyclaminis, Fusarium oxysporum f. sp. dianthi, Fusarium oxysporum f. sp. lentis, Fusarium oxysporum f. sp. lini, Fusarium oxysporum f. sp. lycopersici, Fusarium oxysporum f. sp. medicaginis, Fusarium oxysporum f. sp. pisi, Fusarium oxysporum f. sp. radicis-lycopersici, Fusarium oxysporum f. sp. spinacia, Fusarium oxysporum, Fusarium pallidoroseum, Fusarium patch, Fusarium proliferatum, Fusarium redolens, Fusarium sacchari, Fusarium solani, Fusarium subglutinans, Fusarium sulphureum, Fusarium tricinctum, Fusarium wilt,
Botrytis, Botrytis allii, Botrytis anthophila, Botrytis cinerea, Botrytis fabae, Botrytis narcissicola,
Alternaria, Alternaria alternata, Alternaria brassicae, Alternaria brassicicola, Alternaria carthami, Alternaria cinerariae, Alternaria dauci, Alternaria dianthi, Alternaria dianthicola, Alternaria euphorbiicola, Alternaria helianthi, Alternaria helianthicola, Alternaria japonica, Alternaria leucanthemi, Alternaria limicola, Alternaria linicola, Alternaria padwickii, Alternaria panax, Alternaria radicina, Alternaria raphani, Alternaria saponariae, Alternaria senecionis, Alternaria solani, Alternaria tenuissima, Alternaria triticina, Alternaria zinniae,
Erisyphe, Erisyphe necator, Erysiphe betae, Erysiphe brunneopunctata, Erysiphe cichoracearum, Erysiphe cruciferarum, Erysiphe graminis f. sp. Avenae, Erysiphe graminis f. sp. tritici, Erysiphe heraclei, Erysiphe pisi,
Claviceps, Claviceps fusiformis, Claviceps purpurea, Claviceps sorghi, Claviceps zizaniae,
Gaeumannomyces, Gaeumannomyces graminis,
Leptosphaeria, Leptosphaeria nodorum, Leptosphaeria acuta, Leptosphaeria cannabina, Leptosphaeria coniothyrium, Leptosphaeria libanotis, Leptosphaeria lindquistii, Leptosphaeria maculans, Leptosphaeria musarum, Leptosphaeria pratensis, Leptosphaeria sacchari, Leptosphaeria woroninii,
Microdochium, Microdochium spp. Microdochium bolleyi, Microdochium dimerum, Microdochium panattonianum, Microdochium phragmitis,
Mycosphaerella, Mycosphaerella arachidis, Mycosphaerella areola, Mycosphaerella berkeleyi, Mycosphaerella bolleana, Mycosphaerella brassicicola, Mycosphaerella caricae, Mycosphaerella caryigena, Mycosphaerella cerasella, Mycosphaerella coffeicola, Mycosphaerella confusa, Mycosphaerella cruenta, Mycosphaerella dendroides, Mycosphaerella eumusae, Mycosphaerella gossypina, Mycosphaerella graminicola, Mycosphaerella henningsii, Mycosphaerella horii, Mycosphaerella juglandis, Mycosphaerella lageniformis, Mycosphaerella linicola, Mycosphaerella louisianae, Mycosphaerella musae, Mycosphaerella musicola, Mycosphaerella palmicola, Mycosphaerella pinodes, Mycosphaerella pistaciarum, Mycosphaerella pistacina, Mycosphaerella platanifolia, Mycosphaerella polymorpha, Mycosphaerella pomi, Mycosphaerella punctiformis, Mycosphaerella pyri,
Oculimacula, Oculimacula acuformis, Oculimacula yallundae,
Blumeria, Blumeria graminis,
Pyrenophora, Pyrenophora avenae, Pyrenophora chaetomioides, Pyrenophora graminea, Pyrenophora seminiperda, Pyrenophora teres, Pyrenophora teres f. maculata, Pyrenophora teres f. teres, Pyrenophora tritici-repentis,
Ramularia, Ramularia colic-cygni, Ramularia beticola, Ramularia coryli, Ramularia cyclaminicola, Ramularia macrospora, Ramularia menthicola, Ramularia necator, Ramularia primulae, Ramularia spinaciae, Ramularia subtilis, Ramularia tenella, Ramularia vallisumbrosae,
Rhynchosporium, Rhynchosporium secalis,
Cochliobolus, Cochliobolus, Cochliobolus carbonum, Cochliobolus cymbopogonis, Cochliobolus hawaiiensis, Cochliobolus heterostrophus, Cochliobolus lunatus, Cochliobolus miyabeanus, Cochliobolus ravenelii, Cochliobolus sativus, Cochliobolus setariae, Cochliobolus spicifer, Cochliobolus stenospilus, Cochliobolus tuberculatus, Cochliobolus victoriae,
Microdochium, Microdochium oryzae,
Pyricularia, Pyricularia oryzae,
Sarocladium, Sarocladium oryzae,
Ustilaginoides, Ustilaginoides virens,
Cercospora, Cercospora, Cercospora apii, Cercospora apii f. sp. clerodendri, Cercospora apiicola, Cercospora arachidicola, Cercospora asparagi, Cercospora atrofiliformis, Cercospora beticola, Cercospora brachypus, Cercospora brassicicola, Cercospora brunkii, Cercospora cannabis, Cercospora cantuariensis, Cercospora capsici, Cercospora carotae, Cercospora corylina, Cercospora fuchsiae, Cercospora fusca, Cercospora fusimaculans, Cercospora gerberae, Cercospora halstedii, Cercospora handelii, Cercospora hayi, Cercospora hydrangeae, Cercospora kikuchii, Cercospora lentis, Cercospora liquidambaris, Cercospora longipes, Cercospora longissima, Cercospora mamaonis, Cercospora mangiferae, Cercospora medicaginis, Cercospora melongenae, Cercospora minuta, Cercospora nicotianae, Cercospora odontoglossi, Cercospora papayae, Cercospora penniseti, Cercospora pisa-sativae, Cercospora platanicola, Cercospora puderii, Cercospora pulcherrima, Cercospora rhapidicola, Cercospora rosicola, Cercospora sojina, Cercospora solani, Cercospora solani-tuberosi, Cercospora sorghi, Cercospora theae, Cercospora tuberculans, Cercospora vexans, Cercospora vicosae, Cercospora zeae-maydis, Cercospora zebrina, Cercospora zonata,
Corynespora, Corynespora cassiicola,
Phakospora, Phakospora pachyrhizi, Phakopsora gossypii, Colletotrichum, Colletotrichum acutatum, Colletotrichum arachidis, Colletotrichum capsici, Colletotrichum cereale, Colletotrichum coffeanum, Colletotrichum crassipes, Colletotrichum dematium, Colletotrichum dematium f. spinaciae, Colletotrichum derridis, Colletotrichum destructivum, Colletotrichum gloeosporioides, Colletotrichum glycines, Colletotrichum gossypii, Colletotrichum graminicola, Colletotrichum higginsianum, Colletotrichum kahawae, Colletotrichum lindemuthianum, Colletotrichum lini, Colletotrichum mangenotii, Colletotrichum musae, Colletotrichum nigrum, Colletotrichum orbiculare, Colletotrichum pisi, Colletotrichum sublineolum, Colletotrichum trichellum, Colletotrichum trifolii, Colletotrichum truncatum,
Pythium spp.,
Diplodia, Diplodia allocellula, Diplodia laelio-cattleyae, Diplodia manihoti, Diplodia paraphysaria, Diplodia seriata, Diplodia theae-sinensis,
Monilia, Monilinia azaleae, Monilinia fructicola, Monilinia fructigena, Monilinia laxa, Monilinia oxycocci,
Pezzicula, Pezzicula alba, Pezzicula malicorticis,
Zymoseptoria, Zymoseptoria tritici
Phytophthora, Phytophthora infestans
Guignardia, Guignardia bidwelli, Guignardia camelliae, Guignardia fulvida, Guignardia mangiferae, Guignardia musae, Guignardia philoprina, Plasmopara, Plasmopara viticola,
Puccinia, Puccinia angustata, Puccinia arachidis, Puccinia aristidae, Puccinia asparagi, Puccinia cacabata, Puccinia campanulae, Puccinia carthami, Puccinia coronata, Puccinia dioicae, Puccinia erianthi, Puccinia extensicola, Puccinia helianthi, Puccinia hordei, Puccinia jaceae, Puccinia kuehnii, Puccinia malvacearum, Puccinia mariae-wilsoniae, Puccinia melanocephala, Puccinia menthae, Puccinia oxalidis, Puccinia pelargonii-zonalis, Puccinia pittieriana, Puccinia poarum, Puccinia purpurea, Puccinia recondita, Puccinia schedonnardii, Puccinia sessilis, Puccinia striiformis, Puccinia striiformis, Puccinia subnitens, Puccinia substriata, Puccinia verruca, Puccinia xanthii,
Rhizoctonia, Rhizoctonia solani, Rhizoctonia oryzae, Rhizoctonia cerealis, Rhizoctonia leguminicola, Rhizoctonia rubi,
Sclerotinia, Sclerotinia borealis, Sclerotinia bulborum, Sclerotinia minor, Sclerotinia ricini, Sclerotinia sclerotiorum, Sclerotinia spermophila, Sclerotinia trifoliorum,
Septoria, Septoria ampelina, Septoria azaleae, Septoria bataticola, Septoria campanulae, Septoria cannabis, Septoria cucurbitacearum, Septoria darrowii, Septoria dianthi, Septoria eumusae, Septoria glycines, Septoria helianthi, Septoria humuli, Septoria hydrangeae, Septoria lactucae, Septoria lycopersici, Septoria lycopersici, Septoria menthae, Septoria passerinii, Septoria pisi, Septoria rhododendri, Septoria secalis, Septoria selenophomoides,
Venturia, Venturia inaequalis. Venturia carpophila,
Acrodontium, Acrodontium simplex,
Acrophialophora, Acrophialophora fusispora,
Acrosporium, Acrosporium tingitaninum,
Aecidium, Aecidium aechmantherae, Aecidium amaryllidis, Aecidium breyniae, Aecidium campanulastri, Aecidium cannabis, Aecidium cantensis, Aecidium caspicum, Aecidium foeniculi, Aecidium narcissi,
Ahmadiago,
Albonectria, Albonectria rigidiuscula,
Allodus, Modus podophylli,
Amphobotrys, Amphobotrys ricini,
Anguillosporella, Anguillosporella vermiformis,
Anthostomella, Anthostomella pullulans,
Antrodia, Antrodia albida, Antrodia serialiformis, Antrodia serialis,
Apiospora, Apiospora montagnei,
Appendiculella,
Armillaria, Armillaria heimii, Armillaria sinapina, Armillaria socialis, Armillaria tabescens,
Arthrocladiella,
Arthuriomyces, Arthuriomyces peckianus,
Ascochyta, Ascochyta asparagina, Ascochyta bohemica, Ascochyta caricae, Ascochyta doronici, Ascochyta fabae f. sp. lentis, Ascochyta graminea, Ascochyta hordei, Ascochyta humuli, Ascochyta pisi, Ascochyta prasadii, Ascochyta sorghi, Ascochyta spinaciae, Ascochyta tarda, Ascochyta tritici,
Ascospora, Ascospora ruborum,
Aspergillus, Aspergillus aculeatus, Aspergillus fischerianus, Aspergillus niger,
Asperisporium, Asperisporium caricae,
Asteridiella,
Asteroma, Asteroma caryae,
Athelia, Athelia arachnoidea, Athelia rolfsii,
Aurantiporus, Aurantiporus fissilis,
Aureobasidium, Aureobasidium pullulans,
Bambusiomyces,
Пятнистость банана,
Фузариозное увядание финиковой пальмы,
Beniowskia, Beniowskia sphaeroidea,
Bionectria, Bionectria ochroleuca,
Bipolaris, Bipolaris cactivora, Bipolaris cookei, Bipolaris incurvata, Bipolaris sacchari,
Biscogniauxia, Biscogniauxia capnodes, Biscogniauxia marginata,
Bjerkandera, Bjerkandera adusta,
Черная сигатока,
Blakeslea, Blakeslea trispora,
Botryodiplodia, Botryodiplodia oncidii, Botryodiplodia ulmicola,
Botryosphaeria, Botryosphaeria cocogena, Botryosphaeria dothidea, Botryosphaeria marconii, Botryosphaeria obtusa, Botryosphaeria rhodina, Botryosphaeria ribis, Botryosphaeria stevensii,
Botryosporium, Botryosporium pulchrum,
Botryotinia, Botryotinia fuckeliana, Botryotinia polyblastis,
Болезнь самшита,
Brachybasidiaceae,
Brasiliomyces, Brasiliomyces malachrae,
Briosia, Briosia ampelophaga,
Бурая кольцевая пятнистость,
Фитофтороз томатов,
Bulbomicrosphaera,
Cadophora, Cadophora malorum,
Caespitotheca,
Calonectria, Calonectria ilicicola, Calonectria indusiata, Calonectria kyotensis, Calonectria pyrochroa, Calonectria quinqueseptata,
Camarotella, Camarotella acrocomiae, Camarotella costaricensis,
Ржавчина канны,
Capitorostrum, Capitorostrum cocoes,
Capnodium, Capnodium footii, Capnodium mangiferum, Capnodium ramosum, Capnodium theae,
Cephalosporium, Cephalosporium gramineum,
Ceratobasidium, Ceratobasidium cereale, Ceratobasidium cornigerum, Ceratobasidium noxium, Ceratobasidium ramicola, Ceratobasidium setariae, Ceratobasidium stevensii,
Ceratocystis, Ceratocystis adiposa, Ceratocystis coerulescens, Ceratocystis fimbriata, Ceratocystis moniliformis, Ceratocystis oblonga, Ceratocystis obpyriformis, Ceratocystis paradoxa, Ceratocystis pilifera, Ceratocystis pluriannulata, Ceratocystis polyconidia, Ceratocystis tanganyicensis, Ceratocystis zombamontana,
Ceratorhiza, Ceratorhiza hydrophila,
Ceratospermopsis,
Cercoseptoria, Cercoseptoria ocellata,
Cercosporella, Cercosporella rubi,
Ceriporia, Ceriporia spissa, Ceriporia xylostromatoides,
Cerrena, Cerrena unicolor,
Ceuthospora, Ceuthospora lauri,
Choanephora, Choanephora cucurbitarum, Choanephora infundibulifera,
Chrysanthemum, белая ржавчина хризантемы,
Chrysomyxa, Chrysomyxa cassandrae,
Chrysomyxa, Chrysomyxa himalensis, Chrysomyxa ledi, Chrysomyxa ledi var. rhododendri, Chrysomyxa ledicola, Chrysomyxa nagodhii, Chrysomyxa neoglandulosi, Chrysomyxa piperiana, Chrysomyxa pirolata, Chrysomyxa pyrolae, Chrysomyxa reticulata, Chrysomyxa roanensis, Chrysomyxa succinea,
Cladosporium, Cladosporium arthropodii, Cladosporium cladosporioides, Cladosporium cladosporioides f sp. pisicola, Cladosporium cucumerinum, Cladosporium herbarum, Cladosporium musae, Cladosporium oncobae,
Climacodon, Climacodon pulcherrimus, Climacodon septentrionalis,
Clitocybe, Clitocybe parasitica,
Clonostachys rosea f. rosea,
Clypeoporthe, Clypeoporthe iliau,
Coleosporium, Coleosporium helianthi, Coleosporium ipomoeae, Coleosporium madiae, Coleosporium pacificum, Coleosporium tussilaginis,
Conidiosporomyces,
Coniella, Coniella castaneicola, Coniella diplodiella, Coniella fragariae,
Coniothecium, Coniothecium chomatosporum,
Coniothyrium, Coniothyrium celtidis-australis, Coniothyrium henriquesii, Coniothyrium rosarum, Coniothyrium wernsdorffiae,
Coprinopsis, Coprinopsis psychromorbida,
Cordana, Cordana johnstonii, Cordana musae, Coriolopsis floccosa,
Серая пятнистость листьев кукурузы,
Corticium, Corticium invisum, Corticium penicillatum, Corticium theae,
Coryneopsis, Coryneopsis rubi,
Coryneum, Coryneum rhododendri,
Твердая головня зерна,
Crinipellis, Crinipellis sarmentosa,
Cronartium, Cronartium hbicola,
Cryphonectriaceae,
Cryptobasidiaceae,
Cryptocline, Cryptocline cyclaminis,
Cryptomeliola,
Cryptosporella, Cryptosporella umbrina,
Cryptosporiopsis, Cryptosporiopsis tarraconensis,
Cryptosporium, Cryptosporium minimum,
Curvularia, Curvularia lunata, Curvularia caricae-papayae, Curvularia penniseti, Curvularia senegalensis, Curvularia trifolii,
Cyclaneusma; пожелтение хвои сосны,
Cylindrocarpon, Cylindrocarpon ianthothele var. ianthothele, Cylindrocarpon magnusianum, Cylindrocarpon musae,
Cylindrocladiella, Cylindrocladiella camelliae, Cylindrocladiella parva,
Cylindrocladium, Cylindrocladium clavatum, Cylindrocladium lanceolatum, Cylindrocladium peruvianum, Cylindrocladium pteridis,
Cylindrosporium, Cylindrosporium cannabinum, Cylindrosporium juglandis, Cylindrosporium rubi,
Cymadothea, Cymadothea trifolii,
Cytospora, Cytospora palmarum, Cytospora personata, Cytospora sacchari, Cytospora sacculus, Cytospora terebinthi,
Cytosporina, Cytosporina ludibunda,
Dactuliophora, Dactuliophora elongata,
Davidiella, Davidiella dianthi, Davidiella tassiana,
Deightoniella, Deightoniella papuana, Deightoniella torulosa,
Dendrophora, Dendrophora marconii, Dendrophora erumpens,
Denticularia, Denticularia mangiferae,
Dermea pseudotsugae,
Diaporthaceae,
Diaporthe, Diaporthe arctii, Diaporthe dulcamarae, Diaporthe eres, Diaporthe helianthi, Diaporthe lagunensis, Diaporthe lokoyae, Diaporthe melonis, Diaporthe orthoceras, Diaporthe perniciosa, Diaporthe phaseolorum, Diaporthe phaseolorum var caulivora, Diaporthe phaseolorum var. phaseolorum, Diaporthe phaseolorum var. soja, Diaporthe rudis, Diaporthe tanakae, Diaporthe toxica,
Dicarpella, Dicarpella dryina,
Didymella, Didymella applanata, Didymella bryoniae, Didymella fabae, Didymella lycopersici,
Didymosphaeria, Didymosphaeria arachidicola, Didymosphaeria taiwanensis,
Dilophospora, Dilophospora alopecuri,
Dimeriella, Dimeriella sacchari,
Diplocarpon, Diplocarpon mespili, Diplocarpon rosae,
Discosia, Discosia artocreas,
Discostroma, Discostroma corticola,
Distocercospora, Distocercospora livistonae,
Dothiorella, Dothiorella brevicollis, Dothiorella dominicana, Dothiorella dulcispinae, Dothiorella gregaria,
Drechslera, Drechslera avenacea, Drechslera campanulata, Drechslera dematioidea, Drechslera gigantea, Drechslera glycines, Drechslera musae-sapientium, Drechslera teres f. maculata, Drechslera wirreganensis,
Eballistra, Eballistra lineata, Eballistra oryzae,
Eballistraceae,
Echinodontium, Echinodontium ryvardenii, Echinodontium tinctorium,
Ectendomeliola,

Endomeliola,
Endothia, Endothia radicalis,
Endothiella, Endothiella gyrosa,
Entorrhizomycetes,
Entyloma, Entyloma ageratinae, Entyloma dahliae, Entyloma ellisii,
Epicoccum, Epicoccum nigrum,
Eremothecium, Eremothecium coryli, Eremothecium gossypii,
Erysiphales,
Exobasidiaceae, Exobasidium burtii, Exobasidium reticulatum, Exobasidium vaccinii var. japonicum, Exobasidium vaccinii-uliginosi, Exobasidium vexans, xxophiala alcalophila,
Exophiala, Exophiala angulospora, Exophiala attenuata, Exophiala calicioides, Exophiala castellanii, Exophiala dermatitidis, Exophiala dopicola, Exophiala exophialae, Exophiala heteromorpha, Exophiala hongkongensis, Exophiala jeanselmei, Exophiala lecanii-corni, Exophiala mansonii, Exophiala mesophila, Exophiala moniliae, Exophiala negronii, Exophiala phaeomuriformis, Exophiala pisciphila, Exophiala psychrophila, Exophiala salmonis, Exophiala spinifera,
Fomes, Fomes

Fomitopsis, Fomitopsis rosea,
Fusicladium pisicola,
Fusicoccum, Fusicoccum aesculi, Fusicoccum amygdali, Fusicoccum quercus,
Galactomyces, Galactomyces candidum,
Ganoderma, Ganoderma brownii, Ganoderma lobatum, Ganoderma megaloma, Ganoderma meredithiae, Ganoderma orbiforme, Ganoderma philippii, Ganoderma sessile, Ganoderma tornatum, Ganoderma zonatum,
Geastrumia, Geastrumia polystigmatis,
Georgefischeriaceae,
Georgefischeriales,
Geosmithia, Geosmithia pallida,
Geotrichum, Geotrichum candidum, Geotrichum klebahnii,
Gibberella, Gibberella acuminata, Gibberella avenacea, Gibberella baccata, Gibberella cyanogena, Gibberella fujikuroi, Gibberella intricans, Gibberella pulicaris, Gibberella stilboides, Gibberella tricincta, Gibberella xylarioides, Gibberella zeae,
Gibellina, Gibellina cerealis,
Gilbertella, Gilbertella persicaria,
Gjaerumiaceae,
Gliocladiopsis, Gliocladiopsis tenuis,
Gliocladium, Gliocladium vermoeseni,
Gloeocercospora, Gloeocercospora sorghi,
Gloeocystidiellum, Gloeocystidiellum porosum,
Gloeophyllum, Gloeophyllum mexicanum, Gloeophyllum trabeum,
Gloeoporus, Gloeoporus dichrous,
Gloeosporium, Gloeosporium cattleyae, Gloeosporium theae-sinensis,
Glomerella, Glomerella cingulata, Glomerella graminicola, Glomerella tucumanensis,
Gnomonia, Gnomonia caryae, Gnomonia comari, Gnomonia dispora, Gnomonia iliau, Gnomonia rubi,
Golovinomyces, Golovinomyces cichoracearum,
Graphiola phoenicis,
Graphiolaceae,
Graphium, Graphium rigidum, Graphium rubrum,
Graphyllium, Graphyllium pentamerum,
Grovesinia, Grovesinia pyramidalis,
Gymnoconia, Gymnoconia nitens,
Gymnopus, Gymnopus dryophilus,
Gymnosporangium, Gymnosporangium kernianum, Gymnosporangium libocedri, Gymnosporangium nelsonii, Gymnosporangium yamadae,
Haematonectria, Haematonectria haematococca,
Hansenula, Hansenula subpelliculosa,
Hapalosphaeria, Hapalosphaeria deformans,
Haplobasidion, Haplobasidion musae,
Helicobasidium, Helicobasidium compactum, Helicobasidium longisporum, Helicobasidium purpureum,
Helicoma, Helicoma muelleri,
Helminthosporium, Helminthosporium cookei, Helminthosporium solani,
Hendersonia, Hendersonia creberrima, Hendersonia theicola,
Hericium, Hericium coralloides,
Heterobasidion, Heterobasidion irregulare, Heterobasidion occidentale,
Hexagonia, Hexagonia hydnoides,
Hymenula, Hymenula affinis,
Hyphodermella, Hyphodermella corrugata,
Hyphodontia, Hyphodontia aspera, Hyphodontia sambuci,
Hypoxylon, Hypoxylon tinctor,
Inonotus, Inonotus arizonicus, Inonotus cuticularis, Inonotus dryophilus, Inonotus hispidus, Inonotus ludovicianus,
Irpex, Irpex destruens, Irpex lacteus,
Kabatiella, Kabatiella caulivora,
Карнал бант,
Вилт акации,
Kretzschmaria, Kretzschmaria zonata,
Kuehneola, Kuehneola uredinis,
Kutilakesa, Kutilakesa pironii,
Laetiporus, Laetiporus ailaoshanensis, Laetiporus baudonii, Laetiporus caribensis, Laetiporus conifericola, Laetiporus cremeiporus, Laetiporus gilbertsonii, Laetiporus huroniensis, Laetiporus montanus, Laetiporus portentosus, Laetiporus zonatus,
Laxitextum, Laxitextum bicolor,
Leandria, Leandria momordicae,
Lentinus, Lentinus tigrinus,
Lenzites, Lenzites betulina, Lenzites elegans,
Leohumicola, Leohumicola atra, Leohumicola incrustata, Leohumicola levissima,
Leptodontidium, Leptodontidium elatius,
Leptographium, Leptographium microsporum,
Leptosphaerulina, Leptosphaerulina crassiasca, Leptosphaerulina trifolii,
Leptothyrium, Leptothyrium nervisedum,
Leptotrochila, Leptotrochila medicaginis,
Leucocytospora, Leucocytospora leucostoma,
Leucostoma, Leucostoma auerswaldii, Leucostoma canker, Leucostoma kunzei, Leucostoma persoonii,
Leveillula, Leveillula compositarum, Leveillula leguminosarum, Leveillula taurica,
Limacinula, Limacinula tenuis,
Linochora, Linochora graminis,
Пыльная головня,
Lopharia, Lopharia crassa,
Lophodermium, Lophodermium aucupariae, Lophodermium schweinitzii,
Macrophoma, Macrophoma mangiferae, Macrophoma theicola,
Macrosporium, Macrosporium cocos,
Magnaporthe, Magnaporthe grisea, Magnaporthe salvinii,
Magnaporthiopsis,
Mamianiella, Mamianiella coryli,
Marasmiellus, Marasmiellus cocophilus, Marasmiellus stenophyllus,
Marasmius, Marasmius crinis-equi, Marasmius sacchari, Marasmius semiustus, Marasmius stenophyllus, Marasmius tenuissimus,
Massarina, Massarina walkeri,
Mauginiella, Mauginiella scaettae,
Melampsora, Melampsora lini, Melampsora occidentalis,
Melanconis, Melanconis carthusiana,
Melanconium, Melanconium juglandinum,
Meliola, Meliola mangiferae, Meliola zangii,
Meruliopsis, Meruliopsis ambigua,
Microascus, Microascus brevicaulis,
Microbotryum, Microbotryum silenes-dioicae, Microbotryum violaceum,
Microsphaera, Microsphaera coryli, Microsphaera diffusa, Microsphaera ellisii, Microsphaera euphorbiae, Microsphaera hommae, Microsphaera penicillata, Microsphaera vaccinii, Microsphaera verruculosa,
Microstroma, Microstroma juglandis,
Moesziomyces, Moesziomyces bullatus,
Moniliophthora, Moniliophthora roreri,
Monilochaetes, Monilochaetes infuscans,
Monochaetia, Monochaetia coryli, Monochaetia mali,
Monographella, Monographella albescens, Monographella cucumerina, Monographella nivalis,
Monosporascus, Monosporascus cannonballus, Monosporascus eutypoides,
Monostichella, Monostichella coryli,
Mucor, Mucor circinelloides, Mucor hiemalis, Mucor mucedo, Mucor paronychius, Mucor piriformis, Mucor racemosus,
Mycena, Mycena citricolor,
Mycocentrospora, Mycocentrospora acerina,
Mycoleptodiscus, Mycoleptodiscus terrestris,
Didymella, Didymella rabiei,
Mycosphaerella, Mycosphaerella recutita, Mycosphaerella rosicola, Mycosphaerella rubi, Mycosphaerella stigmina-platani, Mycosphaerella striatiformans,
Mycovellosiella, Mycovellosiella concors,
Passalora, Passalora fulva,
Mycovellosiella, Mycovellosiella koepkei, Mycovellosiella vaginae,
Myriogenospora, Myriogenospora aciculispora,
Myrothecium, Myrothecium roridum, Myrothecium verrucaria,
Naevala, Naevala perexigua,
Naohidemyces, Naohidemyces vaccinii,
Nectria, Nectria cinnabarina, Nectria ditissima, Nectria foliicola, Nectria mammoidea, Nectria mauritiicola, Nectria peziza, Nectria pseudotrichia, Nectria radicicola, Nectria ramulariae,
Nectriella, Nectriella pironii,
Nemania, Nemania diffusa, Nemania serpens,
Neocosmospora, Neocosmospora vasinfecta,
Neodeightonia, Neodeightonia phoenicum,
Neoerysiphe, Neoerysiphe galeopsidis,
Neofabraea, Neofabraea perennans,
Neofusicoccum, Neofusicoccum mangiferae,
Oidiopsis, Oidiopsis gossypii,
Oidium, Oidium arachidis, Oidium caricae-papayae, Oidium indicum, Oidium mangiferae, Oidium manihotis,
Olpidium, Olpidium brassicae,
Omphalia, Omphalia tralucida,
Ophiobolus, Ophiobolus anguillides, Ophiobolus cannabinus,
Ophioirenina,
Ovulinia, Ovulinia azaleae,
Oxyporus, Oxyporus corticola,
Ozonium, Ozonium texanum,
Peltaster, Peltaster fructicola,
Penicillium, Penicillium expansum, Penicillium funiculosum,
Periophora,
Periconia, Periconia circinata,
Periconiella, Periconiella cocoes,
Peridermium, Peridermium californicum,
Pestalosphaeria, Pestalosphaeria concentrica,
Pestalotia, Pestalotia longiseta, Pestalotia rhododendri,
Pestalotiopsis, Pestalotiopsis adusta, Pestalotiopsis arachidis, Pestalotiopsis disseminata, Pestalotiopsis guepini, Pestalotiopsis leprogena, Pestalotiopsis longiseta, Pestalotiopsis mangiferae, Pestalotiopsis palmarum, Pestalotiopsis sydowiana, Pestalotiopsis theae,
Peyronellaea, Peyronellaea curtisii,
Phacidiopycnis, Phacidiopycnis padwickii,
Phaeochoropsis, Phaeochoropsis mucosa,
Phaeocytostroma, Phaeocytostroma iliau, Phaeocytostroma sacchari,
Phaeoisariopsis, Phaeoisariopsis bataticola,
Phaeoramularia, Phaeoramularia heterospora, Phaeoramularia indica, Phaeoramularia manihotis,
Phaeoseptoria, Phaeoseptoria musae,
Phaeosphaerella, Phaeosphaerella mangiferae, Phaeosphaerella theae,
Phaeosphaeria, Phaeosphaeria avenaria, Phaeosphaeria herpotrichoides, Phaeosphaeria microscopica, Phaeosphaeria nodorum,
Phaeosphaeriopsis, Phaeosphaeriopsis obtusispora,
Phaeotrichoconis, Phaeotrichoconis crotalariae,
Phialophora, Phialophora asteris, Phialophora cinerescens, Phialophora gregata, Phialophora tracheiphila,
Phoma, Phoma clematidina, Phoma costaricensis, Phoma cucurbitacearum, Phoma destructive, Phoma draconis, Phoma exigua, Phoma exigua, Phoma exigua var. foveata, Phoma exigua, Phoma glomerata, Phoma glycinicola, Phoma herbarum, Phoma insidiosa, Phoma medicaginis, Phoma microspora, Phoma narcissi, Phoma nebulosa, Phoma oncidii-sphacelati, Phoma pinodella, Phoma sclerotioides, Phoma strasseri,
Phomopsis, Phomopsis asparagi, Phomopsis asparagicola, Phomopsis cannabina, Phomopsis coffeae, Phomopsis ganjae, Phomopsis javanica, Phomopsis longicolla, Phomopsis mangiferae, Phomopsis prunorum, Phomopsis sclerotioides, Phomopsis theae,
Phragmidium, Phragmidium mucronatum, Phragmidium rosae-pimpinellifoliae, Phragmidium rubi-idaei, Phragmidium violaceum,
Phyllachora, Phyllachora banksiae, Phyllachora cannabis, Phyllachora graminis, Phyllachora gratissima, Phyllachora musicola, Phyllachora pomigena, Phyllachora sacchari,
Phyllactinia,
Phyllosticta, Phyllosticta alliariaefoliae Phyllosticta arachidis-hypogaeae, Phyllosticta batatas, Phyllosticta capitalensis, Phyllosticta carpogena, Phyllosticta coffeicola, Phyllosticta concentrica, Phyllosticta coryli, Phyllosticta cucurbitacearum, Phyllosticta cyclaminella, Phyllosticta erratica, Phyllosticta hawaiiensis, Phyllosticta lentisci, Phyllosticta manihotis, Phyllosticta micropuncta, Phyllosticta mortonii, Phyllosticta nicotianae, Phyllosticta palmetto, Phyllosticta penicillariae, Phyllosticta perseae, Phyllosticta pseudocapsici, Phyllosticta sojaecola, Phyllosticta theae, Phyllosticta theicola,
Phymatotrichopsis, Phymatotrichopsis omnivora,
Physalospora, Physalospora disrupta, Physalospora perseae,
Physoderma, Physoderma alfalfae, Physoderma leproides, Physoderma trifolii,
Physopella, Physopella ampelopsidis,
Pileolaria, Pileolaria terebinthi,
Piricaudiopsis, Piricaudiopsis punicae, Piricaudiopsis rhaphidophorae, Piricaudiopsis rosae,
Plenodomus, Plenodomus destruens, Plenodomus meliloti,
Pleosphaerulina, Pleosphaerulina sojicola,
Pleospora, Pleospora alfalfae, Pleospora betae, Pleospora herbarum, Pleospora lycopersici, Pleospora tarda, Pleospora theae,
Pleuroceras,
Podosphaera, Podosphaera fuliginea, Podosphaera fusca, Podosphaera leucotricha, Podosphaera macularis, Podosphaera pannosa,
Polyscytalum, Polyscytalum pustulans,
Poria, Poria hypobrunnea, Postia, Postia tephroleuca, Настоящая мучнистая роса,
Pseudocercospora, Pseudocercospora arecacearum, Pseudocercospora cannabina, Pseudocercospora fuligena, Pseudocercosporella herpotrichoides, Pseudocercospora gunnerae, Pseudocercospora pandoreae, Pseudocercospora puderi, Pseudocercospora rhapisicola, Pseudocercospora theae, Pseudocercospora vitis, Pseudocercosporella capsellae,
Pseudocochliobolus, Pseudocochliobolus eragrostidis,
Pseudoepicoccum, Pseudoepicoccum cocos,
Pseudopeziza, Pseudopeziza jonesii, Pseudopeziza medicaginis, Pseudopeziza trifolii,
Pseudoseptoria, Pseudoseptoria donacis,
Pucciniaceae,
Pucciniastrum, Pucciniastrum americanum, Pucciniastrum arcticum, Pucciniastrum epilobii, Pucciniastrum hydrangeae,
Pycnostysanus, Pycnostysanus azaleae,
Pyrenochaeta, Pyrenochaeta lycopersici, Pyrenochaeta terrestris,
Pyrenopeziza, Pyrenopeziza brassicae,
Ramichloridium, Ramichloridium musae,
Ramulispora, Ramulispora sorghi, Ramulispora sorghicola,
Rhinocladium, Rhinocladium corticola,
Rhizophydium, Rhizophydium graminis,
Rhizopus, Rhizopus arrhizus, Rhizopus circinans, Rhizopus microsporus, Rhizopus oryzae,
Rhytisma, Rhytisma punctatum, Rhytisma vitis, Rigidoporus, Rigidoporus vinctus,
Rosellinia, Rosellinia arcuata, Rosellinia bunodes, Rosellinia necatrix, Rosellinia pepo,
Saccharicola, Saccharicola taiwanensis,
Schiffnerula, Schiffnerula cannabis,
Schizophyllum, Schizophyllum commune,
Schizopora, Schizopora flavipora,
Schizothyrium, Schizothyrium pomi,
Sclerophthora, Sclerophthora macrospora,
Sclerotium, Sclerotium cinnamomi, Sclerotium delphinii,
Scytinostroma, Scytinostroma galactinum,
Seimatosporium, Seimatosporium mariae, Seimatosporium rhododendri,
Selenophoma, Selenophoma linicola,
Septobasidium, Septobasidium bogoriense, Septobasidium euryae-groffii, Septobasidium gaoligongense, Septobasidium pilosum, Septobasidium polygoni, Septobasidium pseudopedicellatum, Septobasidium theae,
Septocyta, Septocyta ruborum,
Serpula, Serpula lacrymans,
Setosphaeria, Setosphaeria rostrata, Setosphaeria turcica,
Spencermartinsia, Spencermartinsia pretoriensis,
Sphaceloma, Sphaceloma arachidis, Sphaceloma menthae, Sphaceloma perseae, Sphaceloma poinsettiae, Sphaceloma sacchari, Sphaceloma theae,
Sphacelotheca, Sphacelotheca reiliana, Sphaerotheca castagnei,
Sphaerulina, Sphaerulina oryzina, Sphaerulina rehmiana, Sphaerulina rubi,
Sphenospora, Sphenospora kevorkianii,
Spilocaea, Spilocaea oleaginea,
Sporisorium, Sporisorium omentum, Sporisorium ehrenbergii, Sporisorium scitamineum, Sporisorium sorghi,
Sporonema, Sporonema phacidioides,
Stagonospora, Stagonospora avenae, Stagonospora meliloti, Stagonospora recedens, Stagonospora sacchari, Stagonospora tainanensis,
Stagonosporopsis,
Stegocintractia, Stegocintractia junci,
Stemphylium, Stemphylium alfalfae, Stemphylium bolickii, Stemphylium cannabinum, Stemphylium globuliferum, Stemphylium lycopersici, Stemphylium sarciniforme, Stemphylium solani, Stemphylium vesicarium,
Stenella, Stenella anthuriicola,
Stigmatomycosis,
Stigmina, Stigmina carpophila, Stigmina palmivora, Stigmina platani-racemosae,
Stromatinia, Stromatinia cepivora,
Sydowiella, Sydowiella depressula,
Sydowiellaceae,
Synchytrium, Synchytrium endobioticum,
Tapesia, Tapesia acuformis, Tapesia yallundae,
Taphrina, Taphrina coryli, Taphrina potentillae,
Thanatephorus, Thanatephorus cucumeris,
Thecaphora, Thecaphora solani,
Thielaviopsis, Thielaviopsis basicola, Thielaviopsis ceramica,
Thyrostroma, Thyrostroma compactum,
Tiarosporella, Tiarosporella urbis-rosarum,
Tilletia, Tilletia barclayana, Tilletia caries, Tilletia controversa, Tilletia laevis, Tilletia tritici, Tilletia walkeri,
Tilletiariaceae,
Togniniaceae,
Tranzschelia, Tranzschelia pruni-spinosae,
Trichoderma, Trichoderma koningii, Trichoderma paucisporum, Trichoderma songyi, Trichoderma theobromicola, Trichoderma viride,
Tubercularia, Tubercularia lateritia,
Tunstallia, Tunstallia aculeata,
Typhula, Typhula blight, Typhula idahoensis, Typhula incarnata, Typhula ishikariensis, Typhula variabilis,
Ulocladium, Ulocladium consortiale,
Uncinula,
Uredo, Uredo behnickiana, Uredo kriegeriana, Uredo musae, Uredo nigropuncta, Uredo rangelii,
Urocystis, Urocystis agropyri, Urocystis brassicae, Urocystis occulta,
Uromyces, Uromyces apiosporus, Uromyces appendiculatus, Uromyces beticola, Uromyces ciceris-arietini, Uromyces dianthi, Uromyces euphorbiae, Uromyces graminis, Uromyces inconspicuus, Uromyces lineolatus, Uromyces musae, Uromyces oblongus, Uromyces pisi-sativi, Uromyces
Urophlyctis, Urophlyctis leproides, Urophlyctis trifolii,
Ustilaginales,
Ustilago, Ustilago avenae, Ustilago esculenta, Ustilago hordei, Ustilago maydis, Ustilago nigra, Ustilago nuda, Ustilago scitaminea, Ustilago tritici,
Vankya, Vankya ornithogali,
Вельветовая болезнь,
Veronaea, Veronaea musae,
Verticillium, Verticillium albo-atrum, Verticillium alfalfae, Verticillium dahliae, Verticillium isaacii, Verticillium klebahnii, Verticillium longisporum, Verticillium nonalfalfae, Verticillium theobromae, Verticillium wilt, Verticillium zaregamsianum,
Waitea, Waitea circinata,
Westea,
Листовая ржавчина пшеницы,
Мучнистая роса пшеницы,
Wuestneiopsis, Wuestneiopsis georgiana,
Xeromphalina, Xeromphalina fraxinophila,
Zopfia, Zopfia rhizophila,
Zygosaccharomyces, Zygosaccharomyces bailii, Zygosaccharomyces florentinus,
Zythiostroma.
Предпочтительно пары грибов, оомицетов или бактерий и сельскохозяйственных культур, охваченные изобретением, являются следующими:
Пшеница (Thticum sativum)
Claviceps purpurea, Erysiphe graminis, Fusarium avenaceum, Fusarium culmorum, Fusarium graminearum, Fusarium langsethiae, Fusarium poae, Fusarium pseudograminearum, Gaeumannomyces graminis, Leptosphaeria nodorum, Microdochium spp., Mycosphaerella graminicola, Oculimacula acuformis, Oculimacula yallundae, Puccinia recondita, Puccinia striiformis, Pyrenophora tritici-repentis, Rhizoctonia cerealis, Microdochium и Zymoseptoria tritici
Кукуруза (Zea mays)
Fusarium graminearum, Fusarium proliferatum, Fusarium subglutinans, Fusarium verticillioides
Ячмень (Hordeum vulgare)
Blumeria graminis, Fusarium spp., Pyrenophora teres, Ramularia collo-cygni, Rhynchosporium secalis
Рис (Oryza sativa)
Cochliobolus miyabeanus, Fusarium

Картофель (Solanum tuberosum)
Alternia alternate, Alternaria solani, Phytophtora infestans, Rhizoctonia solani
Виноградная лоза (Vinis vitifera)
Botrytis cinerea, Erysiphe necator, Plasmopara viticola, Guignardia bidwelli, Erisyphe necator, Diplodia seriata
Соя (Glycine max)
Cercopora kikuchii, Colletotrichum dematium, Corynespora cassiicola, Fusarium graminearum, Pythium spp., Rhizoctonia solani, Sclerotinia sclerotiorum, Septoria glycines
Яблоня (Malus domestica)
Monilia fructigena, Monilia laxa, Pezzicula alba, Pezzicula malicorticis, Venturia inaequalis
Томат (Lycopersicon esculentum)
Phytophtora infestans
Фасоль (Phaseolus vulgaris)
Uromyces appendiculatus
Редис (Raphanus sativus)
Alternaria brassicae
Все фрукты и овощи
Botrytis cinerea
Земляника (Fragaria sp.)
Colletotrichum acutatum
Морковь (Daucus carota)
Aternaria alternate, Alternaria dauci, Alternaria radicina
Персик (Prunus persica) и абрикос (Prunus armeniaca)
Monilia fructicola, Monilia fructigena, Monilia laxa
Особенно предпочтительно пары грибов или бактерий и сельскохозяйственных культур, охваченные изобретением, являются следующими:
Пшеница: Fusarium graminearum, Microdochium и Zymoseptoria tritici
Виноградная лоза: Botrytis cinerea, Erysiphe necator, Plasmopara viticola, Guignardia bidwelli, Erisyphe necator, Diplodia seriata
Картофель: Alternia alternate, Alternerie soleni, Phytophtore infestens, Rhizoctonie soleni
Томат: Phytophtore infestens
Способ получения
Изобретение также относится к способу получения клеточного экстракта одной или более микроскопической(-их) водоросли(-ей) рода Amphidinium, характеризующемуся следующими стадиями:
- сбор свежих клеток одной или более микроскопической(-их) водоросли(-ей) рода Amphidinium;
- необязательно замораживание и/или лиофилизация указанных клеток:
- суспендирование указанных свежих или замороженных клеток или лиофилизата в неорганическом или органическом растворителе в массовом соотношении лиофилизат или биомасса/растворитель, составляющем от 1:200 до 1:2;
- необязательно лиофилизация полученного экстракта.
«Неорганический растворитель» означает воду и водные растворы (растворы в воде), а «органический растворитель» означает углеводородные растворители (алифатические, ароматические), окисленные растворители (спирты, кетоны, кислоты, сложные эфиры и простые эфиры), галогенированные растворители (дихлорметан, хлороформ) и смеси этих растворителей в любом смешиваемом соотношении.
Предпочтительно будут использовать воду или окисленный растворитель, предпочтительно С1-С4 спирт, такой как метанол или этанол.
В случае органического растворителя суспендирование можно выполнять при температуре от 4 до 60°С, предпочтительно от 18 до 60°С, особенно предпочтительно при комнатной температуре.
Предпочтительно суспендирование свежих или замороженных клеток или лиофилизата в неорганическом растворителе проводят при температуре выше 60°С, предпочтительно выше 80°С, особенно предпочтительно при температуре выше 90°С. Предпочтительно свежие или замороженные клетки или лиофилизат в неорганическом растворителе суспендируют менее 5 минут, предпочтительно менее 3 минут, предпочтительно менее 1 минуты. Предпочтительно температуру впоследствии быстро возвращают к комнатной температуре. Предпочтительно температуру смеси возвращают к температуре, близкой к комнатной, помещая смесь в условия охлаждения, например, при температуре, близкой к 0°С, или добавляя к смеси неорганический растворитель при температуре, близкой к 0°С.
Суспендирование выполняют либо путем добавления к указанным свежим или замороженным клеткам или к указанному лиофилизату растворителя, предварительно нагретого до желаемой температуры, или добавляют растворитель, и ресуспендированную смесь доводят до желаемой температуры.
Предпочтительно суспендирование указанных свежих или замороженных клеток или указанного лиофилизата в неорганическом или органическом растворителе проводят при массовом соотношении лиофилизат или биомасса/растворитель, составляющем от 1:100 до 1:50.
«Быстрый» возврат температуры к комнатной означает в течение менее 5 минут, предпочтительно в течение менее 3 минут, предпочтительно в течение менее 1 минуты.
Изобретение также относится к клеточному экстракту или лиофилизату клеток одной или более микроскопических водорослей рода Amphidinium, который может быть получен способом получения клеточного экстракта по изобретению.
Преимущественно свежие клетки, которые собирают и экстрагируют, получают из культуры клеток в условиях температуры, светового периода и солености, подходящих для рассматриваемого штамма, выращенной до концентрации клеток, составляющей от 5⋅104 клеток/мл до 5⋅106 клеток/мл, предпочтительно до концентрации клеток, составляющей от 5⋅105 клеток/мл до 1⋅106 клеток/мл.
Клетки культивируют в течение от 5 до 20 дней.
Интенсивность света составляет от 40 микроэйнштейн (мкЭ) до 200 мкЭ, предпочтительно составляет от 70 мкЭ до 100 мкЭ.
Температура культивирования обычно составляет от 17°С до 25°С.
Световой период день/ночь предпочтительно составляет от 8 ч/16 ч до 16 ч/8 ч.
Минимальная содержание соли составляет 15 частей на тысячу (ч. н. т.).
В соответствии с конкретным воплощением изобретения, Amphidinium carterae культивируют, как описано ниже: клетки инкубируют в природной или искусственной морской водной среде при температуре, составляющей от 17 до 25°С, при этом цикл день/ночь составляет от 8 ч/16 ч до 16 ч/8 ч, предпочтительно 16 ч/8 ч.
Способ борьбы
Изобретение также относится к способу борьбы с патогенными грибами, оомицетами и/или бактериями сельскохозяйственных культур и семян, включающему нанесение на сельскохозяйственные культуры клеточного экстракта одной или более микроскопических водорослей рода Amphidinium или экстракта в соответствии с изобретением.
Эти меры борьбы могут быть лечебными или профилактическими, предпочтительно лечебными.
Изобретение также относится к способу борьбы с патогенными грибами, оомицетами и/или бактериями сельскохозяйственных культур и семян, включающему следующие стадии:
- перемешивание клеточного экстракта в воде в соотношении от 1:4 до 1:800 непосредственно перед применением;
- нанесение этой смеси на сельскохозяйственные культуры и/или покрытие данной смесью семян.
Изобретение также относится к способу борьбы с патогенными грибами, оомицетами и/или бактериями сельскохозяйственных культур и семян, включающему следующие стадии:
- ресуспендирование лиофилизата культуры клеток одной или более микроводорослей рода Amphidinium в массовой концентрации, составляющей от 5 до 500 г/л, предпочтительно в массовой концентрации, составляющей от 50 до 400 г/л, предпочтительно в массовой концентрации, составляющей от 100 до 200 г/л, в воде или в органическом или неорганическом растворителе в массовом соотношении от 1:200 до 1:2;
- перемешивание в воде непосредственно перед применением в соотношении от 1:4 до 1:800.
- нанесение полученного экстракта на сельскохозяйственные культуры и/или покрытие семян этой смесью.
В случае суспендирования в воде или в неорганическом растворителе суспендирование предпочтительно проводят при температуре выше 60°С, а затем температуру возвращают к температуре, близкой к комнатной, путем разведения в воде в соотношении от 1:2 до 1:50.
Нанесение на сельскохозяйственные культуры можно выполнять любыми средствами, известными специалисту в данной области техники, для достижения частей растений, пораженных грибом и/или бактерией.
Экстракт наносят в дозе, составляющей от 0,005 г/л до 20 г/л, предпочтительно от 0,5 г/л до 10 г/л, особенно предпочтительно от 1 г/л до 5 г/л.
Покрытие семян можно выполнять любым методом, известным специалисту в данной области техники, позволяющим активному агенту оставаться в контакте с семенами.
Например, покрытие можно выполнять путем посыпания порошком или опрыскивания.
Например, покрытие может содержать компоненты рецептуры и добавки.
Компоненты рецептуры используют для того, чтобы дать возможность нанесения активного(-ых) вещества (веществ), и для того, чтобы оно (они) оставалось (оставались) на семени в равном и постоянном соотношении на протяжении всего процесса нанесения препарата в очень низких дозах.
Компоненты рецептуры включают: органические растворители или воду, диспергирующие агенты, эмульгаторы, поверхностно-активные вещества (ПАВ) или смачивающие агенты, красители и т.д.
ПАВ и эмульгаторы обладают свойством объединения и стабильного удерживания вместе двух несовместимых жидкостей.
На семена можно наносить различные вспомогательные вещества. Пленочные покрытия соответствуют нанесению микропористой пленки на поверхность семян. Эти пленки не изменяют форму и размер семени. Они улучшают покрывающую способность и однородность обработки. Обращение с семенами при их использовании фермером во время посева упрощается за счет подавления образования пыли и улучшения сыпучести семян в сеялке. Они улучшают действие активного(-ых) вещества (веществ) в условиях культивирования. Покрытия изменяют форму, размер и массу семян. Они улучшают точность посева.
Способы обработки для борьбы с патогенными грибами и/или бактериями сельскохозяйственных культур и семян в соответствии с изобретением особенно пригодны против фузариозной болезни, предпочтительно фузариозных болезней, представленных в таблице 1.


Способы обработки для борьбы с патогенными грибами, оомицетами и/или бактериями сельскохозяйственных культур и семян в соответствии с изобретением особенно пригодны для следующих пар грибов или бактерий и сельскохозяйственных культур:
Пшеница: Fusarium graminearum, Microdochium nivale и Zymoseptoria tritici
Виноградная лоза: Botrytis cinerea, Plasmopara viticola, Guignardia bidwelli, Erisyphe necator, Diplodia seriata
Яблоня: Venturia inaequalis
Банановое дерево: Fusarium oxysporum и Mycosphaerella fijiensis
ПРИМЕРЫ
МАТЕРИАЛЫ И МЕТОДЫ
Пример 1: Культура микроскопической водоросли.
Микроскопическая водоросль Amphidinium carterae, штамм АС208, получена из коллекции Algobank (Caen), а микроскопические водоросли Prymnesium parvum, штамм RCC 1436, и Phaeodactylum tricornutum, штамм ССМР 632, получены из банка морских микроорганизмов г. Роскоф (RCC: Roscoff Culture Collection). Эти микроскопические водоросли культивируют в искусственной морской воде L1 (https://ncma.bigelow.org/algal-recipes) при 19°С и цикле день/ночь 12 ч/12 ч. Используемая интенсивность света составляет 100 мкЭ. Биомассу выделяют в конце экспоненциальной фазы роста путем центрифугирования (15 мин при 3000 об/мин). Полученный в результате осадок клеток замораживают, а затем лиофилизируют, используя лабораторный лиофилизатор (Alpha 1-2 LDplus, Labconco), для стабильного долгосрочного консервирования активной субстанции. После лиофилизации сухое вещество взвешивают.
Пример 2: Получение экстракта.
Для экстракции активного вещества из сухого вещества примера 1 ресуспендируют 20 мг сухого вещества в 1 мл дистиллированной воды при 100°С. После инкубации в течение 2 минут при комнатной температуре (20-25°С) экстракт хранят во льду, а затем центрифугируют в течение 5 мин при 10000 об/мин при комнатной температуре. Супернатант, содержащий активное вещество, замораживают в жидком азоте для долгосрочного консервирования его противогрибковых свойств.
Пример 3: Испытание на прорастание Fusarium graminearum
Споры Fusarium graminearum выращивают в истощенной питательной среде на основе золотистой фасоли. Споры отделяют от мицелия фильтрованием на фильтре Miracloth (Calbiochem), центрифугируют, а затем ресуспендируют в концентрации 1,6⋅106 спор/мл. Приблизительно 16000 спор инкубируют в присутствии контрольного раствора или экстракта A. carterae в различных концентрациях. После инкубации в течение 10 мин при комнатной температуре споры помещают на предметное стекло для подсчета проросших спор через 6 часов или на чашку Петри для наблюдения за ростом мицелия через 72 часа.
ИСПЫТАНИЯ
Экстракты различных видов морских микроорганизмов, относящихся к трем основным типам фитопланктона - динофлагеллятам, гаптофитовым водорослям и диатомовым водорослям - подвергали испытанию на потенциальную противогрибковую активность в отношении криптогамных грибов. Эти микроскопические водоросли обладают способностью продуцировать токсины, которые обеспечивают их активную пролиферацию за счет конкуренции с другими видами и поэтому являются потенциальными источниками молекул, которые могут проявлять противогрибковую активность. Экстракты каждой микроскопической водоросли были получены в соответствии с примером 2. Для испытания действия этих экстрактов на выживаемость фитопатогенных грибов лиофилизированные экстракты ресуспендируют в воде и приводят в контакт с определенным количеством спор Fusarium graminearum.
Пример 4: Ингибирование роста и прорастания спор Fusarium graminearum.
Способность F. graminearum к формированию мицелия на агаровой питательной среде в присутствии этих экстрактов определяют через 72 часа. Из трех исследованных экстрактов (Prymnesium parvum, Amphidinium carterae и Phaeodactylum tricornutum) ингибиторным действием на формирование мицелия обладает только экстракт Amphidinium carterae в соответствии с примером 2 (Фиг. 1А).
Для подтверждения этого результата было проведено испытание доза-ответ путем инкубации спор с экстрактом A. carterae в соответствии с примером 2, разведенным до различных концентраций (Фиг. 1В). Оказалось, что этот экстракт обладает противогрибковым действием, которое является дозозависимым при минимальной ингибиторной концентрации (МИК) 0,4 г/л (Фиг. 1В).
Наконец, было проведено испытание экстракта A. carterae в соответствии с примером 2 на ингибирование прорастания спор F. graminearum. Результаты, полученные in vitro через 6 часов после инкубации с экстрактом, показывают полное ингибирование прорастания спор при концентрации 2 г/л (Фиг. 1С), что позволяет предположить, что этот экстракт ингибирует прорастание спор и рост мицелия F. graminearum.
Пример 5: Влияние лиофилизации экстракта A. carterae на противогрибковую активность экстракта.
Авторы изобретения определяли, ингибирует ли лиофилизация экстракта A. carterae противогрибковую активность. С этой целью культуру A. carterae экстрагировали в соответствии с примером 2, а затем половину экстракта замораживали при -80°С, а вторую половину экстракта лиофилизировали, а затем ресуспендировали в дистиллированной воде. Проводили испытание этих экстрактов на их способность ингибировать рост F. graminearum при различных концентрациях в соответствии с примером 3. Результаты показывают, что в обоих случаях, замороженного экстракта или лиофилизированного экстракта, получили полное ингибирование роста F. graminearum при концентрации 1 г/л, которое сохраняется вплоть до 5 г/л (Фиг. 1D). В заключение, лиофилизация экстракта A. carterae не оказывает никакого влияния на его противогрибковую активность.
Пример 6: Испытания на растениях пшеницы, инфицированных в контролируемых условиях
Колосья пшеницы инокулировали спорами F. graminearum в контролируемых условиях, а затем спустя 24 часа наносили экстракт A. carterae примера 2. Симптомы регистрировали через 20 дней (400°D) и через 22 дня (450°D) после инфекции (Фиг. 2А). В обоих случаях указывали количество колосьев с симптомами (заболеваемость) и балльную оценку симптомов (выраженность болезни, баллы 0-9): в отсутствие (контроль) и в присутствии экстракта примера 2. По сравнению с контрольной обработкой присутствие экстракта значительно снижает количество пораженных болезнью колосьев приблизительно на 30%. Кроме того, симптомы на этих колосьях проявляются с интенсивностью, сниженной приблизительно на 50% (Фиг. 2В). Эти результаты демонстрируют, что экстракт A. carterae примера 2 обладает значительной противогрибковой активностью в отношении роста фитопатогенных грибов пшеницы в условиях in vitro и in planta.
Пример 7: Испытания на нескольких семействах фитопатогенов виноградной лозы: Botrytis cinerea, аскомицета, вызывающего серо-бурую гниль, Plasmopara viticola, оомицета, вызывающего ложную мучнистую росу, Erisyphe necator, аскомицета, вызывающего настоящую мучнистую росу, и Diplodia seriata, одного из возбудителей болезни древесины эска.
В концентрации 1 г/л экстракт A. carterae примера 2 полностью ингибирует рост Р. viticola на отсоединенных листьях (Фиг. 3А, слева), при этом в тех же условиях экстракт не влияет на рост Е. necator (Фиг. 3А, справа). Испытания in vitro, проведенные на Botrytis cinerea, показали, что экстракт полностью ингибирует рост В. cinerea в концентрации 5 г/л (Фиг. 3В). Противогрибковая активность экстракта в отношении В. cinerea была подтверждена на отсоединенных листьях виноградной лозы (Фиг. 3В). В дополнение к этому были проведены испытания in vitro на различных семействах грибов, вызывающих болезнь эска виноградной лозы. Результаты показывают, что экстракт в концентрации 2 г/л в значительной степени ингибирует рост этих грибов (Фиг. 3С).
Пример 8: Испытания in vitro на Microdochium majus, Fusarium graminearum, Zymoseptoria tritici, Fusarium oxysporum, Rhizoctonia solani и Phytophthora infestans
Чтобы определить, обладает ли экстракт A. carterae в соответствии с примером 2, противогрибковой активностью в отношении широкого спектра растительных грибов, были проведены испытания на других грибах, вызывающих фузариозы: Fusarium oxysporum (фузариозный вилт бананового дерева) и Microdochium (красная фузариозная гниль пшеницы), а также другую важную болезнь пшеницы - септориозную пятнистость листьев, вызываемую Zymoseptoria tritici. Испытания были также проведены для двух патогенных организмов картофеля: базидиомицета Rhizoctonia solani и оомицета Phytophthora infestans. Во всех случаях экстракт A. carterae в соответствии с примером 2 в значительной степени ограничивал рост грибов и оомицетов в условиях in vitro (таблицы 2-6).

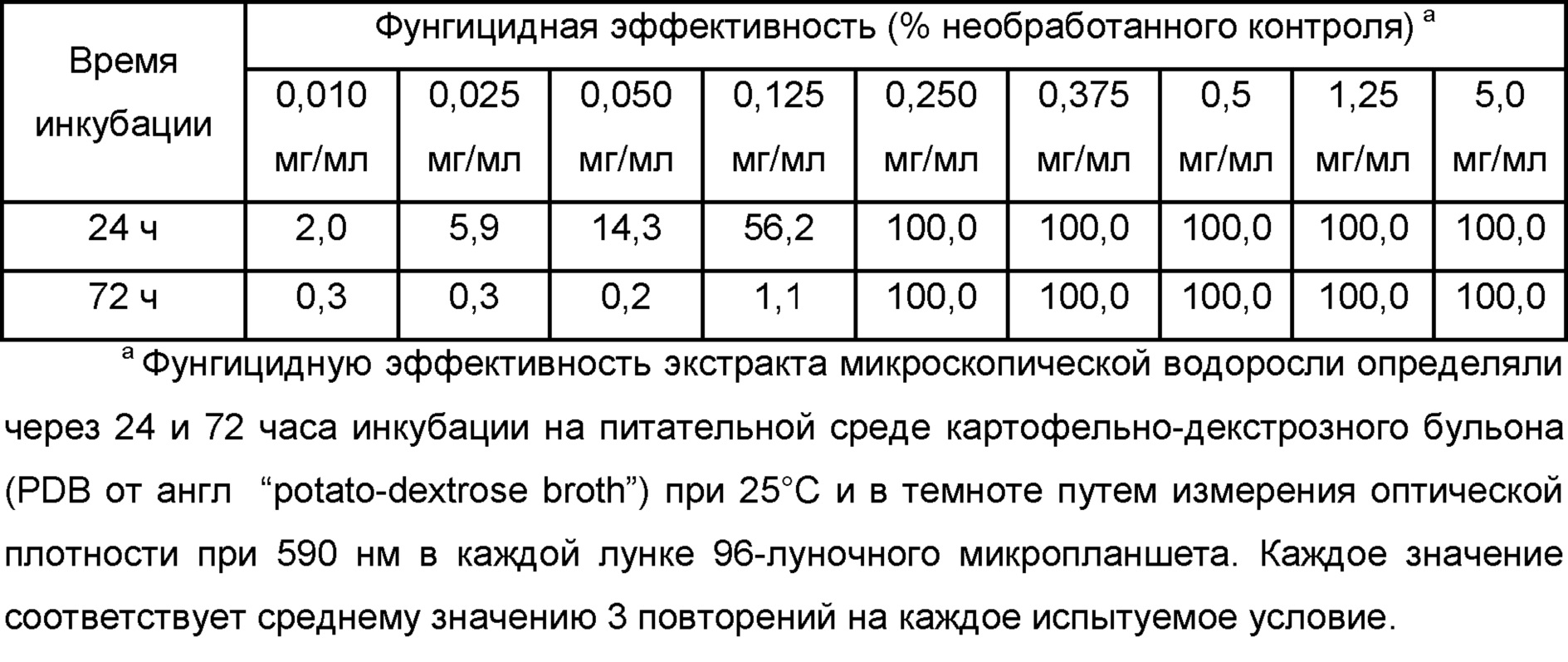
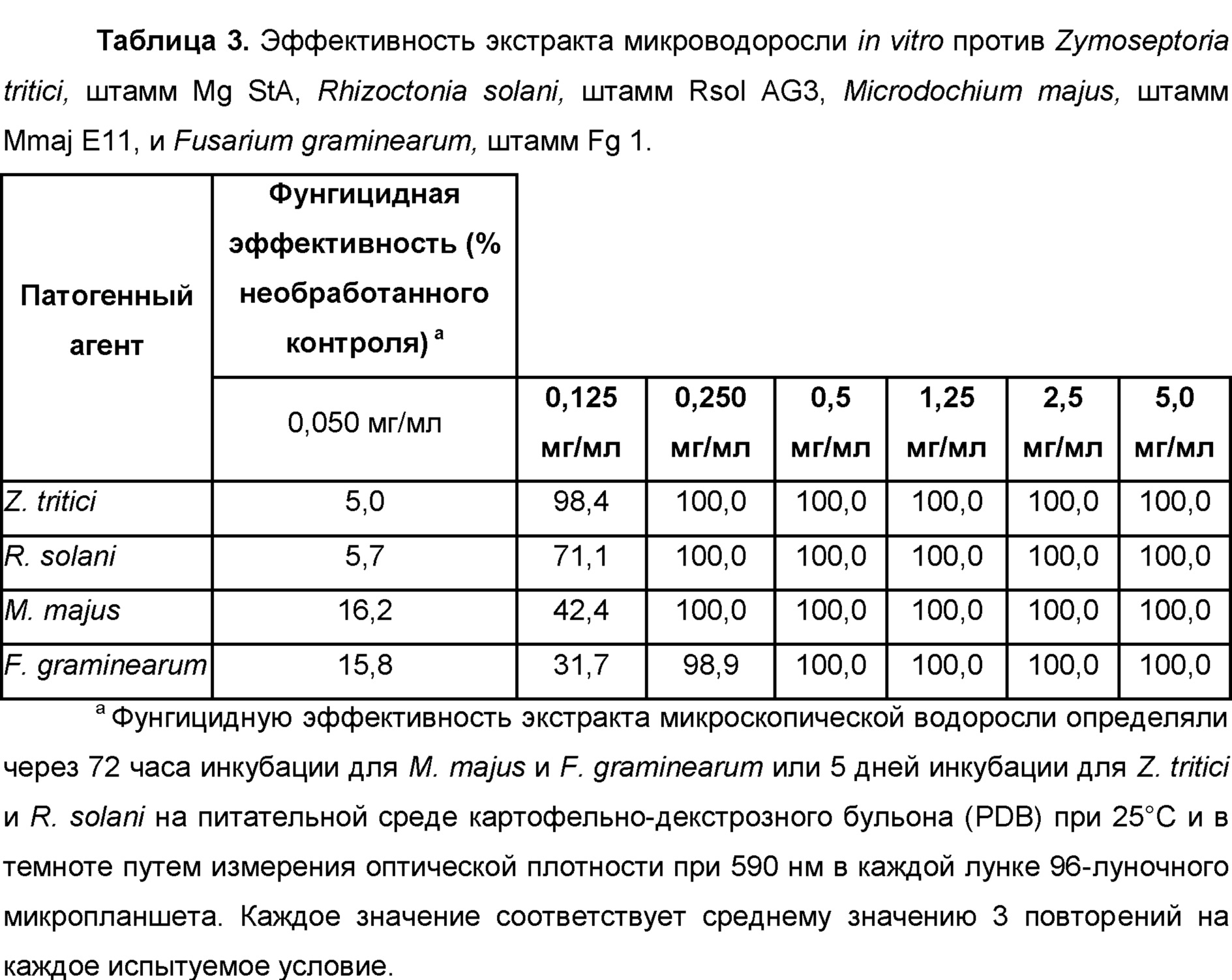

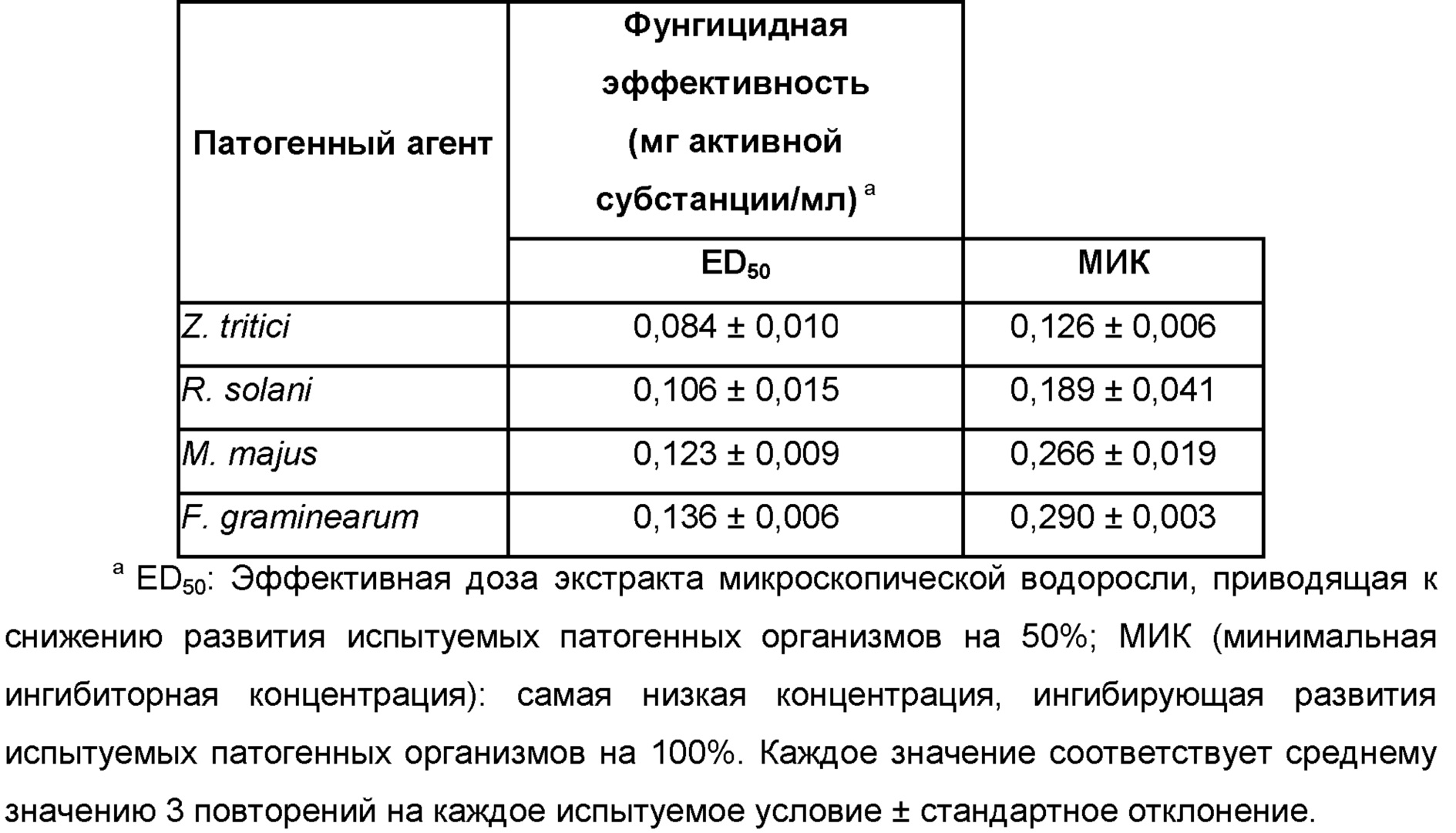
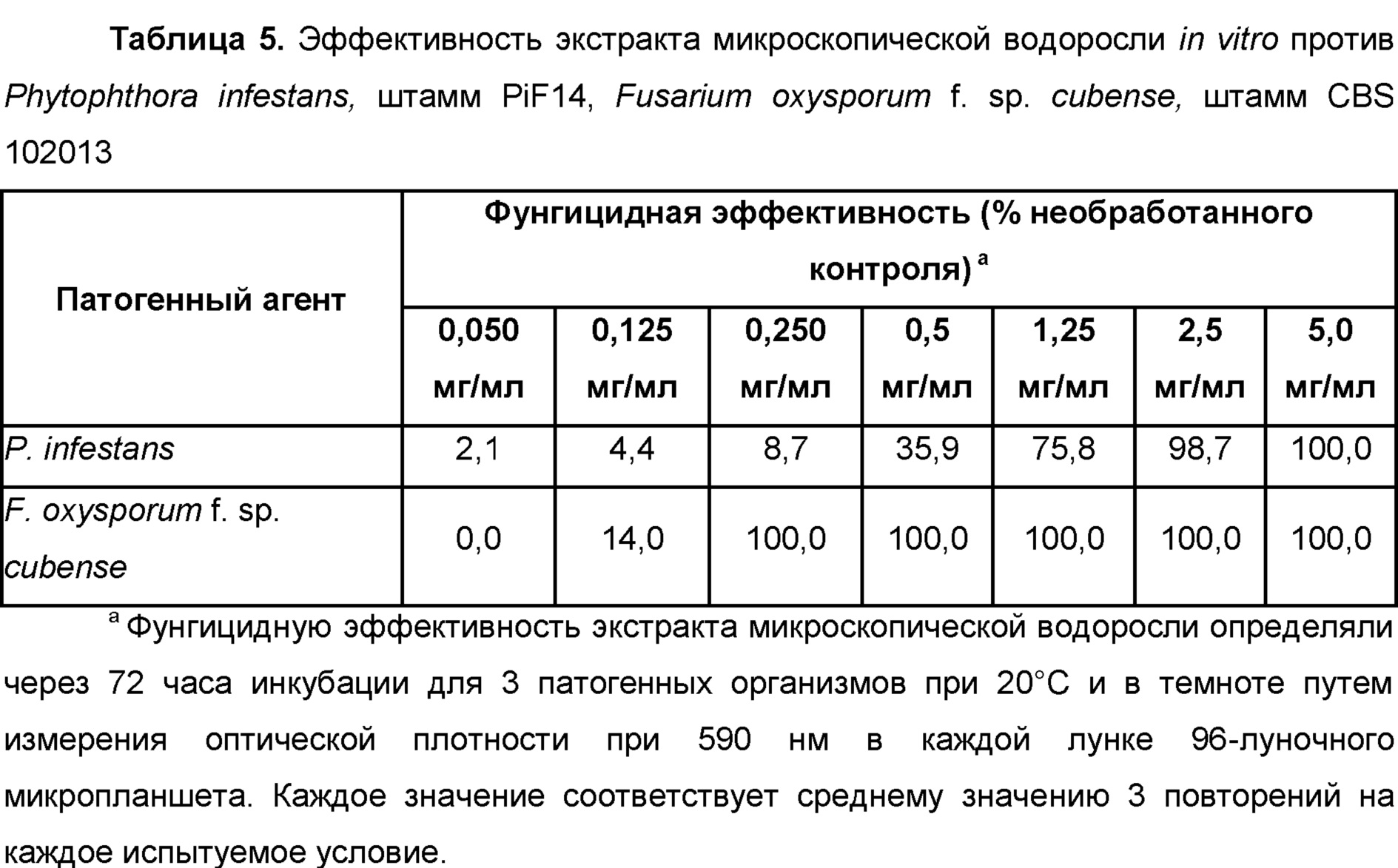

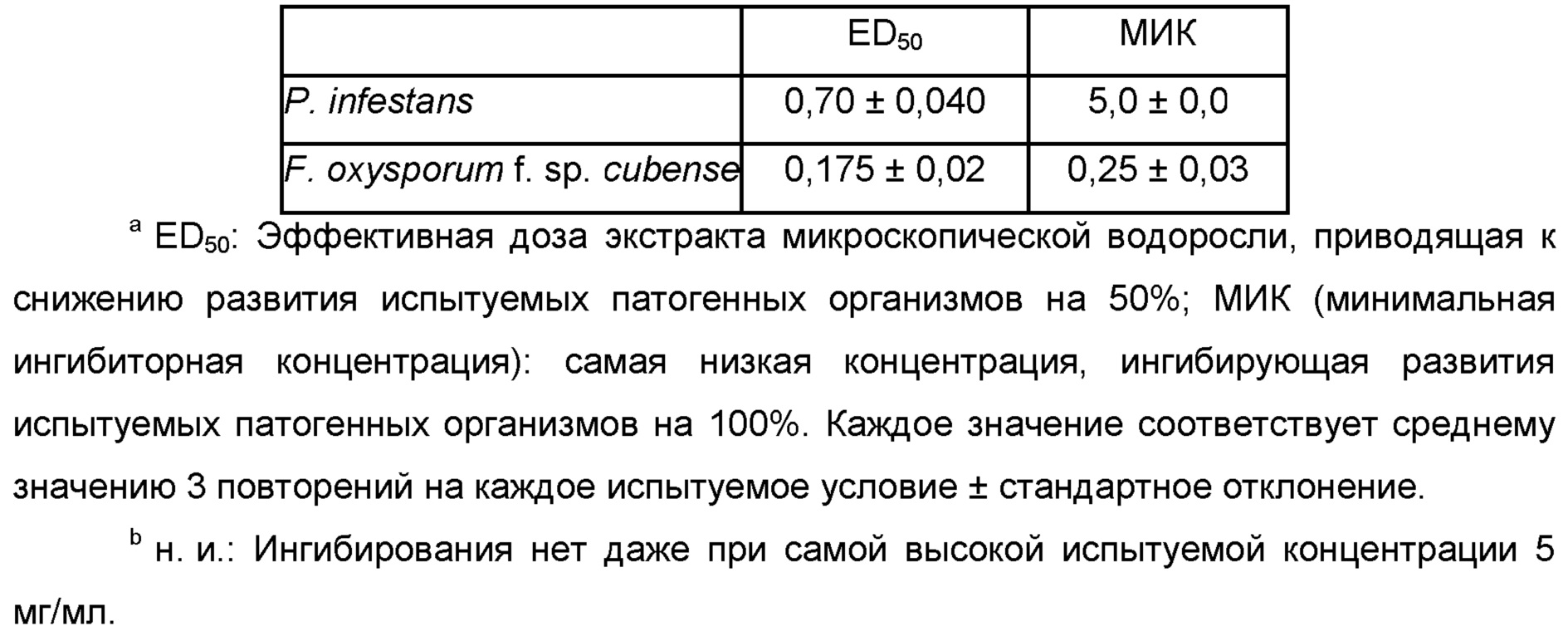
Пример 9: Испытание in vitro и in planta (саженцы яблони) против парши яблок (Venturia inaequalis)
Испытание противогрибковой активности экстракта D в концентрации 5 г/л проводили на чашке Петри в присутствии спор Venturia inaequalis, которые высевали через 2 часа после или за 3 часа до нанесения экстракта D. После 48 часов инкубации оценивали долю прорастания спор в процентах. Присутствие экстракта D за 2 часа до или через 3 часа после инокуляции полностью ингибирует прорастание спор V. inaequalis в отличие от присутствия воды.
Испытания in planta на яблоневых саженцах проводили в соответствии с описанным ниже протоколом:
Испытание проводили в 2 режимах: опрыскивание экстрактом D в концентрации 5 г/л за 2 часа до инокуляции («экстракт до») и опрыскивание экстрактом D в концентрации 5 г/л через 3 часа после инокуляции («экстракт после»).
3 контрольных режима:
2 режима необработанного контроля (опрыскивание водой в то же время, что и препаратом) («вода до» и «вода после»).
1 режим с фунгицидом сравнения (каптаном).
Искусственная инокуляция растений штаммом Venturia inaequalis в количестве 100000 спор/мл.
Симптомы парши отмечали через 4 дня после инокуляции (визуальная оценка % площади, пораженной паршой).
Биологический материал:
Саженцы, полученные из сеянцев яблони приблизительно 3-недельного возраста в почвенных комках - из почек, взятых от сорта Гала (яблони выращены в саду, засаженном сортами Гала и Элстар).
Споры Venturia inaequalis штамма EUB04 на целлофане.
3 ящика по 12 саженцев на каждый режим.
D+9, D+14, D+17, D+21: Запись доли пораженной площади листа в процентах (от 0 до 100% с прибавлениями по 10%).
Поскольку условия нормальности и однородности дисперсий остатков не были выполнены, применяли непараметрический критерий Краскела-Уоллиса. Если этот критерий был статистически значимым, проводили парные сравнения средних значений при различных режимах, используя критерий Неменьи, основанный на расстоянии Тьюки. Эти анализы проводили с помощью программного обеспечения R (версия 3.1.2).
Нанесение экстракта примера 2 не приводило к фитотоксичности для растений: некроза, хлороза или реакций образования пузырей не наблюдалось.
Наблюдения в день (D) 9: симптомы парши отсутствовали при 5 режимах.
На необработанных контрольных растениях (Вода до и ода после) развивалась парша, в среднем достигающая от 50 до 60% в D21. Значимые различия между двумя режимами обработки водой отсутствовали независимо от даты.
На контрольных растениях, обработанных каптаном, развития парши не наблюдалось.
В режимах обработки («экстракт до» и «экстракт после») развитие парши было значительно меньше, чем на необработанных контрольных растениях, независимо от даты наблюдения. Независимо от даты наблюдения значимые различия между режимом «экстракт до» и режимом обработки каптаном отсутствовали. Через 14 дней после инокуляции значимые различия между режимом «экстракт до», режимом «экстракт после» и режимом обработки каптаном отсутствовали, но через 17 и 21 дней после инокуляции парши было значительно больше для режима «экстракт после», чем для режима обработки каптаном. Значимые различия между режимами «экстракт до» и «экстракт после» отсутствовали.
Очень хорошая эффективность экстракта A. carterae против парши отмечена при нанесении за 2 часа до инокуляции или через 3 часа после инокуляции.
Пример 10: Испытание in planta для различных видов Colletotrichum
Чтобы определить, обладает ли экстракт A. carterae биоцидным действием на аскомицет Colletotrichum, испытанию подвергали два различных вида Colletotrichum: Colletotrichum fructicola и Colletotrichum orbiculare. Для этих двух видов процедура была идентичной: некоторый объем экстракта в определенной концентрации (от 1 до 10 г/л) или некоторый объем воды смешивали с некоторым объемом, содержащим 2⋅106 спор Colletotrichum fructicola или 2⋅106 спор Colletotrichum orbiculare, затем наносили 10 мкл спор на отсоединенные листья земляники или огурца соответственно. Листья выдерживали в чашке Петри во влажной атмосфере в течение 6 дней при 24°С в камере для культур клеток (день/ночь; 14 ч/12 ч). Результаты показывают, что экстракт A. carterae полностью ингибирует рост этих двух видов Colletotrichum, начиная с концентрации 1 г/л.
Пример 11: Испытание на целостность клеточной стенки и плазматической мембраны конидиоспор Fusarium graminearum
Конидиоспоры F. graminearum инкубируют в течение 1 часа в присутствии экстракта в соответствии с примером 2 (5 г/л) или такого же экстракта (5 г/л), инактивированного температурой 60°С. Для определения целостности мембраны спор добавляли пропидиумйодид.
На фотографиях показано окрашивание конидиоспор пропидиумйодидом в присутствии экстракта в соответствии с примером 2 (5 г/л) и отсутствие окрашивания в присутствии такого же инактивированного экстракта (5 г/л).
Данные результаты показывают, что под действием экстракта в соответствии с примером 2 пористость клеточной стенки и плазматической мембраны конидиоспор Fusarium graminearum значительно увеличивается.
Все эти результаты показывают, что экстракт, полученный из A. carterae, обладает существенной противогрибковой активностью в отношении широкого спектра фитопатогенных грибов как в условиях in vitro, так и в условиях in planta.
Пример 12: Фракционирование экстракта D методом ВЭЖХ и испытание различных полученных фракций на противогрибковую активность
При попытке охарактеризовать, какие молекулы, присутствующие в экстракте A. carterae, ответственны за противогрибковую активность, была выбрана стратегия биологически управляемого фракционирования: экстракт A. carterae фракционируют на колонке для высокоэффективной жидкостной хроматографии (ВЭЖХ) и проводят испытание на биоцидную активность против спор F. graminearum для каждой фракции, чтобы определить, какая из фракций содержит молекулу(-ы), ответственную(-ые) за противогрибковую активность.
200 мг лиофилизированных клеток солюбилизируют в 1 мл метанола. После центрифугирования в течение 10 мин при 4400 об/мин супернатант выделяют, и суммарную выделенную спиртовую жидкую фазу фильтруют через бумагу, затем раствор выпаривают на роторном испарителе при низкой температуре; таким образом, собирают 48,5 мг экстрагированного продукта. Для подтверждения активности экстракта A. carterae испытание активности проводят для спор F. graminearum (Фиг. 4А).
При наличии активности экстракта проводили эксперименты по полупрепаративному фракционированию на колонке С18 с обращенной фазой, используя высокоэффективный жидкостный хроматограф Thermo Scientific Ultimate 3000, в соответствии с описанным ниже протоколом:
Экстракт A. carterae растворяли до концентрации 5 г/л в метаноле. Для экстракта применяли следующие условия:
Скорость потока: 2,5 мл/мин.
Колонка: С18 с обращенной фазой: L = 250 мм; внутренний диаметр = 10 мм; диаметр пор = 5 мкм.
Объем ввода пробы: 150 мкл.
Температура ввода пробы: 24°С.
Длина волны детектирования: 280 нм.
Программа градиента описана в таблице 7 ниже.

Хроматограмма, полученная в этих условиях, представлена на Фиг. 4В. Как описано на Фиг. 4В, образовалось пять фракций F1-F5, так, что каждый основной пик соответствует фракции. Испытание действия на рост F. graminearum проводили для 5 мг/мл каждой фракции. Эти результаты указывают на то, что только во фракции F1 сохраняется биоцидная активность (Фиг. 4С); для этой фракции была определена МИК 0,75 мг/мл (Фиг. 4D).
Фракцию F1 подвергали новому фракционированию в соответствии со следующим протоколом:
Экстракт F1 растворяли до концентрации 5 г/л в метаноле. Для экстракта D применяли следующие условия:
Скорость потока: 2,5 мл/мин.
Колонка: С18 с обращенной фазой: L = 250 мм; внутренний диаметр = 10 мм; диаметр пор = 5 мкм.
Объем ввода пробы: 150 мкл.
Температура ввода пробы: 24°С.
Длина волны детектирования: = 280 мкм.
Программа градиента описана в таблице 8 ниже.


Хроматограмма, полученная в этих условиях, представлена на Фиг. 5А. Как описано на Фиг. 5А, образовалось пять фракций F1-1-F1-5, при этом основной пик соответствовал фракции F1-2. Определение активности в отношении роста F. graminearum проводили для 5 мг/мл каждой фракции. Эти результаты указывают на то, что только фракции F1-2 и F1-3 обладают биоцидной активностью (Фиг. 5В); для фракции F1-2 была определена МИК 0,75 мг/мл (Фиг. 5С).
Пример 13: Анализ фракции F1-2 методом масс-спектрометрии.
Чтобы лучше охарактеризовать молекулу, присутствующую во фракции F1-2, был проведен масс-спектрометрический анализ в описанных ниже условиях:
Эксперименты проводили в режиме нагнетания на спектрометре QStar Elite (Applied Biosystems).
Режим ионизации: Электрораспыление (ИЭР) в режиме регистрации положительных ионов.
Напряжение пробоя игольчатого разрядника при электрораспылении: 4500 В при комнатной температуре.
Условия ввода пробы: 20 мкл образца растворяли в метаноле при скорости потока метанола 400 мкл/мин.
Диапазон сканирования: 100-2000 Дальтон.
На Фиг. 6А показан масс-спектр, полученный при ионизации электрораспылением в режиме регистрации положительных ионов для интересующей молекулы из собранной фракции F1-2.
Точная масса определена как 1381,8276 Дальтон. Она соответствует аддукту натрия, образующемуся в процессе ионизации ([M+Na]+). После вычитания массы натрия 23 Дальтон масса молекулярного пика соединения составляет 1358,8 Да.
Тщательный анализ точной массы дает возможность определить одну или более эмпирических формул, соответствующих этой массе, с ошибкой 5 частей на миллион (ч.н.м.). После отклонения нереалистических предположений выбранная эмпирическая формула представляет собой C71H122O24.
Для этого пика 1381,8276 Да был проведен тщательный анализ методом тандемной масс-спектрометрии. Масс-спектр представлен на Фиг. 6В. Несколько пиков (круговые пики - Фиг. 6В) аналогичны пикам, полученным в результате фрагментации амфидинола 18 (7).
Пример 14: Анализ фракции F1-2 методом ядерного магнитного резонанса (ЯМР)
Чтобы определить структуру молекулы с молекулярной массой 1358,8 Да, был проведен анализ ЯМР в соответствии с описанной ниже процедурой:
Образцы, соответствующие пику F1-2, собирали, а затем полностью растворяли в 350 мкл дейтерированного метанола (MeOD4).
Эксперименты проводили на спектрометре Bruker Avance 14.1Т, оборудованном многоядерным датчиком. Одно- и двухмерные протонные и углеродные спектры регистрировали, используя последовательности импульсов, имеющиеся в библиотеке последовательностей Bruker.
Условия регистрации описаны ниже:
-1Н: Число сканирований: 512; Импульс: 8 мкс; Регистрация: 5,0 с; Релаксация: 1,0 с.
-13С DEPT135: Число сканирований: 44666; Импульс: 12 мкс; Регистрация: 1,0 с; Релаксация: 3,0 с.
- HSQC: Число сканирований: 64; 512 шагов.
- НМВС: Число сканирований: 48; 512 шагов.
- COSY: Число сканирований: 56; 256 шагов.
- TOCSY: Число сканирований: 48; 256 шагов.
Последовательность13С DEPT135 представляет собой эксперимент по сортировке атомов углерода в соответствии с числом протонов, связанных простой связью: СН3 и СН>0 и СН2<0.
Последовательность корреляционной спектроскопии (COSY) представляет собой двухмерный 2D гомоядерный эксперимент по идентификации протонов в скалярном взаимодействии, разделенных расстоянием в 2 или 3 связи.
Последовательность полной корреляционной спектроскопии (TOCSY) представляет собой двухмерный 2D гомоядерный эксперимент по идентификации протонов в скалярном взаимодействии, разделенных расстоянием в 3 или более связей.
Последовательность гетероядерной одноквантовой корреляции (HSQC) представляет собой 2D гетероядерный эксперимент, который показывает прямые взаимодействия между атомом углерода и протоном(-ами), непосредственно связанном(-ыми) с ним.
Последовательность гетероядерной многосвязной корреляции (НМВС) представляет собой 2D гетероядерный эксперимент, который показывает корреляции между протонами и атомами углерода, разделенными интервалами в 2 или 3 связи.
Спектры 1D обрабатывали с помощью Фурье-преобразования. Спектры обрабатывали с помощью Фурье-преобразования в обоих измерениях.
На основании полученных результатов на зарегистрированном спектре1Н, показанном на Фиг. 7А, показана серия пиков, распределенных в широком спектральном окне, составляющем от 1 до 6 ч. н.м. Это подтверждает, что исследуемое соединение содержит алифатические и олефиновые протоны. Эти сдвиги также позволяют предположить присутствие гетероатомов, таких как атомы кислорода.
На спектре13С DEPT135 (Фиг. 7В) показана серия пиков в спектральном окне, составляющем от 13 до 211 ч.н.м. Последовательности 2D НМВС и HSQC показывают, как связаны друг с другом атомы углерода в цепи. Последовательности 2D COSY и TOCSY подтверждают структурную последовательность на основании протонных сдвигов и корреляций.
Наблюдаемое количество ненасыщенных связей и колец согласуется с расчетным значением для молекулы на основании ее эмпирической формулы (=11):
Ni=(2nС+2-nH+nN-nX)/2,
где: nC: число атомов углерода, nH: число атомов водорода, nN: число атомов азота, nX: число атомов галогена.
11 ненасыщенных связей распределены следующим образом:
- 1 кетоновая функциональная группа;
- 8 двойных связей, включая 2 концевых связи;
- 2 кольца.
Последовательности COSY и TOCSY дают возможность реконструировать весь углеродный скелет структуры. Эти корреляции указаны на Фиг. 8 как связи полужирным шрифтом. Эти результаты показывают, что вторичная структура молекулы соответствует структуре амфидинола 18. Общая интерпретация была подтверждена сравнением результатов, полученных в этом исследовании, с результатами, найденными в литературе (7).
В заключение, все результаты, полученные методами ядерного магнитного резонанса и масс-спектрометрии, подтверждают, что молекула, присутствующая во фракции F1-2, полученной после фракционирования экстракта D и обладающей противогрибковой активностью против F. graminearum, представляет собой молекулу амфидинола 18, конкретные химические данные которой описаны ниже:
Эмпирическая формула: C71H122O24
Молярная масса: 1358,83 г. моль-1
СПИСОК ЛИТЕРАТУРЫ
(1) Arseniuk, Е., Foremska, Е., Goral, Т., Chelkowski, J. 1999. Fusarium head blight reactions and accumulation ofdeoxynivalenol (DON) and some of its derivatives in kernels of wheat, triticale and rye. Journal of Phytopathology 147, 577-590.
(2) Devi P, Wahidulla S, Kamat T and D'Souza L (2011). Screening marine organisms for antimicrobial activity against clinical pathogens. Indian J.Geomar.Sci. 40; 338-346.
(3) Mayer AM, Rodriguez AD, Taglialatela-Scafati O, Fusetani N (2013). Marine pharmacology in 2009-2011: marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar Drugs. 11(7): 2510-73.
(4) Bowler, C, Vardi, A., & Allen, A. E. (2010). Oceanographic and biogeochemical insights from diatom genomes. Annual Review of Marine Science, 2, 333-65. doi: 10.1146/annurev-marine-120308-081051.
(5) Murray S, Garby T, Hoppenrath M, Neilan BA (2012). Genetic diversity, morphological uniformity and polyketide production in dinoflagellates (Amphidinium, Dinoflagellata). PLoS One. 7(6).
(6) Morsy N, Houdai T, Matsuoka S, Matsumori N, Adachi S, et al. (2006). Structures of new amphidinols with truncated polyhydroxyl chain and their membrane-permeabilizing activities. Bioorganic and Medicinal Chemistry 14: 6548-6554.
(7) Nuzzo G, Cutignano A, Sardo A, Fontana A (2014). Antifungal Amphidinol 18 and its 7-sulfate derivative from marine dinoflagellate Amphidinium carterae, J. Nat. Prod., 1524-1527.
Реферат
Группа изобретений относится к биотехнологии и может быть использована в сельском хозяйстве. Способ борьбы с патогенными грибами, оомицетами и/или бактериями сельскохозяйственных культур и/или семян предусматривает получение клеточного экстракта одной или более микроскопических водорослей рода Amphidinium путем сбора свежих клеток одной или более микроскопической(-их) водоросли(-ей)рода Amphidinium при концентрации клеток, составляющей от 5⋅104 клеток/мл до 5⋅106 клеток/мл, необязательного замораживания и/или лиофилизации указанных клеток и ресуспендирования указанного лиофилизата или указанных свежих или замороженных клеток в неорганическом или органическом растворителе в массовом соотношении от 1:200 до 1:50. Полученный клеточный экстракт микроскопических водорослей рода Amphidinium содержит амфидинол 18 или амфидинол 19, предпочтительно амфидинол 18, и его наносят на сельскохозяйственные культуры и/или покрывают им указанные семена. При этом микроскопические водоросли рода Amphidinium представляет собой Amphidinium carterae. Группа изобретений позволяет повысить урожайность сельскохозяйственных растений. 8 з.п. ф-лы, 10 ил, 8 табл., 14 пр.
Комментарии