Способы борьбы с вредителем - совкой agrotis - RU2311767C2
Код документа: RU2311767C2
Чертежи



















Описание
Перекрестная ссылка на близкие заявки
Данная заявка претендует на приоритет со времени подачи предварительной заявки на патент США с порядковым № 60/150319, поданной 23 августа 1999 г.
Предпосылка изобретения
Насекомые и другие вредители обходятся фермерам в миллиарды долларов ежегодно из-за потери урожая и затрат на борьбу с этими вредителями. Потери, причиняемые насекомыми-вредителями в условиях производства сельскохозяйственной продукции, включают снижение урожайности и снижение качества урожая. Повышенная стоимость уборки урожая также является результатом заражений насекомыми.
Частично проблемы с насекомыми принимались решать способами культивирования, такими как севооборот, или внесением удобрений с высокими уровнями фосфата, чтобы стимулировать рост сильных корневых систем у растений. Однако севооборот может быть нарушен, например, возникающей у северной длинноусой блошки особенностью - двухлетней диапаузой (или перезимовкой). Больше всего надеются на химические инсектициды, чтобы гарантировать желательный уровень контроля.
Химические пестициды обеспечили эффективный способ контроля вредителей. Однако общественность стала беспокоиться о количестве остаточных химических соединений, которые могут быть обнаружены в пище, грунтовой воде и других местах в окружающей среде. Поэтому синтетические химические пестициды все больше и больше тщательно исследуются в отношении их потенциальных неблагоприятных последствий для окружающей среды. Некоторые синтетические химические пестициды могут отравлять почву и нижележащие водоносные пласты, загрязнять поверхностные воды в результате стока и уничтожать нецелевые формы жизни. Некоторые синтетические средства химической борьбы имеют дополнительный недостаток, представляющий угрозу для безопасности общества, когда они применяются в районах, где с ними могут контактировать домашние животные, сельскохозяйственные животные или дети. Они также могут представлять опасность для здоровья людей, которые их применяют, особенно, если не следуют надлежащим техническим приемам применения. Регулирующие ведомства во всем мире ограничивают и/или запрещают применение многих синтетических пестицидов, особенно тех, которые являются стойкими в окружающей среде и которые вступают в пищевую цепь. Жесткие новые ограничения на применение пестицидов и исключение некоторых эффективных пестицидов из мест сбыта могут ограничить экономичные и эффективные варианты борьбы со значимыми вредителями.
Из-за проблем, связанных с применением многих синтетических химических пестицидов, существует очевидная необходимость ограничить применение этих агентов и идентифицировать альтернативные средства борьбы. Замена синтетических химических пестицидов или комбинация этих средств с биологическими пестицидами может снизить уровни токсичных химикатов в окружающей среде.
Биологическим пестицидным средством, популярность применения которого возрастает, является почвенный микроорганизм Bacillus thuringiensis (B.t.). Почвенный микроорганизм Bacillus thuringiensis (B.t.) представляет собой грамположительную спорообразующую бактерию. Большинство штаммов B.t. не проявляет пестицидной активности. Некоторые штаммы B.t. продуцируют пестицидные белковые включения в параспорах. Эти белковые включения под микроскопом часто выглядят в виде кристаллов различной формы. Форма и тип кристаллических включений могут использоваться для характеристики штаммов B.t. Присутствующие в кристаллических включениях "δ-эндотоксины", которые обычно обладают специфичной пестицидной активностью, отличаются от экзотоксинов, которые не имеют определенного круга хозяев.
Коммерческое применение пестицидов B.t. изначально ограничено узким диапазоном вредителей - чешуекрылых (гусениц). Например, препараты спор и кристаллов В. thuringiensis подвида kurstaki многие годы используются как коммерческие инсектициды для чешуекрылых вредителей. Однако совсем недавно исследователи открыли пестициды B.t., специфичные для более широкого круга вредителей. Например, B.t. israelensis и morrisoni были использованы в промышленном масштабе для контроля насекомых отрядов Diptera и Coleoptera соответственно (Gaertner, F.H. [1989] "Cellular Delivery Systems for Insecticidal Proteins: Living and Non-Living Microorganisms" in Controlled Delivery of Crop Protection Agents, R.M.Wilkins, ed., Taylor and Francis, New York and London, 1990, pp.245-255).
В настоящее время идентифицированы новые подвиды B.t. и выделены и секвенированы гены, кодирующие активные белки δ-эндотоксинов (Höfte, H., H.R.Whiteley [1989] Microbiological Reviews 52 (2): 242-255). Höfte и Whiteley сгруппировали гены кристаллических белков B.t. в четыре больших класса. Классы представляли собой cryI (специфичный для Lepidoptera), cryII (специфичный для Lepidoptera и Diptera), cryIII (специфичный для Coleoptera) и cryIV (специфичный для Diptera). Сообщалось об открытии штаммов со специфичной токсичностью для других вредителей (Feitelson, J.S., J.Payne, L.Kim [1992] Bio/Technology 10: 271-275). Например, обозначения CryV и CryVI были предложены для двух новых групп токсинов, активных против нематод.
Многие белки δ-эндотоксинов Bacillus thuringiensis составлены из двух функциональных сегментов. Данные "полноразмерные" белки построены из устойчивого к протеазам корового токсина, который примерно соответствует первой половине (N-концевой части) белковой молекулы. Известна трехмерная структура корового сегмента CryIIIA δ-эндотоксина B.t. и было сделано предположение, что все родственные токсины имеют такую же полную структуру (Li, J., J.Carroll, D.J.Ellar [1991] Nature 353: 815-821). Вторая половина (С-концевая часть) молекулы часто называется "сегментом протоксина". Предполагают, что сегмент протоксина принимает участие в образовании кристаллов токсина (Arvidson, Н., Р.Е.Dunn, S.Strand, A.I.Aronson [1989] Molecular Microbiology 3: 1533-1534; Choma С.Т., W.K.Surewicz, P.R.Carey, M.Pozsgay, T.Raynor, H.Kaplan [1990] Eur. J. Biochem. 189: 523-527). В кишке насекомых полная молекула токсина с М.м. 130 кД обычно процессируется протеазами до устойчивого корового сегмента. Таким образом, сегмент протоксина может придавать токсину частичную специфичность к насекомым за счет ограничения доступности кора для насекомых путем снижения процессинга молекулы токсина протеазами (Haider, M.Z., B.H.Knowles, D.J.Ellar [1986] Eur. J. Biochem. 156: 531-540) или путем снижения растворимости токсина (Aronson, A.I., E.S.Han, W.McGaughey, D.Johnson [1991] Appl. Environ. Microbiol. 57: 981-986).
Номенклатура и схема классификации 1989 г. Höfte и Whiteley была основана как на рассчитанной аминокислотной последовательности, так и на круге хозяев для токсина. Эта система была адаптирована для того, чтобы охватить 14 различных типов генов токсинов, которые были разделены на пять больших классов. Была предложена видоизмененная схема номенклатуры, которая основана только на идентичности аминокислот (Crickmore et al. [1996] Society for Invertebrate Pathology, 29th Annual Meeting, IIIrd International Colloquium on Bacillus thuringiensis, University of Cordoba, Cordoba, Spain, September 1-6, 1996, abstract). Мнемонический термин "cry" был сохранен для всех генов токсина за исключением cytA и cytB, которые находятся в отдельном классе. Латинские цифры были заменены арабскими цифрами в первичной категории и круглые скобки в третичной категории были удалены. В настоящее время границы отображают 95% (третичная категория), 75% (вторичная категория) и 48% (первичная категория) идентичности последовательностей. Многие из исходных названий были сохранены, хотя некоторое количество было классифицировано повторно. Смотри также "Revisions of the Nomenclature for the Bacillus thuringiensis Pesticidal Crystal Proteins", N.Crickmore, D.R.Zeigler, J.Feitelson, E.Schnepf, J.Van Rie, D.Lereclus, J.Baum and D.H.Dean, Microbiology and Molecular Biology Reviews (1988) Vol.62: 807-813 и Crickmore, Zeigler, Feitelson, Schnepf, Van Rie, Lereclus, Baum and Dean, "Bacillus thuringiensis toxin nomenclature" (1999) http://www.biols.susx.ac.uk/Home/Neil_Crickmore/Bt/index.html. Данная система использует легкодоступные приложения для компьютерных программ CLUSTAL W и PHYLIP. Приложение NEIGHBOR в пакете PHYLIP использует алгоритм средних арифметических (UPGMA).
Выделение новых изолятов B.t. и генов их токсинов, а также определение точного диапазона пестицидного действия токсинов было медленным эмпирическим процессом. В результате интенсивного исследования и вложения инвестиций выданы патенты, посвященные новым изолятам, токсинам и генам B.t. и новым применениям известных токсинов и изолятов B.t. Смотри для обзора Feitelson et al. выше. Однако открытие новых изолятов B.t. и новых применений известных изолятов и токсинов B.t. остается эмпирической непредсказуемой областью.
Smulevitch et al. ([1991] FEBS Lett. 293: 25-26), Gleave et al. ([1991] JGM 138: 55-62) и Shevelev et al. ([1993] FEBS Lett. 336: 79-82) описали характеристику Cry9 токсинов, активных против чешуекрылых. Lambert et al. (Lambert, В., L.Buysse, С.Decock, S.Jansens, C.Piens, В.Saey, J.Seurinck, K. van Audenhove, J.Van Rie, A.Van Vliet, M.Peferoen [1996] Appl. Environ. Microbiol. 62 (1): 80-86) и опубликованные РСТ заявки WO 94/05771 и WO 94/24264 также описывают изоляты B.t. и Cry9С токсины, активные против чешуекрылых вредителей.
В патентах США №5126133; 5188960; 5246852 и 5691308 заявлены Cry1Fa токсин и ген (811A) изолята PS81I B.t. Патенты США №5527883; 5508264; 5827514 и 5840554 имеют отношение к химерным токсинам CryIF. В WO 99/24581 заявлены различные оптимизированные для растений гены cry1F. В патенте США №5686069 заявлены токсин и ген Cry1Fb изолята PS91C2 B.t.
С применением генно-инженерных методов разрабатываются различные подходы для применения токсинов B.t. в сельском хозяйстве. В литературе, опубликованной более 15 лет назад, было описано клонирование и экспрессия гена кристаллического белка B.t. в Escherichia coli (Schnepf, H.E., H.R.Whiteley [1981] Proc. Natl. Acad. Sci. USA 78: 2893-2897). В обоих патентах США №4448885 и 4467036 заявлена экспрессия кристаллического белка B.t. в Е. coli. Были получены продукты B.t., основанные на рекомбинантной ДНК, и одобрены для применения, включая применение растений, модифицированных генами B.t. устойчивости к насекомым, и применение стабильных клеток микроорганизмов в качестве средств доставки белков B.t. Были разработаны различные усовершенствования путем модификации токсинов B.t. и/или их генов. Например, патенты США №5380831 и 5567862 относятся к получению синтетических генов инсектицидных кристаллических белков с повышенным уровнем экспрессии в растениях. Таким образом, выделенные гены эндотоксинов B.t. становятся коммерчески доступными.
Препятствия для успешного применения токсинов B.t. в сельском хозяйстве включают развитие у насекомых устойчивости к токсинам B.t. Кроме того, некоторые насекомые могут быть невосприимчивы к действиям B.t. К последним относятся такие насекомые, как долгоносики и совки, для которых ранее было показано отсутствие видимой чувствительности к большинству δ-эндотоксинов B.t.
Совки в развитии проходят несколько возрастных стадий, как, например, стадию личинки. Хотя подрезание проростков личинкой на поздней стадии является причиной наиболее широко распространенного повреждения и экономических потерь, поедание листьев обычно приводит к потере урожая таких культур, как кукуруза. При достижении поздних возрастных стадий (например, от третьей до четвертой) личинка начинает подрезать растения и части растений, особенно проростки. Из-за изменений в режиме питания популяции, вызывающие экономически значимые повреждения, могут возникать неожиданно при небольшом количестве ранних предвестников. Крупные совки могут уничтожить несколько проростков в день, и сильное заражение может полностью истребить все насаждения культур. Ночной образ жизни и поведение, связанное с зарыванием в почву, также делает обнаружение и обработку проблематичными.
Совка ипсилон (Agrotis ipsilon (Hufnagel); Lepidoptera: Noctuidae) является серьезным вредителем многих культур, включая кукурузу, хлопок, культуры капусты (Brassica, брокколи, капуста, китайская капуста) и дерновые. Дополнительные хозяйские растения включают свекловицу, Capsicum (перцы), нут, бобы, салат-латук, люцерну, лук, картофель, редьку, рапс (канолу), рис, сою, землянику, сахарную свеклу, табак, томаты и лесные деревья. В Северной Америке вредители рода Agrotis поедают клевер, зерновые, табак, коноплю, лук, землянику, ежевику, малину, люцерну, ячмень, бобы, капусту, овес, горох, картофель, батат, томат, садовые цветы, травяные культуры, люцерну посевную, кукурузу, спаржу, виноград, почти все виды листьев, сорняки и многие другие культуры и садовые растения. Вредителями являются другие совки трибы Agrotini, в частности совки рода Feltia (например, F. jaculifera (Guenée); эквивалент ducens subgothica) и Euxoa (например, Е. messoria (Harris), E. scandens (Riley), E. auxiliaris Smith, E. detersa (Walker), E. tessellata (Harris), E. ochrogaster (Guenée). Такие совки, как Peridroma saucia, также могут приносить значительный вред. Цитрусовые растения, например, также могут быть мишенью для нападения совок (примером является "цитрусовая совка" Xylomyges curialis).
Совки также являются вредителями и за пределами Северной Америки и более значимые для экономики вредители нападают на нут, пшеницу, овощи, сахарную свеклу, люцерну, кукурузу, картофель, турнепс, рапс, салат-латук, землянику, логанову ягоду, лен, хлопок, сою, табак, свекловицу, китайскую капусту, томаты, баклажаны, сахарный тростник, пастбищные растения, капусту, арахис, Cucurbita, турнепс, подсолнечник, Brassica, лук, лук-порей, сельдерей, кунжут, спаржу, ревень, цикорий, тепличные культуры и шпинат. Совка ипсилон A. ipsilon встречается как вредитель за пределами Северной Америки, включая Центральную Америку, Европу, Азию, Австралию, Африку, Индию, Тайвань, Мексику, Египет и Новую Зеландию.
Основными видами совок в Аргентине являются Agrotis malefida, Porosagrotis gypaetiana и Agrotis ipsilon (также имеющей написание ypsilon). Эти совки нападают, например, на культурные сорта зерновых, сои и подсолнечника. Данные насекомые могут серьезно уменьшить популяцию проростков и в случае мощных атак могут полностью уничтожить целые делянки.
Контроль культур для A. ipsilon, такой как контроль окружающих сорняков, может помочь предотвратить тяжелые заражения; однако такие способы не всегда осуществимы или эффективны. Заражения очень случайны и в прошлом применение инсектицидов перед посадкой или при посадке не было эффективным. Некоторые приманки пригодны для контроля совок в посевах. Чтобы защитить дерновые травы, такие как полевицу побегообразующую, применялись химические инсектициды. Применение химических пестицидов на покрытых дерном площадях вызывает особую озабоченность (например, площадки для игры в гольф, стадионы, парки и другие рекреационные зоны, профессиональная ландшафтная архитектура и домашние газоны) из-за близкого контакта людей с этими площадями. Природные продукты (например, нематоды и азадирахтин) обычно производятся недостаточно.
До настоящего времени продукты Bacillus thuringiensis для контроля совки ипсилон широко не использовались из-за того, что раньше они были не достаточно эффективны. Высокая скорость питания совки и поведение, связанное с зарыванием в почву, делает их контроль имеющимися в настоящее время пестицидными растворами, основанными на биологических средствах, чрезвычайно трудным. Например, Cry1A(b) токсины в основном не эффективны против совок.
Сущность изобретения
Заявленное изобретение относится к удивительному открытию того, что белки Cry1F активны против таких совок, как совка ипсилон (Agrotis ipsilon). Таким образом, заявленное изобретение предоставляет способы контролирования этих вредителей, где указанный способ включает в себя осуществление контакта указанного вредителя с оказывающим пестицидное действие количеством токсина Bacillus thuringiensis, содержащего, по меньшей мере, пестицидную часть Cry1F токсина. Поэтому в заявленное изобретение включены полноразмерные, укороченные или химерные Cry1F белки и гены. В предпочтительных вариантах Cry1F токсином является токсин Cry1Fa. Согласно заявленному изобретению могут быть использованы белки Cry1F дикого типа и синтетические Cry1F белки. Таким образом, в данное изобретение включено применение полинуклеотидов (и/или их комплементов, предпочтительно их полных комплементов), которые гибридизуются с известными cry1F генами, предпочтительно с частями, кодирующими кор токсина. В предпочтительных вариантах используются полинуклеотиды, оптимизированные для растений.
Заявленное изобретение включает применение трансгенных хозяев, включая такие хозяйские растения, как зерновые, хлопок и подсолнечник. В предпочтительном варианте заявленное изобретение относится к клеткам растений, трансформированным, по меньшей мере, одной полинуклеотидной последовательностью заявленного изобретения так, что трансформированные растительные клетки экспрессируют пестицидные белки в тканях, уничтожаемых вредителями-мишенями. Такая трансформация растений может быть достигнута с использованием технологий, хорошо известных специалистам в данной области, и в типичном случае может вовлекать модификацию гена, чтобы оптимизировать экспрессию токсина в растениях.
Краткое описание рисунков
На фигуре 1 показана масса интактных проростков пшеницы и относительная оценка повреждений через 24 часа после заражения личинками совки ипсилон 4ой возрастной стадии и после различных обработок.
На фигуре 2 показан сырой вес проростков пшеницы, зараженных личинками совки ипсилон 3ей возрастной стадии через 48 часов после заражения и после различных обработок.
Фигуры 3 и 4 иллюстрируют результаты, обсуждаемые в примере 4.
Краткое описание последовательностей
SEQ ID NO. 1 является полинуклеотидной последовательностью полноразмерного оптимизированного для растений гибридного гена cryIF/CryIA(b), названного 1F1AB-PO.
SEQ ID NO. 2 является аминокислотной последовательностью полноразмерного оптимизированного для растений химерного токсина CryIF/CryIA(b). Данный токсин кодируется геном 1F1AB-PO.
SEQ ID NO. 3 является полинуклеотидной последовательностью укороченного оптимизированного для растений гена cryIF, названного 1F-T-PO.
SEQ ID NO. 4 является аминокислотной последовательностью укороченного оптимизированного для растений токсина CryIF. Данный токсин кодируют гены, названные 1F-T-PO, 1F-7G-PO и 1F-7Z-PO.
SEQ ID NO. 5 является полинуклеотидной последовательностью полноразмерного гена токсина B.t. дикого типа, названного 81IA (cryIFa).
SEQ ID NO. 6 является аминокислотной последовательностью полноразмерного токсина B.t. дикого типа, названного 81IA (CryIFa).
SEQ ID NO. 7 является полинуклеотидной последовательностью гена, названного 1F-7G-PO, который оптимизирован для экспрессии в хлопке.
SEQ ID NO. 8 является полинуклеотидной последовательностью гена, названного 1F-7Z-PO, который оптимизирован для экспрессии в кукурузе.
Подробное описание изобретения
Заявленное изобретение относится к удивительному открытию, что Cry1F белки активны против таких совок, как совка ипсилон (Agrotis ipsilon). Особенно удивительными являются данные о том, что с личинками 3ей возрастной стадии и выше можно бороться с помощью токсинов и генов согласно заявленному изобретению. Борьба с личинками совки первой и второй возрастных стадий гораздо менее важна, чем борьба на более поздних стадиях. Хотя было известно, что токсины CryI активны против чешуекрылых в целом, считали, что совки вообще не поддаются обработке токсинами B.t., как обсуждалось выше более подробно в разделе, посвященном предпосылкам.
Заявленное изобретение включает применение рекомбинантных хозяев и предоставляет оптимизированные для растений полинуклеотидные последовательности. Данные полинуклеотидные последовательности включают оптимизированные для растений гены, названные 1F1AB-PO, 1F-T-PO, 1F-7G-PO и 1F-7Z-PO. Данные гены заявлены в WO 99/24581. К предпочтительным растениям-хозяевам относятся зерновые, соя, хлопок, пшеница, канола и подсолнечник.
В некоторых вариантах заявленного изобретения гены кодируют токсин CryIF, который является укороченным по сравнению с полноразмерным токсином CryIF. В укороченных токсинах заявленного изобретения в типичном случае отсутствует весь или часть сегмента протоксина. Также укороченные гены заявленного изобретения могут быть использованы для получения химерных генов и белков. Одним из примеров является оптимизированный для растений ген, содержащий часть cryIF и часть cryIA(b), где гибридный ген кодирует химерный токсин. Предпочтительные химерные гены и токсины заявлены в патенте США №5527883 и 5840554. Другие химерные гены и токсины, которые могут быть использованы согласно заявленному изобретению, заявлены в патентах США №5508264 и 5827514. В предпочтительном варианте часть CryIF химерного токсина сама по себе является пестицидом. Слитые токсины также могут быть использованы согласно заявленному изобретению.
В предпочтительном варианте заявленное изобретение относится к клеткам растений, трансформированным, по меньшей мере, одной полинуклеотидной последовательностью так, что трансформированные растительные клетки экспрессируют пестицидные токсины в тканях, уничтожаемых вредителями-мишенями, которые при этом контактируют с пестицидным белком. Такая трансформация растений может быть достигнута с использованием технологий, хорошо известных специалистам в данной области, и в типичном случае может вовлекать модификацию гена, чтобы оптимизировать экспрессию токсина в растениях.
Специалистам в данной области должно быть очевидно, что когда даны последовательности генов, которые здесь определены, гены заявленного изобретения могут быть получены несколькими способами. В предпочтительных вариантах представленные гены могут быть сконструированы синтетически с применением, например, синтезатора генов. Специфичные гены, приведенные здесь в качестве примеров, также могут быть получены с помощью модифицирования в соответствии с инструкциями заявленного изобретения определенных генов дикого типа (например, технологией точечных мутаций) определенных изолятов, помещенных на хранение в банке культур, как обсуждается ниже. Например, ген дикого типа cryIF может быть получен из изолята B.t. PS81I. Подобным образом части cryIA(b) гибридных генов заявленного изобретения могут быть получены синтетически или могут быть получены в результате модификации генов дикого типа. Токсины и гены CryIA(b), например, описаны в Höfte et al. (1986) Eur. J. Biochem. 161; 273; Geiser et al. (1986) Gene 48: 109 и Haider et al. (1988) Nucleic Acids Res. 16: 10927. Клоны и дополнительные изоляты дикого типа обсуждаются более детально выше - в разделе, названном "Предпосылки изобретения", и в списке ниже.
Культуры, обсуждаемые в данной заявке, были помещены в соответствии с Будапештским соглашением в коллекцию запатентованных культур службы сельскохозяйственных исследований (NRRL), Northern Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA. Депонированные штаммы, приведенные в списке ниже, заявлены в патентных публикациях, как обсуждалось выше, в разделе, названном "Предпосылки изобретения".
Следует понимать, что доступность депозита не дает разрешения для практического использования заявленного изобретения в нарушении патентных прав, охраняемых правительственным документом.
Гены и токсины. Полинуклеотиды заявленного изобретения могут быть использованы для образования полных "генов" для кодирования белков или пептидов в желательной клетке-хозяине. Например, как может легко определить специалист, некоторые полинуклеотиды в прилагаемом списке последовательностей показаны без стоп-кодонов. Также представленные полинуклеотиды могут быть соответствующим образом помещены под контроль промотора в интересующей клетке-хозяине, что легко понятно специалисту в данной области.
Как легко может представить специалист, ДНК обычно существует в двухнитевой форме. В данной структуре одна нить комплементарна другой нити и наоборот. Когда ДНК реплицируется в растении (например), продуцируются дополнительные комплементарные нити ДНК. Таким образом, заявленное изобретение включает применение полинуклеотидов, приведенных в качестве примеров в прилагаемом списке последовательностей, и комплементарных им нитей. Термин "кодирующая нить" часто используется в данной области по отношению к нити, которая связывается с антисмысловой нитью. Для того чтобы экспрессировать белок in vivo, нить ДНК обычно транскрибируется в комплементарную нить мРНК, которая используется как матрица для трансляции белка, мРНК фактически воспроизводит "антисмысловую" нить ДНК. "Смысловая" или "кодирующая" нить имеет ряд кодонов (кодон представляет собой три нуклеотида, которые могут считываться по три одновременно, чтобы получить конкретную аминокислоту), которые могут быть считаны в виде открытой рамки считывания (ORF), чтобы образовать интересующий белок или пептид. РНК и ПНК (нуклеиновая кислота пептида), которые функционально эквивалентны приведенным в качестве примеров ДНК, включены в заявленное изобретение.
Гены и токсины заявленного изобретения могут быть идентифицированы, получены и охарактеризованы с использованием, например, олигонуклеотидных зондов. Зонды представляют собой определяемые нуклеотидные последовательности. Полинуклеотиды заявленного изобретения, приведенные в качестве конкретных примеров, включая их части, которые достаточны для кодирования активного токсина, сами могут быть использованы в качестве зондов. Зондами могут быть ДНК, РНК или ПНК. Эти последовательности могут быть обнаружены в силу включения подходящей метки или могут быть, например, сделаны исходно флюоресцирующими, как описано в международной заявке № WO 93/16094. Как хорошо известно в данной области, если молекула зонда и образец нуклеиновой кислоты гибридизуются с образованием прочной связи между двумя молекулами, этого может быть достаточно, чтобы полагать, что данная проба и образец имеют значительную гомологию. Предпочтительно гибридизация проводится в жестких условиях с использованием технологий, хорошо известных в данной области, как описано, например, в Keller, G.H., M.M.Manak (1987) DNA Probes, Stockton Press, New York, NY., pp.169-170. Например, как установлено в этой работе, условия гибридизации высокой жесткости могут быть достигнуты первой промывкой 2х SSC (стандартный солевой цитратный раствор)/0,1% SDS (додецилсульфат натрия) в течение 15 минут при комнатной температуре. Обычно выполняют две промывки. Высокая жесткость может быть достигнута понижением концентрации соли и/или повышением температуры. Например, после указанного выше этапа гибридизации может следовать промывка 0,1x SSC/0,1% SDS в течение 15 минут при комнатной температуре, за которой в свою очередь следует промывка 0,1x SSC/0,1% SDS в течение 30 минут при 55°С. Температуры, используемые на данных этапах, также могут быть использованы в других обсуждаемых здесь протоколах (где, например, вместо SSC используется SSPE), как может быть известно в данной области. 2x SSC/0,1% SDS может быть получен добавлением 50 мл 20x SSC и 5 мл 10% SDS к 445 мл воды. 20x SSC можно получить в результате объединения NaCl (175,3 г/0,150 М), цитрата натрия (88,2 г/0,015 М) и воды до 1 литра с последующим доведением рН до 7,0 с помощью 10 н. NaOH. 10% SDS можно получить, растворяя 10 г SDS в 50 мл автоклавированной воды разбавлением до 100 мл и разделением на аликвоты.
Регистрация зонда предоставляет средства для выявления известным способом того, произошла ли гибридизация. Такой анализ зондов предоставляет быстрый способ идентификации генов заявленного изобретения, кодирующих токсины. Нуклеотидные фрагменты, которые используются в качестве зондов согласно изобретению, могут быть синтезированы с применением ДНК синтезатора и стандартных процедур.
В используемом здесь смысле термин "жесткие" условия гибридизации относится к условиям, при которых достигается такая же или почти такая же степень специфичности гибридизации, как и в условиях, применяемых данными заявителями. В частности, гибридизацию иммобилизованной ДНК на Саузерн-блотах с 32Р-меченными геноспецифичными зондами выполняли стандартными способами (Maniatis, Т., E.F.Fritsch, J.Sambrook [1982] Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY). В общем, гибридизация и последующие промывки выполняли в жестких условиях, которые позволяли выявлять последовательности мишени. Для двухнитевых генных ДНК зондов гибридизацию выполняли в течение ночи при температуре на 20-25°С ниже температуры плавления (Тm) гибрида ДНК в 6Х SSPE, 5X раствора Денхардта, 0,1% SDS, 0,1 мг/мл денатурированной ДНК. Температура плавления описывается следующей формулой (Beltz, G.A., К.A.Jacobs, Т.Н.Eickbush, P.Т.Cherbas and F.C.Kafatos [1983] Methods of Enzymology, R.Wu, L.Grossman and K.Moldave [eds.] Academic Press, New York 100: 266-285).
Тm=81,5°C + 16,6 Log [Na+] + 0,41 (%G + С) - 0,61 (% формамида) - 600/длина дуплекса в парах нуклеотидов.
Промывки обычно выполняют следующим образом:
(1) Дважды при комнатной температуре в течение 15 минут в 1X SSPE, 0,1% SDS (промывка низкой жесткости).
(2) Однократно при Tm - 20°С в течение 15 минут в 0,2Х SSPE, 0,1% SDS (промывка средней жесткости).
Для олигонуклеотидных зондов гибридизацию выполняли в течение ночи при температуре на 10-20°С ниже температуры плавления (Тm) гибрида в 6Х SSPE, 5X раствора Денхардта, 0,1% SDS, 0,1 мг/мл денатурированной ДНК. Температуру плавления определяли по следующей формуле:
Тm (°С) = 2 (число Т/А пар нуклеотидов) + 4 (число G/C пар нуклеотидов)
(Suggs, S.V., Т.Miyake, E.H.Kawashime, M.J.Johnson, К.Itakura and R.B.Wallace [1981] ICN-UCLA Symp. Dev. Biol. Using Purified Genes, D.D.Brown [ed.]. Academic Press, New York, 23: 683-693).
Промывки обычно выполняют следующим образом:
(1) Дважды при комнатной температуре в течение 15 минут в 1X SSPE, 0,1% SDS (промывка низкой жесткости).
(2) Однократно при температуре гибридизации в течение 15 минут в 1X SSPE, 0,1% SDS (промывка средней жесткости).
Модификация генов и токсинов. Гены и токсины, пригодные согласно заявленному изобретению, включают не только приведенные конкретные примеры последовательностей, но также включают части и/или фрагменты (включая внутренние и/или концевые делеции по сравнению с полноразмерными белками), варианты, мутанты, замещенные белки (белки, имеющие замещенные аминокислоты), химерные и слитые белки, которые сохраняют характерную пестицидную активность белков, приведенных здесь в качестве конкретных примеров. В используемом здесь смысле термины "варианты" или "вариации" генов относятся к нуклеотидным последовательностям, которые кодируют такие же токсины или которые кодируют эквивалентные токсины, обладающие пестицидной активностью. В используемом здесь смысле термин "эквивалентные токсины" относится к токсинам, обладающим такой же или по существу такой же биологической активностью, направленной против вредителей мишеней, как и заявленные токсины.
Гены могут быть модифицированы и варианты генов могут быть легко сконструированы с помощью стандартных технологий. Например, технологии получения точечных мутаций хорошо известны в данной области. Также в патенте США № 5605793 описаны, например, способы создания дополнительного различия молекул с использованием повторной сборки ДНК после случайной фрагментации. Фрагменты полноразмерных генов могут быть получены с применением доступных из коммерческих источников эндонуклеаз и могут быть использованы экзонуклеазы в соответствии со стандартными процедурами. Например, такие ферменты, как Ваl31 или сайт-специфичный мутагенез, могут быть использованы для того, чтобы систематично отрезать нуклеотиды от концов данных генов. Также гены, которые кодируют активные фрагменты, могут быть получены с использованием различных ферментов рестрикции. Чтобы непосредственно получить активные фрагменты данных токсинов, могут быть использованы протеазы.
Эквивалентные токсины и/или гены, кодирующие эти эквивалентные токсины, могут быть получены из изолятов B.t. и/или ДНК библиотек с использованием предоставленных здесь руководств. Существует ряд способов получения пестицидных токсинов настоящего изобретения. Например, обнаруженные и заявленные здесь антитела к пестицидным токсинам могут быть использованы для идентификации и выделения других токсинов из смеси белков. В частности, антитела могут быть выработаны к частям токсинов, которые наиболее константны и наиболее отличаются от других токсинов B.t. Данные антитела могут быть затем использованы, в частности, для того, чтобы идентифицировать эквивалентные токсины с характерной активностью путем иммунопреципитации, ферментным иммуносорбентным анализом (ELISA), или вестерн-блоттингом. Заявленные здесь антитела к токсинам, или эквивалентным токсинам, или фрагментам данных токсинов легко могут быть получены с использованием стандартных в данной области процедур. Гены, которые кодируют данные токсины, могут быть затем получены из микроорганизмов.
Вследствие избыточности генетического кода множество различных последовательностей ДНК могут кодировать одни и те же аминокислотные последовательности. Специалистам в данной области легко создать данные альтернативные ДНК-последовательности, кодирующие те же самые или в значительной степени такие же токсины. Данные варианты последовательностей ДНК находятся в рамках заявленного изобретения. В используемом здесь смысле упоминание "в значительной степени такой же" последовательности относится к последовательностям, которые имеют аминокислотные замены, делеции, присоединения или инсерции, которые существенно не влияют на пестицидную активность. Фрагменты, сохраняющие пестицидную активность, также включены в данное определение.
Эквивалентные токсины будут обладать сходством аминокислот (и/или гомологией) с приведенным в качестве примера токсином. Сходство/идентичность аминокислот обычно будет больше чем 60%, предпочтительно больше чем 75%, более предпочтительно больше чем 80%, еще более предпочтительно больше чем 90% и может быть более 95%. Предпочтительные полинуклеотиды и белки заявленного изобретения также могут быть определены терминами более конкретных пределов идентичности и/или сходства. Например, идентичность и/или сходство может составлять 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% при сравнении с приведенной здесь в качестве примера последовательностью. Если не оговорено особо, используемый здесь процент идентичности и/или сходства последовательностей двух нуклеиновых кислот определяется с использованием алгоритма Karlin и Altschul (1990), Proc. Natl. Acad. Sci. USA 87: 2264-2268, модифицированного как в Karlin and Altschul (1993), Proc. Natl. Acad. Sci. USA 90: 5873-5877. Такой алгоритм включен в программы NBLAST и XBLAST Altschul et al. (1990), J.Mol. Biol. 215: 402-410. BLAST-исследования нуклеотидов выполняли с помощью программы NBLAST, оценка = 100, длина кодовой комбинации = 12, чтобы получить нуклеотидные последовательности с желаемым процентом идентичности последовательностей. Для получения выравниваний с пробелами в целях сравнения используется Gapped BLAST, как описано в Altschul et al. (1997), Nucl. Acid Res. 25: 3389-3402. В случае применения программ BLAST и Gapped BLAST используются по умолчанию параметры соответствующих программ (NBLAST и XBLAST). Смотри http://www.ncbi.nih.gov. Количественная оценка идентичности также может быть рассчитана с использованием способов и алгоритмов Crickmore et al., как описано выше в разделе, посвященном предпосылке.
Гомология аминокислот будет самой высокой в критических районах токсина, которые отвечают за биологическую активность или вовлечены в определение трехмерной конфигурации, которая, в конечном счете, ответственна за биологическую активность. В этом отношении приемлемы и могут быть вероятными некоторые аминокислотные замены, если эти замены находятся в районах, которые не критичны для активности, или являются консервативными аминокислотными заменами, которые не влияют на трехмерную конфигурацию молекулы. Например, аминокислоты могут быть распределены по следующим классам: неполярные, незаряженные полярные, основные или кислые. Консервативные замены, посредством которых аминокислота одного класса заменяется другой аминокислотой того же типа, входит в рамки заявленного изобретения, поскольку замена существенно не изменяет биологическую активность соединения. В таблице 1 представлен список примеров аминокислот, относящихся к каждому классу.
В некоторых случаях также могут быть сделаны неконсервативные замены. Критическим фактором является то, что данные замены не должны значительно снижать способность растений экспрессировать представленные последовательности ДНК или биологическую активность токсина.
В используемом здесь смысле упоминание "изолированных" полинуклеотидов и/или "очищенных" токсинов относится к данным молекулам в том случае, когда они не ассоциированы с другими молекулами, с которыми они могут быть обнаружены в природе: это может относиться к их использованию в растениях. Таким образом, упоминание "изолированный" и/или "очищенный" означает участие человека, как здесь описано.
Хотя заявленное изобретение предоставляет конкретные варианты синтетических генов, также могут быть использованы другие гены, которые функционально эквивалентны генам, приведенным здесь в качестве примеров, для того, чтобы трансформировать хозяев, предпочтительно растительных хозяев. Дополнительное руководство для получения синтетических генов можно найти, например, в патенте США № 5380831.
Рекомбинантные хозяева. Гены заявленного изобретения, кодирующие токсины, могут быть введены в широкое множество хозяев - микроорганизмов или растений. В предпочтительных вариантах экспрессия гена-токсина прямо или косвенно приводит в результате к внутриклеточной продукции и сохранению пестицида. Когда трансгенные/рекомбинантные/трансформированные клетки хозяина будут проглатываться вредителем, вредители будут поглощать токсин. Данный способ является предпочтительным способом, при котором вызывается контакт вредителя с токсином. Результатом является контроль (уничтожение или ослабление) вредителей.
В некоторых вариантах заявленного изобретения трансформированные хозяева-микроорганизмы могут быть использованы на предварительных этапах получения предшественников, например, таких, которые в конечном итоге будут использованы для трансформации, в предпочтительных вариантах, растительных клеток и растений так, чтобы они экспрессировали токсины, кодируемые генами заявленного изобретения. Трансформированные и использованные таким образом микроорганизмы входят в рамки заявленного изобретения. Рекомбинантными микроорганизмами, например, могут быть B.t., E.coli или Pseudomonas.
Получение различных рекомбинантных организмов может быть осуществлено специалистами в данной области с применением стандартных технологий. Материалы, необходимые для таких трансформаций, сообщаются здесь или иным способом легко доступны специалистам. Имеется широкое множество способов введения гена B.t., кодирующего токсин, в хозяина-мишень в условиях, которые позволяют стабильно охранять и экспрессировать ген. Данные способы хорошо известны специалистам в данной области, и описаны, например, в патенте США №5135867, который включен здесь в виде ссылки.
Гены заявленного изобретения, кодирующие токсины, могут быть введены посредством приемлемого вектора в широкое множество хозяев - микроорганизмов и растений. Существует много культур, представляющих интерес, таких как зерновые, пшеница, рис, хлопок, соя и подсолнечник. Некоторые гены заявленного изобретения, в частности, хорошо подходят для обеспечения стабильного сохранения и экспрессии в трансформированном растении гена, экспрессирующего полипептидный пестицид, и, желательно, обеспечивают улучшенную защиту пестицида от деградации и инактивации под действием окружающей среды. Экспрессия гена-токсина в результате прямо или косвенно приводит к внутриклеточной продукции и сохранению пестицидных белков. Таким образом, вредитель-мишень может контактировать с пестицидными белками при заглатывании растительной ткани, содержащей пестицидные белки, которые токсичны для вредителя. Результатом является контроль вредителя. В альтернативном случае приемлемые хозяева-микроорганизмы, например Pseudomonas fluorescens, могут применяться в местах обитания вредителей, где некоторые из них могут размножаться и проглатываться вредителями-мишенями. Микроорганизмы, в которые помещен ген токсина, могут быть обработаны в условиях, которые продлевают активность токсина и стабилизируют клетку. Затем обработанная клетка, которая сохраняет токсическую активность, может применяться в среде вредителя-мишени.
Поскольку ген токсина B.t. вводят посредством подходящего вектора в хозяина-микроорганизм и указанный хозяин применяется в окружающей среде в живом состоянии, должны использоваться определенные хозяева-микроорганизмы. Выбираются хозяева-микроорганизмы, о которых известно, что они занимают "фитосферу" (филлоплан, филлосфера, ризосфера и/или ризоплан) одного или большего количества интересующих культур. Данные микроорганизмы выбираются так, чтобы они были способны конкурировать в конкретной окружающей среде (сельскохозяйственная культура и среды обитания в других насекомых) с микроорганизмами дикого типа, обеспечивая стабильное сохранение и экспрессию гена, экспрессирующего полипептидный пестицид, и, желательно, обеспечивая улучшенную защиту пестицида от деградации и инактивации под действием окружающей среды.
Известно большое количество микроорганизмов для заселения филлоплана (поверхности листьев растений) и/или ризосферы (почвы, окружающей корни растений) широкого многообразия важных культур. Эти микроорганизмы включают бактерии, водоросли и грибы. Особый интерес представляют такие микроорганизмы, как бактерии, например, родов Pseudomonas, Erwinia, Serratia, Klebsiella, Xanthomonas, Streptomyces, Rhizobium, Rhodopseudomonas, Methylophilius, Agrobacterium, Acetobacter, Lactobacillus, Arthrobacter, Azotobacter, Leuconostoc и Alcaligenes; грибы, особенно дрожжи, например, родов Saccharomyces, Cryptococcus, Kluyveromyces, Sporobolomyces, Rhodotorula и Aureobasidium. Особый интерес представляют такие виды бактерий фитосферы, как Pseudomonas syringae, Pseudomonas fluorescens, Serratia marcescens, Acetobacter xylinum, Agrobacterium tumefaciens, Rhodopseudomonas spheroides, Xanthomonas campestris, Rhizobium melioti, Alcaligenes entrophus и Azotobacter vinlandii; и такие виды дрожжей фитосферы, как Rhodotorula rubra, R. glutinis, R. marina, R. aurantiaca, Cryptococcus albidus, C. diffluens, C. laurentii, Saccharomyces rosei, S. pretoriensis, S. cerevisiae, Sporobolomyces roseus, S. odorus, Kluyveromyces veronae и Aureobasidium pollulans. Особый интерес представляют пигментированные микроорганизмы.
Имеется широкое множество способов введения гена B.t., кодирующего токсин, в хозяина-мишень в условиях, которые позволяют стабильно сохранять и экспрессировать ген. Данные способы хорошо известны специалистам в данной области и описаны, например, в патенте США № 5135867, который включен здесь в виде ссылки.
Обработка клеток. Как отмечалось выше, B.t. или рекомбинантные клетки, экспрессирующие токсин B.t., могут быть обработаны, для того чтобы продлить активность токсина и стабилизировать клетку. Образуемая пестицидная микрокапсула содержит токсин B.t. внутри клеточной структуры, которая была стабилизирована, и будет защищать токсин тогда, когда микрокапсула будет применяться в среде вредителя-мишени. Подходящие клетки-хозяева могут включать либо прокариот, либо эукариот, которые обычно ограничены такими клетками, которые не производят веществ, токсичных для высших организмов, таких как млекопитающие. Однако организмы, которые производят вещества, токсичные для высших организмов, могут использоваться в тех случаях, когда токсичные вещества нестабильны или используемый уровень довольно низкий, так что исключает любую возможность проявления токсичности для хозяев-млекопитающих. В качестве хозяев особый интерес будут представлять прокариоты и низшие эукариоты, такие как грибы.
Как правило, при обработке клетка будет интактной и, в основном, в пролиферативной форме, а не в виде споры, хотя в некоторых случаях могут использоваться споры.
Обработка клеток микроорганизмов, например микроорганизмов, содержащих ген токсина B.t., может осуществляться химическими или физическими способами или при комбинации химических и/или физических способов при условии, что технология не оказывает вредного воздействия на свойства токсина, не уменьшает способность клеток защищать токсин. Примерами химических реагентов являются галогенирующие средства, в частности галогены с номерами атомов 17-80. Более конкретно, можно использовать иод при умеренных условиях и в течение времени, достаточного для достижения желаемого результата. Другие подходящие технологии включают обработку альдегидами, такими как глутаральдегид; антиинфекционными средствами, такими как хлорид зефирана и хлорид цетилпиридиния; спиртами, такими как изопропил и этанол; различными гистологическими фиксажами, такими как иодсодержащий раствор Люголя, фиксатор Боуина, различные кислоты и фиксатор Хелли (смотри: Humason, Gretchen L., Animal Tissue Techniques, W.H.Freeman and Company, 1967); или комбинацией физических (нагревание) и химических средств, которые сохраняют и продлевают активность токсина, продуцируемого в клетке в том случае, когда клетка вводится в среду хозяина. Примерами физических способов являются коротковолновое излучение, такое как гамма-излучение и Х-излучение, замораживание, УФ-излучение, лиофилизация и им подобные. Способы обработки клеток микроорганизмов заявлены в патентах США № 4695455 и 4695462, которые включены здесь в виде ссылки.
Как правило, клетки будут обладать повышенной стабильностью структуры, которая будет повышать устойчивость в условиях окружающей среды. В тех случаях, когда пестицид представляет собой проформу, способ обработки клеток следует выбирать так, чтобы не ингибировать процессинг проформы в зрелую форму пестицида патогенным вредителем-мишенью. Например, формальдегид будет перекрестно сшивать белки и может ингибировать процессинг проформы полипептидного пестицида. Способ обработки должен сохранять, по меньшей мере, значительную часть биодоступности и биоактивности токсина.
Характеристики, представляющие особый интерес при отборе клетки-хозяина в целях продукции, включают простоту введения гена B.t. в хозяина, пригодность систем экспрессии, эффективность экспрессии, стабильность пестицида в хозяине и наличие дополнительных генетических возможностей. Характеристики, представляющие интерес для применения в виде пестицидных микрокапсул, включают особенности защиты пестицида, такие как толщина клеточных стенок, пигментация и внутриклеточная упаковка или образование внутриклеточных телец; выживаемость в водной среде; отсутствие токсичности для млекопитающих; привлечение вредителей для проглатывания; простота уничтожения и фиксации без повреждения токсина и им подобные. Другие соображения включают простоту образования композиции и обработку, экономические характеристики, стабильность при хранении и им подобные.
Выращивание клеток. Клетка-хозяин, содержащая ген инсектицида B.t., может быть выращена в любой обычной питательной среде, предпочтительно там, где конструкция ДНК обеспечивает избирательное преимущество, предоставляя такую селективную среду, чтобы почти все или все клетки сохраняли ген B.t. Затем эти клетки могут быть собраны в соответствии с традиционными способами. Альтернативно клетки могут быть обработаны перед сбором.
Клетки B.t. изобретения можно культивировать, используя стандартные для данной области среды и технологии ферментации. После завершения цикла ферментации бактерии можно собрать предпочтительно отделением спор и кристаллов B.t. от бульона ферментации хорошо известными в данной области способами. Извлеченные споры и кристаллы B.t. могут быть приготовлены в виде смачивающегося порошка, жидкого концентрата, гранул или других композиций при добавлении поверхностно-активных веществ, диспергирующих средств, инертных носителей и других компонентов, чтобы обеспечить обращение и применение для конкретных вредителей-мишеней. Все данные композиции и процедуры применения хорошо известны в данной области.
Композиции. Приготовленные гранулы-ловушки, содержащие аттрактант и споры и кристаллы изолятов B.t., или рекомбинантные микроорганизмы, содержащие гены, которые можно получить из заявленных здесь изолятов B.t., можно вносить в почву. Приготовленный продукт также можно применять в виде покрытия семян или обработки корней, или общей обработки растения на более поздних стадиях цикла культуры. Обработки растений и почвы клетками B.t. могут осуществляться смачивающимися порошками, гранулами или распыляемыми порошками при смешивании с различными инертными материалами, такими как неорганические минеральные вещества (филлосиликаты, карбонаты, сульфаты, фосфаты и им подобные) или ботаническими материалами (порошкообразными початками кукурузы, рисовой шелухой, скорлупой грецкого ореха и им подобными). Композиции могут включать разбрызгивающие-приклеивающие адъюванты, стабилизирующие средства, другие пестицидные добавки или поверхностно-активные вещества. Жидкие композиции могут иметь водную или неводную основу и применяться в виде пен, гелей, суспензий, эмульгированных концентратов или им подобных. Ингредиенты могут включать реологические агенты, поверхностно-активные вещества, эмульгаторы, диспергирующие средства или полимеры.
Как хорошо может быть понятно специалисту в данной области, концентрация пестицида будет широко варьировать в зависимости от природы конкретной композиции, особенно от того, является ли композиция концентратом или используется непосредственно. Пестицид будет присутствовать в концентрации, по меньшей мере, 1 мас.% и может быть 100 мас.%. Сухие композиции будут иметь примерно 1-95 мас.% пестицида, в то время как жидкие композиции, как правило, будут иметь примерно 1-60 мас.% твердых веществ в жидкой фазе. Композиции, как правило, будут иметь примерно от 102 до 104 клеток/мг. Данные композиции будут вводиться примерно в дозе от 50 мг (жидкости или сухого вещества) до 1 кг или больше на гектар.
Композиции можно вносить в среду обитания вредителей, например почву и на листья, опрыскиванием, опыливанием, орошением или подобными способами.
Мутанты. Мутанты изолятов изобретения могут быть получены с помощью процедур, хорошо известных в данной области. Например, аспорогенный мутант может быть получен посредством мутагенеза изолята при действии этилметансульфоната (EMS). Мутанты могут быть получены с использованием ультрафиолетового излучения и нитрозогуанидина в процедурах, хорошо известных в данной области.
Меньший процент аспорогенных мутантов будет оставаться интактным и не лизировать в течение продленных периодов ферментации; данные штаммы обозначают лизис минус (-). Лизис минус штаммы можно идентифицировать в результате скрининга аспорогенных мутантов во встряхиваемом флаконе со средой и отбора таких мутантов, которые в конце ферментации еще интактны и содержат кристаллы токсина. Лизис минус штаммы пригодны для процесса клеточной обработки, который будет давать защищенный инкапсулированный белок токсина.
Чтобы получить устойчивый к фагам вариант указанного аспорогенного мутанта, аликвоту фагового лизата наносят на питательный агар и дают высохнуть. Затем аликвоту чувствительного к фагу бактериального штамма помещают непосредственно поверх высушенного лизата и дают высохнуть. Чашки инкубируют при 30°С. Чашки инкубируют в течение 2 дней и в это время можно увидеть многочисленные колонии, выросшие на агаре. Некоторые из этих колоний отбирают и субкультивируют на чашках с питательным агаром. Данные вероятные устойчивые культуры тестируют на устойчивость, высевая перекрестной штриховкой с фаговым лизатом. Наносят штрих фагового лизата на чашку и дают высохнуть. Затем предполагаемые устойчивые культуры наносят штрихами поперек фагового штриха. В случае устойчивых бактериальных культур нигде на штрихе поперек фагового штриха не видно лизиса после инкубации в течение ночи при 30°С. Затем устойчивость к фагу повторно подтверждается путем посева устойчивой культуры в виде газона на чашку с питательным агаром. Чувствительный штамм также высевают таким же образом, чтобы он служил в качестве позитивного контроля. После высушивания каплю фагового лизата помещают в центр чашки и дают возможность высохнуть. В случае устойчивых культур не видно лизиса в той области, где был помещен фаговый лизат, после инкубации при 30°С в течение 24 часов.
Все патенты, заявки на патенты, предварительные заявки и публикации, на которые здесь ссылаются или которые здесь цитируют, включены в виде ссылок в полном объеме в пределах, в которых они не противоречат заключительным руководствам данной спецификации.
Нижеследующее представляет собой примеры, которые иллюстрируют практическое применение изобретения. Данные примеры не следует истолковывать как ограничивающие.
Пример 1. Биоанализ Cry1F против личинок совки ипсилон (Agrotis ipsilon; Lepidoptera: Noctuidae).
Биологическую активность Cry1F, направленную против личинок совки ипсилон (BCW), определяли, используя стандартные процедуры биоанализа и лиофилизированные порошкообразные препараты MR872 (клон Pseudomonas fluorescens, экспрессирующий cry1Fa/cry1Ab химерный ген, заявленный в патенте США № 5840554). Биоанализ BCW проводили путем включения тестируемых образцов в искусственный корм и контрольного заражения личинок обработанным и необработанным кормом. Все анализы проводили с искусственным кормом для BCW (BioServ Corporation, Frenchtown, HJ). Тесты с добавками в корм проводили, смешивая образцы с искусственным кормом (сначала охлажденным до 55°С или ниже) в соотношении 6 мл суспензии плюс 54 мл корма. Кратные концентрации для обработки получали серийным разведением. После встряхивания и перемешивания данную смесь вливали в пластиковые кюветы, разделенные на ячейки объемом 3 мл (Nutrend Container Corporation, Jacksonville, FL), при норме 30 мл корма на 24-ячеечную кювету (каждую ячейку заполняли наполовину). Кюветы Nutrend с большими лунками использовали для биоанализов с насекомыми третьей возрастной стадии или старше; их также заполняли примерно наполовину искусственным кормом. Чистая вода, не содержащая тестируемого материала, служила контролем. После, по меньшей мере, 30 минут, чтобы дать возможность корму остыть и загустеть, на кормовую смесь помещали личинок (French Ag Resources, Lamberton, MN), одну личинку в ячейку. Затем ячейки герметично закрывали майларовой пленкой (ClearLam Packaging, IL), используя аппарат для термосклеивания, и делали несколько проколов в каждой ячейке, чтобы обеспечить газообмен. Личинок выдерживали при 25°С в течение 6 дней в комнате для содержания в режиме 14:10 (свет : темнота). Смертность и замедление роста регистрировали примерно через шесть дней. Для биоанализа ЛК50 (LK50) выполняли минимум 3 повтора, каждый с 5-7 дозами и примерно по 24 насекомых/дозу. Результаты показаны в таблице 2 (биоанализ против личинок Agrotis ipsilon первой и третьей возрастной стадий с использованием лиофилизированного порошкообразного препарата Cry1F MR872).
Пример 2. Эффективность Cry1F приманки против личинок совки ипсилон.
Cry1F тестировали по активности против личинок совки ипсилон, используя композицию приманки. Клоны MR872 смешивали с приманкой серийного производства (SoilServ, Salinas, СА) и полученную в результате смесь разносили по поверхности почвы выращиваемых в горшках проростков пшеницы (на стадии роста 1.5 Feekes) при норме 50 фунтов/акр (22,68 кг/4046,9 м2). Тестировали две концентрации Cry1F путем добавления различных количеств токсина к приманке, получая нормы, эквивалентные 6,25 г и 12,5 г Cry1F токсина/акр (4046,9 м2). Затем растения заражали личинками 3ей и 4ойвозрастной стадии при норме примерно 1 насекомое на 5 растений, и повреждение растений измеряли через 24-48 часов. Шкала оценок при оценке повреждений простирается в пределах 0-4, при этом 0 означает, что растения полностью срезаны до почвы, и 4 означает, что видимых повреждений нет. В контроли включали растения без добавления приманки ("без приманки"), с добавлением приманки, но без токсина ("пустая приманка") и растения без приманки или насекомых ("контроль"). Результаты показаны на фигурах 1 и 2.
Пример 3. Эффективность трансгенных по Cry1F растений подсолнечника против Agrotis ipsilon, измеренная по росту и смертности насекомых и повреждению растений.
Обработки растений:
- Контрольные проростки на стадии V2
- B.t. проростки на стадии V2 (доноры с фиксированным набором генов); данные растения экспрессировали ген, по существу показанный в SEQ ID NO: 3.
Насекомые: Agrotis ipsilon. Личинки второй возрастной стадии (третье лабораторное поколение).
Эксперимент: B.t. проростки: пять повторов, пять проростков на повтор, одна личинка на проросток. Контроль: пять повторов, пять проростков на повтор, одна личинка на проросток.
Протокол:
- Каждый повтор состоял из пяти семян, которые высевали в прозрачные прямоугольные кюветы (15 см × 25 см) и накрывали, чтобы было можно удержать личинок насекомых.
- В каждом повторе заражали пятью личинками. Кюветы выдерживали в камере при 25°С и 75% влажности. Оценку делали через 24, 48, 72 и 96 часов после заражения.
- Оценка.
1. Растения делили на категории неповрежденных, поврежденных (покусанный стебель, семядоли или листья) или срезанных ниже семядолей. Также была проведена оценка по баллам для каждого повтора: от 1 (неповрежденные растения или со стеблем с несколькими укусами) до 9 (все растения срезаны и более 80% тканей съедено).
2. Насекомые - подсчитывали мертвые личинки через 96 часов после заражения и выживших личинок взвешивали.
3. Данные анализировали посредством анализа таблицы сопряженности признаков.
Результаты: Имеются рисунки, на которых показан повтор четыре (48 и 96 часов после заражения). Данные представлены в следующих таблицах, которые показывают оценку повторов через 24, 48, 72 и 96 часов после заражения Agrotis ipsilon.
Заключение: B.t. проростки проявляют активность по отношению к Agrotis ipsilon при высоком уровне заражения (1 личинка/проросток). Данная активность достаточна для того, чтобы неблагоприятно влиять на рост совок так, что достигается их контроль.
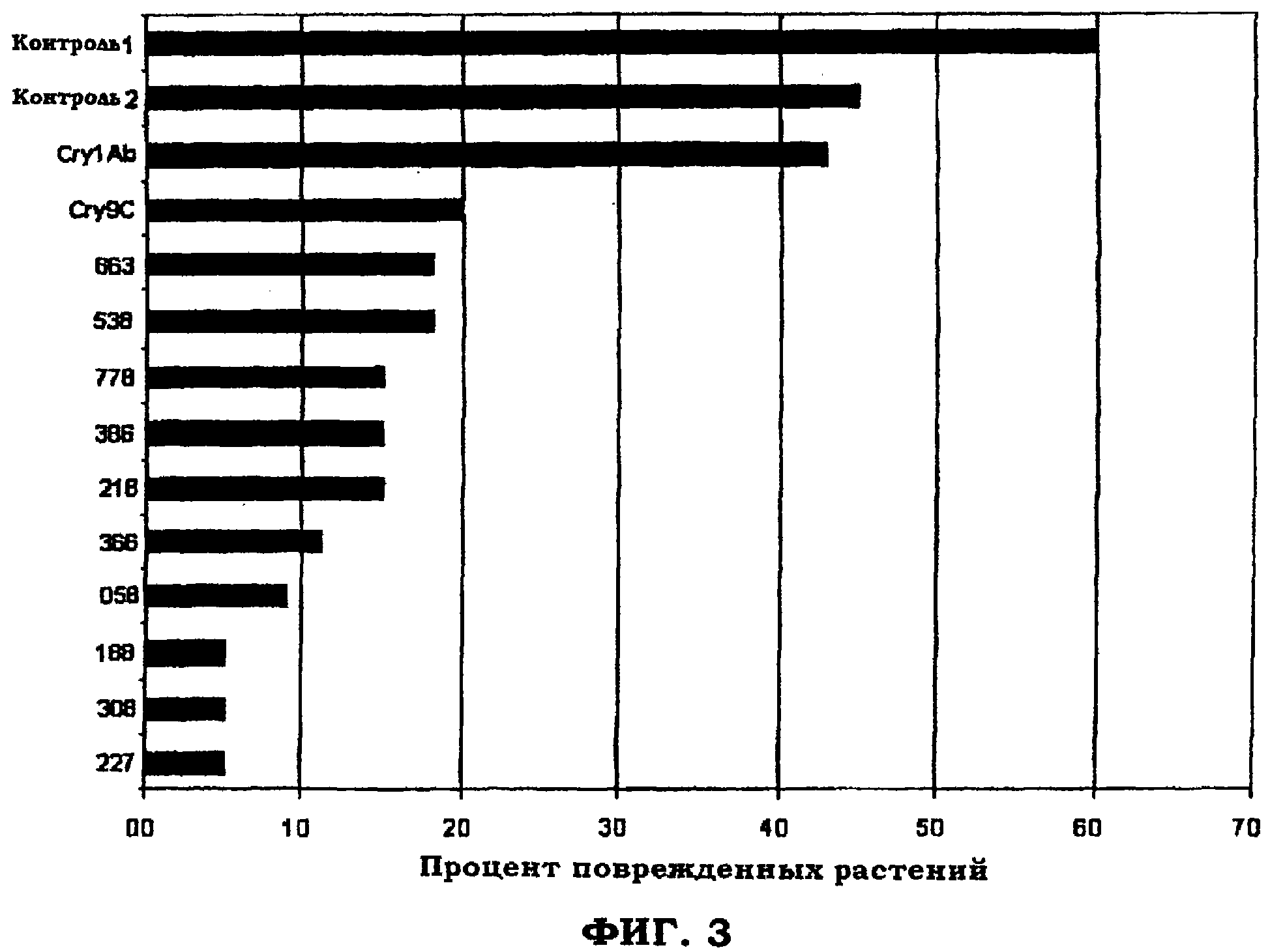
Пример 4. Контроль совок ипсилон (BCW) в случаях новых Cry1F растений кукурузы.
Краткое изложение. Сравнивали способность Cry1F растений кукурузы в десяти случаях (экспрессирующих по существу полинуклеотид SEQ ID NO: 3) контролировать совку ипсилон (Agrotis ipsilon (Hufnagel)) в полевых условиях с двумя не-B.t. гибридами (контроль 1 и контроль 2), гибридом, экспрессирующим Cry1Ab, и гибридом, экспрессирующим Cry9С белок, в целях сравнения. Во всех случаях с Cry1F обеспечивалась хорошая защита против гибели насаждений по сравнению с не-B.t. гибридами.
Материалы и способы. Двадцать пять семян каждого растения-участника испытаний высевали, используя конусообразную сеялку Almaco, в день 1. Высевали четыре повтора - каждый отдельно в случайном порядке. Борозды были парными, между каждой парой борозд была незасеянная дорожка. Каждая делянка была огорожена барьером из гальванизированной стали, который составляет 8 дюймов (20,32 см) в высоту, 2,5 фута (0,76 м) в ширину и 6 футов (1,83 м) в длину. Барьеры вбивали в почву на глубину примерно 2 дюйма (5,08 см) на 13 и 14 день. Края барьеров обрабатывали вазелином, чтобы помешать убежать личинкам. Внутри барьеров разбрасывали солому пшеницы, чтобы обеспечить укрытие для совок. Вскоре после появления взятые для исследования растения с Cry1F тестировали методом ELISA на полосках и насаждения прореживали до 10 растений, удаляя неэкспрессирующие растения или избыток экспрессирующих растений. На трех делянках конечные насаждения представляли собой 8 или 9 растений. Растения, не имеющие Cry1F, также прореживали до 10 растений. Небольшой пластиковый колышек помещали позади каждого растения так, чтобы было видно, сколько растений пропало, если это произойдет. На 15 день, когда растения достигали поздней VI или ранней V2 зрелости, совок ипсилон в третьей личиночной стадии помещали на каждое растение. Второе заражение 2 BCW четвертой возрастной стадии на растение проводили на 19 день.
Делянки проверяли каждый день с 16 дня по 26 день. Дополнительную оценку проводили на 29 день и окончательную оценку делали на 34 день. Какое-либо растение, получившее повреждение, отмечали небольшим пластиковым колышком перед растением и тип повреждения записывали в таблицу данных. В конце испытания делали окончательный подсчет насаждений.
Результаты. В данном испытании был достигнут средний уровень подавления совки ипсилон, при этом для некоторых не-B.t. испытываемых растений получено сокращение насаждений. По неизвестным причинам в данном испытании у BCW имелась тенденция поражать сердцевину растений вместо их срезания. По этой причине также сравнивали данные по поврежденным растениям.
В таблице 8 показан тип растения и процент сокращения насаждений (% СН) и процент поврежденных растений (% Повр.) для каждого типа растений. Сокращение насаждений означает, что растение погибло. Поврежденные растения имели явные повреждения в результате поедания, но сохраняли свою точку роста.
Данные результаты графически показаны на фигурах 3 и 4.
Хотя не было растений-участников испытания без повреждений, это не удивительно, поскольку гусеница должна съесть растение, чтобы погибнуть. Однако, в заключение, имеются фотографии, которые ясно показывают слабое воздействие на Cry1F растения рядом с полностью срезанными контрольными растениями.
Пример 5. Инсерция генов токсина в растения.
Одним аспектом заявленного изобретения является трансформация растений представленными полинуклеотидными последовательностями, кодирующими инсектицидные токсины. Трансформированные растения устойчивы к нападению вредителей-мишеней. Предпочтительными генами согласно заявленному изобретению являются гены, оптимизированные для применения на растениях. Примерами токсинов и генов, активных против совок, для применения согласно заявленному изобретению являются те, которые показаны в SEQ ID NO: 1-8. Предпочтительны белок и ген SEQ ID NO: 1 и 2.
Очевидно, что необходим промоторный район, способный экспрессировать ген. Таким образом, для экспрессии в растении (in planta) ДНК заявленного изобретения находится под контролем и оперативно сцеплена с подходящим промоторным районом. Технологии получения экспрессии in planta с использованием таких конструкций известны в данной области.
Гены, кодирующие пестицидные токсины, которые здесь заявлены, могут быть встроены в клетки растений с помощью множества технологий, которые хорошо известны в данной области. Например, большое количество клонирующих векторов, содержащих систему репликации в Е.coli и маркер, который дает возможность отбора трансформированных клеток, пригодно для препарата для встраивания чужеродных генов в высшие растения. Векторы включают, например, pBR322, pUC серию, M13mp серию, pACYC184 и т.д. Соответственно, последовательность, кодирующая B.t. токсин, может быть встроена в вектор в подходящий сайт рестрикции. Полученную в результате плазмиду используют для трансформации Е. coli. Клетки Е. coli культивируют в подходящей питательной среде, затем собирают и лизируют. Плазмиду извлекают. Как правило, выполняются анализ последовательностей, рестрикционный анализ, электрофорез и другие биохимические - молекулярно-биологические приемы в качестве способов анализа. После каждой манипуляции используемая молекула ДНК может быть расщеплена и соединена с другой последовательностью ДНК. Каждая плазмидная последовательность может быть клонирована в той же самой или в других плазмидах.
В зависимости от способа инсерции требуемого гена в растение могут быть необходимы другие последовательности ДНК. Если, например, для трансформации растительной клетки используется Ti или Ri плазмида, в таком случае, по меньшей мере, правая граница, а часто правая и левая граница Ti или Ri плазмидной Т-ДНК, должна быть соединена в виде фланкирующего района встраиваемых генов. Применение Т-ДНК для трансформации растительных клеток интенсивно исследовано и достаточно описано в ЕР 120516; Hoekema (1985) В: The Binary Plant Vector System, Offset-durkkerij Kanters B.V., Alblasserdam, Chapter 5; Fraley et al., Crit. Rev. Plant Sci. 4: 1-46 и An et al. (1985) EMBO J. 4: 277-287.
Один раз встроенная ДНК интегрирована в геном, там она относительно стабильна и, как правило, снова не выходит. Обычно ДНК содержит селектируемый маркер, который придает трансформированным растительным клеткам устойчивость к биоциду или антибиотику, такому как, в числе других, канамицин, G418, блеомицин, гигромицин или хлорамфеникол. Индивидуально применяемый маркер должен соответственно давать возможность отбора трансформированных клеток, а не клеток, которые не содержат встроенной ДНК.
Имеется большое количество технологий встраивания ДНК в клетку растения-хозяина. Данные технологии включают трансформацию Т-ДНК с использованием Agrobacterium tumefaciens или Agrobacterium rhizogenes в качестве трансформирующего агента, слияния, инъекции, баллистических средств (бомбардировка микрочастицами) или электропорации, а также других возможных способов. Если для трансформации используются агробактерии, встраиваемая ДНК должна быть клонирована в специальных плазмидах, а именно либо в промежуточный вектор, либо в бинарный вектор. Промежуточные векторы могут быть интегрированы в Ti или Ri плазмиду путем гомологичной рекомбинации благодаря последовательностям, которые гомологичны последовательностям в Т-ДНК. Ti или Ri плазмида также содержит район vir, необходимый для переноса Т-ДНК. Промежуточные векторы не могут сами реплицироваться в агробактерии. Промежуточный вектор можно перенести в Agrobacterium tumefaciens посредством хелперной плазмиды (конъюгации). Бинарные векторы могут реплицироваться сами в Е.coli и в агробактерии. Они содержат селектируемый маркерный ген и линкер, или полилинкер, который находится в рамке из районов правой и левой границы Т-ДНК. Они могут быть непосредственно перенесены в результате трансформации в агробактерии (Holsters et al. [1978] Mol. Gen. Genet. 163: 181-187). Agrobacterium, используемая в качестве клетки-хозяина, должна содержать плазмиду, несущую район vir. Район vir необходим для переноса Т-ДНК в клетку растения. Может содержаться дополнительная Т-ДНК. Трансформированная таким образом бактерия используется для трансформации растительных клеток. Растительный эксплантат может предпочтительно культивироваться с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в клетку растения. Затем могут быть регенерированы целые растения из инфицированного растительного материала (например, кусочков листа, кусочков стебля, корней, а также протопластов или клеток суспензионной культуры) в подходящей среде, которая может содержать антибиотики или биоциды для селекции. Затем полученные таким образом растения могут быть тестированы на наличие встроенной ДНК. Особые требования не предъявляются к плазмидам в случае инъекции или электропорации. Возможно применение обычных плазмид, таких, например, как производные pUC.
Трансформированные клетки растут внутри растения обычным образом. Они могут формировать зародышевые клетки и передавать по наследству трансформированное свойство(ва) потомкам растения. Такие растения можно выращивать обычным образом и скрещивать с растениями, которые имеют такие же трансформированные наследственные факторы или другие наследственные факторы. Полученные в результате гибридные индивидуумы обладают соответствующими фенотипическими признаками.
Следует понимать, что описанные здесь примеры и варианты служат только для иллюстративных целей и что в свете этого различные модификации или изменения будут предложены специалистам в данной области, подпадают под объем данной заявки и прилагаемой формулы изобретения.
Реферат
Изобретение относится к сельскому хозяйству. Способ заключается в пестицидном действии токсина Cry1F при потреблении вредителем указанного токсина. Способ позволяет повысить эффективность борьбы с вредителем. 14 з.п.ф-лы, 4 ил., 8 табл.
Комментарии